



Brucella ceti Infection in Striped Dolphins from Italian Seas: Associated Lesions and Epidemiological Data

 , , , ,
, , , ,  , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
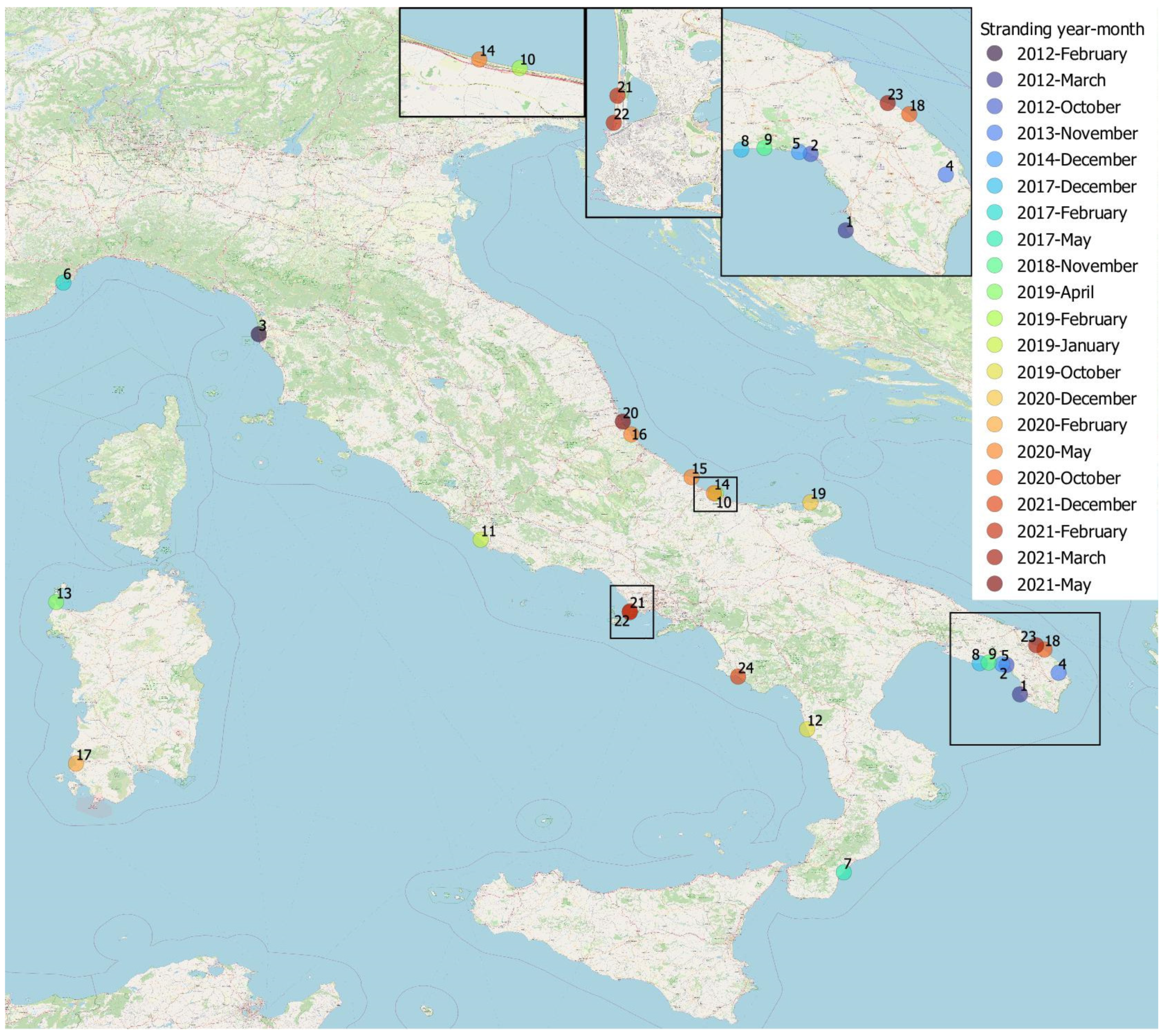
2.1. Dolphins and Samples Included in the Study
2.2. Post-Mortem Examination
2.3. Diagnostic Investigations
2.4. Investigations Focused on Brucella Infection Diagnosis
2.4.1. Brucella Isolation and Identification
2.4.2. Molecular Detection of Brucella spp. from Tissues
2.4.3. Serological Tests for Brucellosis
2.4.4. Whole Genome Sequencing and Bioinformatics
2.5. Cases/Pathological Evaluation
2.6. Statistical Analyses
3. Results
3.1. Postmortem Pathological and Diagnostic Laboratory Investigations
3.1.1. Individual Data and Gastric Contents
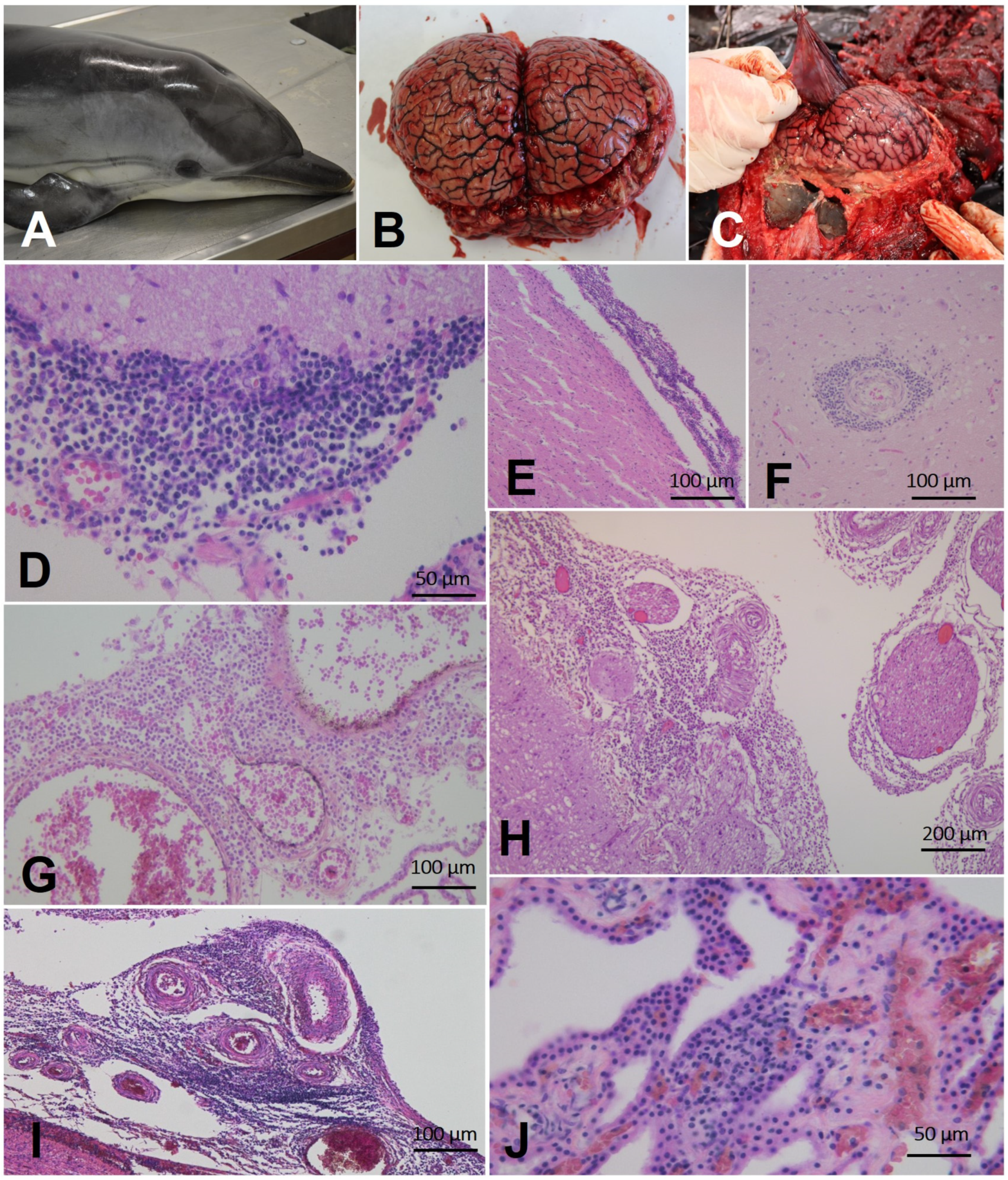
3.1.2. Gross and Microscopic Findings
General Gross Findings
Gross Brucella ceti-Associated Findings
Microscopic Findings Associated with Brucella ceti Infection (CNS)
Microscopic Brucella ceti-Associated Findings (Tissues Other Than CNS)
3.1.3. Coinfections
3.1.4. Cause of Death and Pathological Evaluation
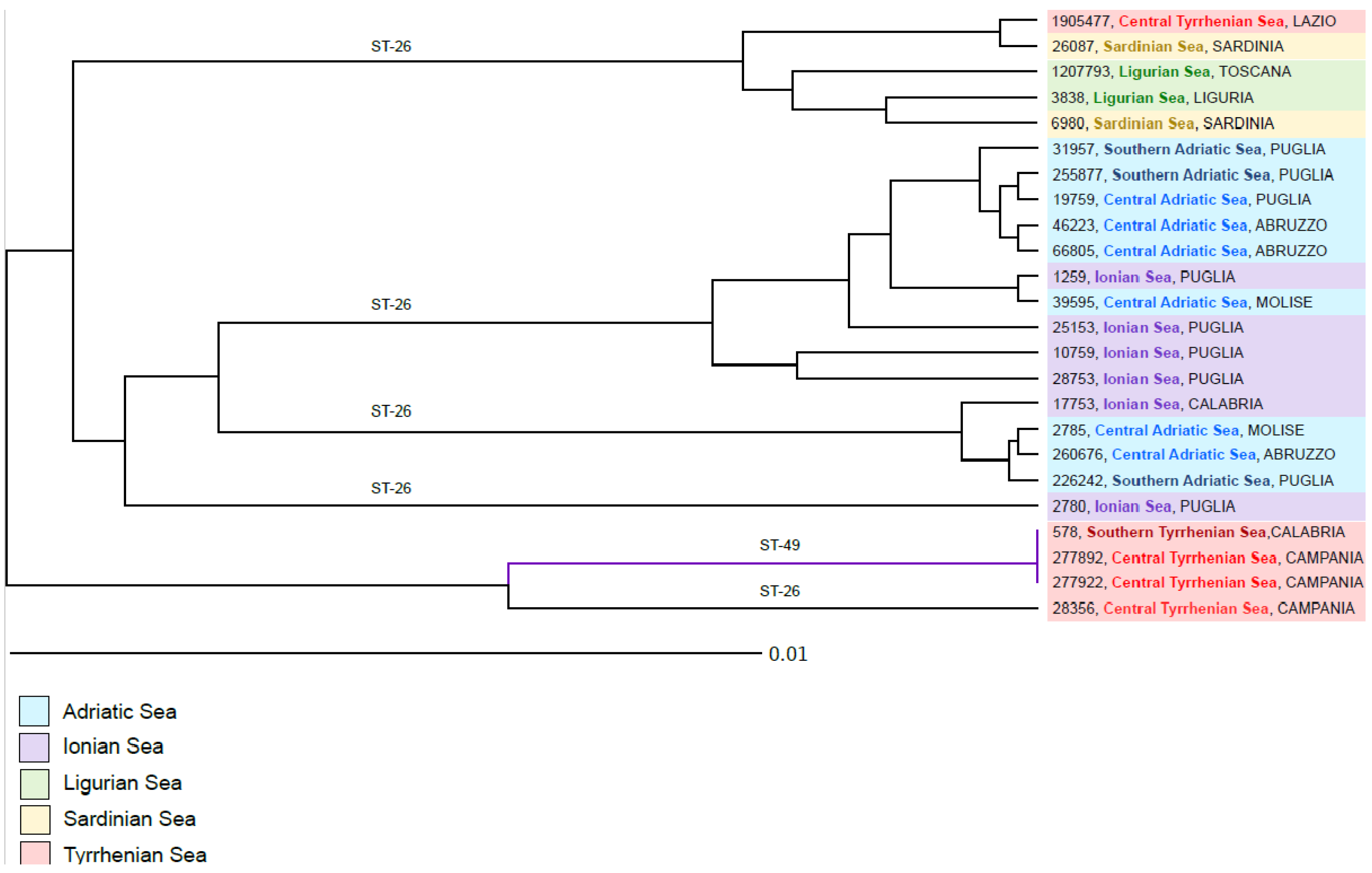
3.2. Brucella Infection Diagnosis
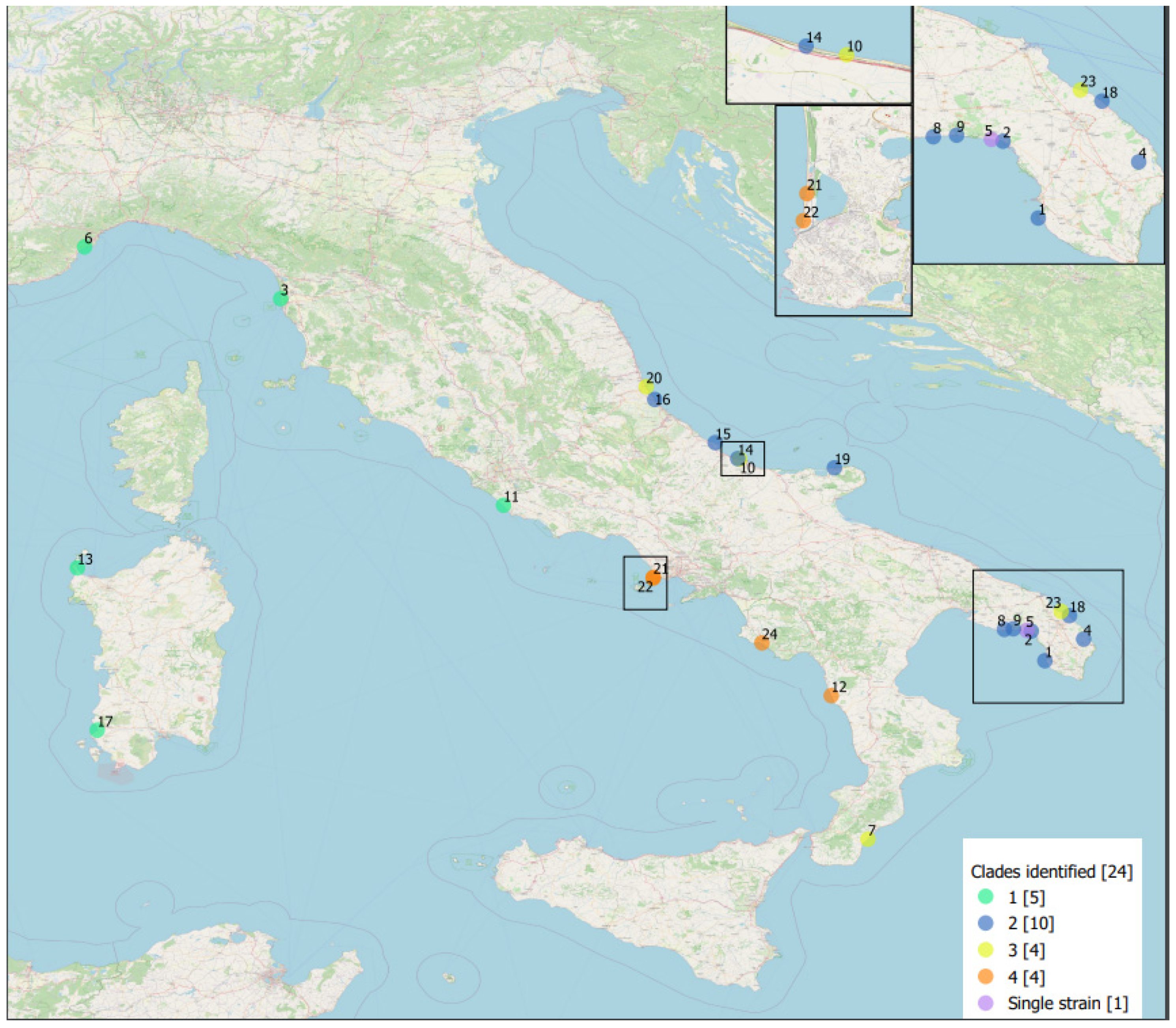
3.3. Brucella ceti Epidemiological Data
3.4. Statistical Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Corbel, M.J.; World Health Organization. Food and Agriculture Organization of the United Nations; International Office of Epizootics. In Brucellosis in Humans and Animals; World Health Organization: Geneva, Switzerland, 2006; ISBN 9241547138. [Google Scholar]
- Whatmore, A.M. Current Understanding of the Genetic Diversity of Brucella, an Expanding Genus of Zoonotic Pathogens. Infect. Genet. Evol. 2009, 9, 1168–1184. [Google Scholar] [CrossRef] [PubMed]
- Godfroid, J.; Scholz, H.C.; Barbier, T.; Nicolas, C.; Wattiau, P.; Fretin, D.; Whatmore, A.M.; Cloeckaert, A.; Blasco, J.M.; Moriyon, I.; et al. Brucellosis at the Animal/Ecosystem/Human Interface at the Beginning of the 21st Century. Prev. Vet. Med. 2011, 102, 118–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewalt, D.R.; Payeur, J.B.; Martin, B.M.; Cummins, D.R.; Miller, W.G. Characteristics of a Brucella Species from a Bottlenose Dolphin (Tursiops Truncatus). J. Vet. Diagn. Investig. 1994, 6, 448–452. [Google Scholar] [CrossRef] [Green Version]
- Ross, H.; Foster, G.; Reid, R.; Jahans, K.; MacMillan, A. Brucella Species Infection in Sea-Mammals. Vet. Rec. 1994, 134, 359. [Google Scholar] [CrossRef]
- Foster, G.; Osterman, B.S.; Godfroid, J.; Jacques, I.; Cloeckaert, A. Brucella ceti sp. Nov. and Brucella pinnipedialis sp. Nov. for Brucella Strains with Cetaceans and Seals as Their Preferred Hosts. Int. J. Syst. Evol. Microbiol. 2007, 57, 2688–2693. [Google Scholar] [CrossRef] [Green Version]
- Whatmore, A.M.; Dawson, C.E.; Groussaud, P.; Koylass, M.S.; King, A.C.; Shankster, S.J.; Sohn, A.H.; Probert, W.S.; McDonald, W.L. Marine Mammal Brucella Genotype Associated with Zoonotic Infection. Emerg. Infect. Dis. 2008, 14, 517–518. [Google Scholar] [CrossRef] [PubMed]
- Nymo, I.H.; Tryland, M.; Godfroid, J. A Review of Brucella Infection in Marine Mammals, with Special Emphasis on Brucella Pinnipedialis in the Hooded Seal (Cystophora cristata). Vet. Res. 2011, 42, 93. [Google Scholar] [CrossRef] [Green Version]
- Guzmán-Verri, C.; González-Barrientos, R.; Hernández-Mora, G.; Morales, J.A.; Baquero-Calvo, E.; Chaves-Olarte, E.; Moreno, E. Brucella Ceti and Brucellosis in Cetaceans. Front. Cell Infect. Microbiol. 2012, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Mora, G.; Palacios-Alfaro, J.D.; González-Barrientos, R. Wild Reservoirs of Brucellosis: Brucella in Aquatic Environments. Rev. Sci. Tech. De. L’oie 2013, 32, 89–103. [Google Scholar] [CrossRef]
- Whatmore, A.M.; Dawson, C.; Muchowski, J.; Perrett, L.L.; Stubberfield, E.; Koylass, M.; Foster, G.; Davison, N.J.; Quance, C.; Sidor, I.F.; et al. Characterisation of North American Brucella Isolates from Marine Mammals. PLoS ONE 2017, 12, e0184758. [Google Scholar] [CrossRef]
- González, L.; Patterson, I.A.; Reid, R.J.; Foster, G.; Barberán, M.; Blasco, J.M.; Kennedy, S.; Howie, F.E.; Godfroid, J.; MacMillan, A.P.; et al. Chronic Meningoencephalitis Associated with Brucella Sp. Infection in Live-Stranded Striped Dolphins (Stenella coeruleoalba). J. Comp. Pathol. 2002, 126, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.G.; Adams, L.G.; Ficht, T.A.; Cheville, N.F.; Payeur, J.P.; Harley, D.R.; House, C.; Ridgway, S.H. Brucella-Induced Abortions, and Infection in Bottlenose Dolphins (Tursiops truncatus). J. Zoo. Wildl. Med. 1999, 30, 100–110. [Google Scholar]
- Foster, G.; MacMillan, A.P.; Godfroid, J.; Howie, F.; Ross, H.M.; Cloeckaert, A.; Reid, R.J.; Brew, S.; Patterson, I.A.P. A Review of Brucella Sp. Infection of Sea Mammals with Particular Emphasis on Isolates from Scotland. Vet. Microbiol. 2002, 90, 563–580. [Google Scholar] [CrossRef]
- Ohishi, K.; Zenitani, R.; Bando, T.; Goto, Y.; Uchida, K.; Maruyama, T.; Yamamoto, S.; Miyazaki, N.; Fujise, Y. Pathological and Serological Evidence of Brucella-Infection in Baleen Whales (Mysticeti) in the Western North Pacific. Comp. Immunol. Microbiol. Infect. Dis. 2003, 26, 125–136. [Google Scholar] [CrossRef]
- Hernández-Mora, G.; González-Barrientos, R.; Morales, J.-A.; Chaves-Olarte, E.; Guzmán-Verri, C.; Baquero-Calvo, E.; De-Miguel, M.-J.; Marín, C.-M.; Blasco, J.-M.; Moreno, E. Neurobrucellosis in Stranded Dolphins, Costa Rica. Emerg. Infect. Dis. 2008, 14, 1430–1433. [Google Scholar] [CrossRef] [PubMed]
- González-Barrientos, R.; Morales, J.-A.; Hernández-Mora, G.; Barquero-Calvo, E.; Guzmán-Verri, C.; Chaves-Olarte, E.; Moreno, E. Pathology of Striped Dolphins (Stenella coeruleoalba) Infected with Brucella Ceti. J. Comp. Pathol. 2010, 142, 347–352. [Google Scholar] [CrossRef]
- Alba, P.; Terracciano, G.; Franco, A.; Lorenzetti, S.; Cocumelli, C.; Fichi, G.; Eleni, C.; Zygmunt, M.S.; Cloeckaert, A.; Battisti, A. The Presence of Brucella Ceti ST26 in a Striped Dolphin (Stenella coeruleoalba) with Meningoencephalitis from the Mediterranean Sea. Vet. Microbiol. 2013, 164, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Sierra, E.; Sánchez, S.; Saliki, J.T.; Blas-Machado, U.; Arbelo, M.; Zucca, D.; Fernández, A. Retrospective Study of Etiologic Agents Associated with Nonsuppurative Meningoencephalitis in Stranded Cetaceans in the Canary Islands. J. Clin. Microbiol. 2014, 52, 2390–2397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, S.C.; Palmer, M.V. Advancement of Knowledge of Brucella Over the Past 50 Years. Vet. Pathol. 2014, 51, 1076–1089. [Google Scholar] [CrossRef]
- Isidoro-Ayza, M.; Ruiz-Villalobos, N.; Pérez, L.; Guzmán-Verri, C.; Muñoz, P.M.; Alegre, F.; Barberán, M.; Chacón-Díaz, C.; Chaves-Olarte, E.; González-Barrientos, R.; et al. Brucella Ceti infection in Dolphins from the Western Mediterranean Sea. BMC Vet. Res. 2014, 10, 206. [Google Scholar] [CrossRef] [Green Version]
- Colegrove, K.; Venn-Watson, S.; Litz, J.; Kinsel, M.; Terio, K.; Fougeres, E.; Ewing, R.; Pabst, D.; McLellan, W.; Raverty, S.; et al. Fetal Distress and in Utero Pneumonia in Perinatal Dolphins during the Northern Gulf of Mexico Unusual Mortality Event. Dis. Aquat. Organ. 2016, 119, 1–16. [Google Scholar] [CrossRef]
- Grattarola, C.; Giorda, F.; Iulini, B.; Pintore, M.D.; Pautasso, A.; Zoppi, S.; Goria, M.; Romano, A.; Peletto, S.; Varello, K.; et al. Meningoencephalitis and Listeria monocytogenes, Toxoplasma gondii and Brucella Spp. Coinfection in a Dolphin in Italy. Dis. Aquat. Organ. 2016, 118, 169–174. [Google Scholar] [CrossRef]
- Davison, N.J.; Perrett, L.L.; Dawson, C.; Dagleish, M.P.; Haskins, G.; Muchowski, J.; Whatmore, A.M. Brucella ceti Infection in a Common Minke Whale (Balaenoptera acutorostrata) with Associated Pathology. J. Wildl. Dis. 2017, 53, 572–576. [Google Scholar] [CrossRef]
- Buckle, K.; Roe, W.D.; Howe, L.; Michael, S.; Duignan, P.J.; Burrows, E.; Ha, H.J.; Humphrey, S.; McDonald, W.L. Brucellosis in Endangered Hector’s Dolphins (Cephalorhynchus hectori). Vet. Pathol. 2017, 54, 838–845. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Sarmiento, A.M.; Carvalho, V.L.; Díaz-Delgado, J.; Ressio, R.A.; Fernandes, N.C.C.A.; Guerra, J.M.; Sacristán, C.; Groch, K.R.; Silvestre-Perez, N.; Ferreira-Machado, E.; et al. Molecular, Serological, Pathological, Immunohistochemical and Microbiological Investigation of Brucella Spp. in Marine Mammals of Brazil Reveals New Cetacean Hosts. Transbound. Emerg. Dis. 2019, 66, 1674–1692. [Google Scholar] [CrossRef] [PubMed]
- Sierra, A.; Paolicelli, R.C.; Kettenmann, H. Cien Años de Microglía: Milestones in a Century of Microglial Research. Trends Neurosci. 2019, 42, 778–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davison, N.; Dagleish, M.; Ten Doeschate, M.; Muchowski, J.; Perrett, L.; Rocchi, M.; Whatmore, A.; Brownlow, A. Meningoencephalitis in a Common Minke Whale Balaenoptera Acutorostrata Associated with Brucella Pinnipedialis and Gamma-Herpesvirus Infection. Dis. Aquat. Organ. 2021, 144, 231–235. [Google Scholar] [CrossRef]
- Curtiss, J.B.; Colegrove, K.M.; Dianis, A.; Kinsel, M.J.; Ahmed, N.; Fauquier, D.; Rowles, T.; Niemeyer, M.; Rotstein, D.S.; Maddox, C.W.; et al. Brucella Ceti Sequence Type 23, 26, and 27 Infections in North American Cetaceans. Dis. Aquat. Organ. 2022, 148, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Granados-Zapata, A.; Robles-Malagamba, M.J.; González-Barrientos, R.; Kot, B.C.-W.; Barquero-Calvo, E.; Cordero-Chavaría, M.; Suárez-Esquivel, M.; Guzmán-Verri, C.; Palacios-Alfaro, J.D.; Tien-Sung, C.; et al. Pathological Studies and Postmortem Computed Tomography of Dolphins with Meningoencephalomyelitis and Osteoarthritis Caused by Brucella ceti. Oceans 2022, 3, 189–203. [Google Scholar] [CrossRef]
- Di Francesco, G.; Petrini, A.; D’angelo, A.R.; Di Renzo, L.; Luciani, M.; Di Febo, T.; Ruggieri, E.; Petrella, A.; Grattarola, C.; Iulini, B.; et al. Immunohistochemical Investigations on Brucella Ceti-Infected, Neurobrucellosis-Affected Striped Dolphins (Stenella coeruleoalba). Vet. Ital. 2019, 55, 363–367. [Google Scholar] [CrossRef]
- Obiako, O.R.; Ogoina, D.; Danbauchi, S.S.; Kwaifa, S.I.; Chom, N.D.; Nwokorie, E. Neurobrucellosis—A Case Report and Review of Literature. Niger. J. Clin. Pract. 2010, 13, 347–350. [Google Scholar]
- Bossart, G.D. Marine Mammals as Sentinel Species for Oceans and Human Health. Vet. Pathol. 2011, 48, 676–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davison, N.J.; Cranwell, M.P.; Perrett, L.L.; Dawson, C.E.; Stubberfield, E.J.; Deaville, R.; Jepson, P.D.; Jarvis, D.S. Meningoencephalitis Associated with Brucella Species in a Live-Stranded Striped Dolphin (Stenella coeruleoalba) in South-West England. Vet. Rec. 2009, 165, 86–89. [Google Scholar] [CrossRef]
- Di Francesco, G.; Di Renzo, L.; Garofolo, G.; Tittarelli, M.; Di Guardo, G. Two Neurotropic Pathogens of Concern for Striped Dolphins. Vet. Rec. 2020, 187, e92. [Google Scholar] [CrossRef]
- Garofolo, G.; Petrella, A.; Lucifora, G.; Di Francesco, G.; Di Guardo, G.; Pautasso, A.; Iulini, B.; Varello, K.; Giorda, F.; Goria, M.; et al. Occurrence of Brucella Ceti in Striped Dolphins from Italian Seas. PLoS ONE 2020, 15, e0240178. [Google Scholar] [CrossRef]
- Di Guardo, G.; Centelleghe, C.; Mazzariol, S. Cetacean Host-Pathogen Interaction(s): Critical Knowledge Gaps. Front. Immunol. 2018, 9, 2815. [Google Scholar] [CrossRef] [PubMed]
- Angelucci, C.B.; Giacominelli-Stuffler, R.; Baffoni, M.; Di Francesco, C.E.; Di Francesco, G.; Di Renzo, L.; Tittarelli, M.; Petrella, A.; Grattarola, C.; Mazzariol, S.; et al. Cellular Prion Protein Expression in the Brain Tissue from Brucella ceti-Infected Striped Dolphins (Stenella coeruleoalba). Animals 2022, 12, 1304. [Google Scholar] [CrossRef]
- Maquart, M.; Le Flèche, P.; Foster, G.; Tryland, M.; Ramisse, F.; Djønne, B.; Al Dahouk, S.; Jacques, I.; Neubauer, H.; Walravens, K.; et al. MLVA-16 Typing of 295 Marine Mammal Brucella Isolates from Different Animal and Geographic Origins Identifies 7 Major Groups within Brucella ceti and Brucella pinnipedialis. BMC Microbiol. 2009, 9, 145. [Google Scholar] [CrossRef]
- Suárez-Esquivel, M.; Baker, K.S.; Ruiz-Villalobos, N.; Hernández-Mora, G.; Barquero-Calvo, E.; González-Barrientos, R.; Castillo-Zeledón, A.; Jiménez-Rojas, C.; Chacón-Díaz, C.; Cloeckaert, A.; et al. Brucella Genetic Variability in Wildlife Marine Mammals Populations Relates to Host Preference and Ocean Distribution. Genome Biol. Evol. 2017, 9, 1901–1912. [Google Scholar] [CrossRef] [Green Version]
- Thompson, L.A.; Goertz, C.E.C.; Quakenbush, L.T.; Burek Huntington, K.; Suydam, R.S.; Stimmelmayr, R.; Romano, T.A. Serological Detection of Marine Origin Brucella Exposure in Two Alaska Beluga Stocks. Animals 2022, 12, 1932. [Google Scholar] [CrossRef] [PubMed]
- Garofolo, G.; Zilli, K.; Troiano, P.; Petrella, A.; Marotta, F.; Di Serafino, G.; Ancora, M.; Di Giannatale, E. Brucella Ceti from Two Striped Dolphins Stranded on the Apulia Coastline, Italy. J. Med. Microbiol. 2014, 63, 325–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cvetnić, Ž.; Duvnjak, S.; Đuras, M.; Gomerčić, T.; Reil, I.; Zdelar-Tuk, M.; Špičić, S. Evidence of Brucella Strain ST27 in Bottlenose Dolphin (Tursiops truncatus) in Europe. Vet. Microbiol. 2016, 196, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Duvnjak, S.; Špičić, S.; Kušar, D.; Papić, B.; Reil, I.; Zdelar-Tuk, M.; Pavlinec, Ž.; Đuras, M.; Gomerčić, T.; Hendriksen, R.S.; et al. Whole-Genome Sequence of the First Sequence Type 27 Brucella Ceti Strain Isolated from European Waters. Genome Announc. 2017, 5, e00988-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geraci, J.R.; Lounsbury, V.J. Marine Mammals Ashore: A Field Guide for Strandings; National Aquarium in Baltimore: Baltimore, Maryland, 2005. [Google Scholar]
- Ijsseldijk, L.L.; Brownlow, A.C.; Mazzariol, S. Best Practice on Cetacean Postmortem Investigation and Tissue Sampling Joint ACCOBAMS and ASCOBANS Document Editors. In Proceedings of the 25th Meeting of the Advisory Committee, Stralsund, Germany, 17–19 September 2019. [Google Scholar]
- Carlini, R. Biometric Measures Indicating Sexual Dimorphism in Stenella Coeruleoalba (Meyen, 1833) (Delphinidae) in the North-Central Tyrrhenian Sea. Aquat Mamm 2014, 40, 59–68. [Google Scholar] [CrossRef]
- Perrin, W.F.; Reilly, S.B. Reproductive Parameters of Dolphins, and Small Whales of the Family Deiphinidae. Rep. Int. Whal. Comm. 1984, 6, 97–133. [Google Scholar]
- Cozzi, B.; Huggenberger, S.; Oelschläger, H. Urinary System, Genital Systems, and Reproduction. In Anatomy of Dolphins; Elsevier: Amsterdam, The Netherlands, 2017; pp. 369–409. [Google Scholar]
- Anderson, R.M. The Regulation of Host Population Growth by Parasitic Species. Parasitology 1978, 76, 119–157. [Google Scholar] [CrossRef] [PubMed]
- Khalil, L.F.; Jones, A.; Bray, R.A. Keys to the Cestode Parasites of Vertebrates. Trans. R. Soc. Trop. Med. Hyg. 1995, 89, 344. [Google Scholar] [CrossRef]
- Pintore, M.D.; Mignone, W.; Di Guardo, G.; Mazzariol, S.; Ballardini, M.; Florio, C.L.; Goria, M.; Romano, A.; Caracappa, S.; Giorda, F.; et al. Neuropathologic Findings in Cetaceans Stranded in Italy (2002–2014). J. Wildl. Dis. 2018, 54, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Di Guardo, G.; Proietto, U.; Di Francesco, C.E.; Marsilio, F.; Zaccaroni, A.; Scaravelli, D.; Mignone, W.; Garibaldi, F.; Kennedy, S.; Forster, F.; et al. Cerebral Toxoplasmosis in Striped Dolphins (Stenella coeruleoalba) Stranded Along the Ligurian Sea Coast of Italy. Vet. Pathol. 2010, 47, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Verna, F.; Giorda, F.; Miceli, I.; Rizzo, G.; Pautasso, A.; Romano, A.; Iulini, B.; Pintore, M.D.; Mignone, W.; Grattarola, C.; et al. Detection of Morbillivirus Infection by RT-PCR RFLP Analysis in Cetaceans and Carnivores. J. Virol. Methods 2017, 247, 22–27, Erratum in J. Virol. Methods 2018, 262, 89. [Google Scholar] [CrossRef]
- Vitale, M. A Highly Sensitive Nested PCR for Toxoplasma Gondii Detection in Animal and Food Samples. J. Microb. Biochem. Technol. 2013, 5, 1000097. [Google Scholar] [CrossRef] [Green Version]
- VanDevanter, D.R.; Warrener, P.; Bennett, L.; Schultz, E.R.; Coulter, S.; Garber, R.L.; Rose, T.M. Detection and Analysis of Diverse Herpesviral Species by Consensus Primer PCR. J. Clin. Microbiol. 1996, 34, 1666–1671. [Google Scholar] [CrossRef] [PubMed]
- OIE. OIE Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, 8th ed.; OIE: Paris, France, 2018. [Google Scholar]
- Alton, G.G.; Jones, L.M.; Pietz, D.E. Laboratory Techniques in Brucellosis; World Health Organization: Geneva, Switzerland, 1988. [Google Scholar]
- Cloeckaert, A.; Verger, J.-M.; Grayon, M.; Grepinet, O. Restriction Site Polymorphism of the Genes Encoding the Major 25 KDa and 36 KDa Outer-Membrane Proteins of Brucella. Microbiology 1995, 141, 2111–2121. [Google Scholar] [CrossRef] [Green Version]
- Baily, G.G.; Krahn, J.B.; Drasar, B.S.; Stoker, N.G. Detection of Brucella Melitensis and Brucella Abortus by DNA Amplification. J. Trop. Med. Hyg. 1992, 95, 271–275. [Google Scholar]
- Bounaadja, L.; Albert, D.; Chénais, B.; Hénault, S.; Zygmunt, M.S.; Poliak, S.; Garin-Bastuji, B. Real-Time PCR for Identification of Brucella Spp.: A Comparative Study of IS711, Bcsp31 and per Target Genes. Vet. Microbiol. 2009, 137, 156–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Goñi, I.; García-Yoldi, D.; Marín, C.M.; de Miguel, M.J.; Muñoz, P.M.; Blasco, J.M.; Jacques, I.; Grayon, M.; Cloeckaert, A.; Ferreira, A.C.; et al. Evaluation of a Multiplex PCR Assay (Bruce-Ladder) for Molecular Typing of All Brucella Species, Including the Vaccine Strains. J. Clin. Microbiol. 2008, 46, 3484–3487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.-I.; Her, M.; Kim, J.W.; Kim, J.-Y.; Ko, K.Y.; Ha, Y.-M.; Jung, S.C. Advanced Multiplex PCR Assay for Differentiation of Brucella Species. Appl. Env. Microbiol. 2011, 77, 6726–6728. [Google Scholar] [CrossRef] [Green Version]
- Andrews, K.R.; Karczmarski, L.; Au, W.W.L.; Rickards, S.H.; Vanderlip, C.A.; Bowen, B.W.; Gordon Grau, E.; Toonen, R.J. Rolling Stones, and Stable Homes: Social Structure, Habitat Diversity and Population Genetics of the Hawaiian Spinner Dolphin (Stenella longirostris). Mol. Ecol. 2010, 19, 732–748. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [Green Version]
- Jünemann, S.; Sedlazeck, F.J.; Prior, K.; Albersmeier, A.; John, U.; Kalinowski, J.; Mellmann, A.; Goesmann, A.; von Haeseler, A.; Stoye, J.; et al. Updating Benchtop Sequencing Performance Comparison. Nat. Biotechnol. 2013, 31, 294–296. [Google Scholar] [CrossRef] [Green Version]
- Arbelo, M.; Espinosa de los Monteros, A.; Herráez, P.; Andrada, M.; Sierra, E.; Rodríguez, F.; Jepson, P.; Fernández, A. Pathology and Causes of Death of Stranded Cetaceans in the Canary Islands (1999–2005). Dis. Aquat. Organ. 2013, 103, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Giorda, F.; Ballardini, M.; Di Guardo, G.; Pintore, M.D.; Grattarola, C.; Iulini, B.; Mignone, W.; Goria, M.; Serracca, L.; Varello, K.; et al. Postmortem Findings in Cetaceans Found Stranded in the Pelagos Sanctuary, Italy, 2007–2014. J Wildl Dis 2017, 53, 795–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sierra, E.; Fernández, A.; Felipe-Jiménez, I.; Zucca, D.; Díaz-Delgado, J.; Puig-Lozano, R.; Câmara, N.; Consoli, F.; Díaz-Santana, P.; Suárez-Santana, C.; et al. Histopathological Differential Diagnosis of Meningoencephalitis in Cetaceans: Morbillivirus, Herpesvirus, Toxoplasma gondii, Brucella sp., and Nasitrema sp. Front. Vet. Sci. 2020, 7, 650. [Google Scholar] [CrossRef]
- Shakir, R.A.; Al-Din, A.S.N.; Araj, G.F.; Lulu, A.R.; Mousa, A.R.; Saadah, M.A. Clinical Categories of Neurobrucellosis. Brain 1987, 110, 213–223. [Google Scholar] [CrossRef]
- Davison, N.J.; Brownlow, A.; Doeschate, M.T.; Dale, E.-J.; Foster, G.; Muchowski, J.; Perrett, L.L.; Rocchi, M.; Whatmore, A.M.; Dagleish, M.P. Neurobrucellosis Due to Brucella Ceti ST26 in Three Sowerby’s Beaked Whales (Mesoplodon bidens). J. Comp. Pathol. 2021, 182, 1–8. [Google Scholar] [CrossRef]
- West, K.L.; Levine, G.; Jacob, J.; Jensen, B.; Sanchez, S.; Colegrove, K.; Rotstein, D. Coinfection and Vertical Transmission of Brucella and morbillivirus in a Neonatal Sperm Whale (Physeter macrocephalus) in Hawaii, USA. J. Wildl. Dis. 2015, 51, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Castro, I.; Melero, M.; Crespo-Picazo, J.L.; de los Á Jiménez, M.; Sierra, E.; Rubio-Guerri, C.; Arbelo, M.; Fernández, A.; García-Párraga, D.; Sánchez-Vizcaíno, J.M. Systematic Determination of Herpesvirus in Free-Ranging Cetaceans Stranded in the Western Mediterranean: Tissue Tropism and Associated Lesions. Viruses 2021, 13, 2180. [Google Scholar] [CrossRef]
- Kennedy, S.; Lindstedt, I.J.; McAliskey, M.M.; McConnell, S.A.; McCullough, S.J. Herpesviral Encephalitis in a Harbor Porpoise (Phocoena phocoena). J. Zoo. Wildl. Med. 1992, 23, 374–379. [Google Scholar]
- Esperón, F.; Fernández, A.; Sánchez-Vizcaíno, J. Herpes Simplex-like Infection in a Bottlenose Dolphin Stranded in the Canary Islands. Dis. Aquat. Organ. 2008, 81, 73–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Elk, C.; van de Bildt, M.; van Run, P.; de Jong, A.; Getu, S.; Verjans, G.; Osterhaus, A.; Kuiken, T. Central Nervous System Disease and Genital Disease in Harbor Porpoises (Phocoena phocoena) Are Associated with Different Herpesviruses. Vet. Res. 2016, 47, 28. [Google Scholar] [CrossRef] [Green Version]
- Blanchard, T.W.; Santiago, N.T.; Lipscomb, T.P.; Garber, R.L.; McFee, W.E.; Knowles, S. Two Novel Alphaherpesviruses Associated with Fatal Disseminated Infections in Atlantic Bottlenose Dolphins. J. Wildl. Dis. 2001, 37, 297–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arbelo, M.; Sierra, E.; Esperón, F.; Watanabe, T.; Bellière, E.; Espinosa de los Monteros, A.; Fernández, A. Herpesvirus Infection with Severe Lymphoid Necrosis Affecting a Beaked Whale Stranded in the Canary Islands. Dis. Aquat. Organ. 2010, 89, 261–264. [Google Scholar] [CrossRef] [Green Version]
- Soto, S.; González, B.; Willoughby, K.; Maley, M.; Olvera, A.; Kennedy, S.; Marco, A.; Domingo, M. Systemic Herpesvirus and Morbillivirus Co-Infection in a Striped Dolphin (Stenella coeruleoalba). J. Comp. Pathol. 2012, 146, 269–273. [Google Scholar] [CrossRef]
- Van Bressem, M.-F.; Duignan, P.; Banyard, A.; Barbieri, M.; Colegrove, K.; De Guise, S.; Di Guardo, G.; Dobson, A.; Domingo, M.; Fauquier, D.; et al. Cetacean Morbillivirus: Current Knowledge and Future Directions. Viruses 2014, 6, 5145–5181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nymo, I.H.; Godfroid, J.; Åsbakk, K.; Larsen, A.K.; das Neves, C.G.; Rødven, R.; Tryland, M. A Protein A/G Indirect Enzyme-Linked Immunosorbent Assay for the Detection of Anti-Brucella Antibodies in Arctic Wildlife. J. Vet. Diagn. Investig. 2013, 25, 369–375. [Google Scholar] [CrossRef] [Green Version]
- Roca-Monge, K.; González-Barrientos, R.; Suárez-Esquivel, M.; Palacios-Alfaro, J.D.; Castro-Ramírez, L.; Jiménez-Soto, M.; Cordero-Chavarría, M.; García-Párraga, D.; Barratclough, A.; Moreno, E.; et al. Age and Sexual Maturity Estimation of Stranded Striped Dolphins, Stenella Coeruleoalba, Infected with Brucella Ceti. Oceans 2022, 3, 494–508. [Google Scholar] [CrossRef]
- Muñoz, P.M.; Mick, V.; Sacchini, L.; Janowicz, A.; de Miguel, M.J.; Cherfa, M.-A.; Nevado, C.R.; Girault, G.; Andrés-Barranco, S.; Jay, M.; et al. Phylogeography and Epidemiology of Brucella Suis Biovar 2 in Wildlife and Domestic Swine. Vet. Microbiol. 2019, 233, 68–77. [Google Scholar] [CrossRef]
- Davison, N.J.; Perrett, L.L.; Law, R.J.; Dawson, C.E.; Stubberfield, E.J.; Monies, R.J.; Deaville, R.; Jepson, P.D. Infection with Brucella ceti and High Levels of Polychlorinated Biphenyls in Bottlenose Dolphins (Tursiops truncatus) Stranded in South-West England. Vet. Rec. 2011, 169, 14. [Google Scholar] [CrossRef] [PubMed]
- Nymo, I.H.; das Neves, C.G.; Tryland, M.; Bårdsen, B.-J.; Santos, R.L.; Turchetti, A.P.; Janczak, A.M.; Djønne, B.; Lie, E.; Berg, V.; et al. Brucella Pinnipedialis Hooded Seal (Cystophora cristata) Strain in the Mouse Model with Concurrent Exposure to PCB 153. Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 195–204. [Google Scholar] [CrossRef]
- Brew, S.D.; Perrett, L.L.; Stack, J.A.; MacMillan, A.P.; Staunton, N.J. Human Exposure to Brucella Recovered from a Sea Mammal. Vet. Rec. 1999, 144, 483. [Google Scholar]
- Sohn, A.H.; Probert, W.S.; Glaser, C.A.; Gupta, N.; Bollen, A.W.; Wong, J.D.; Grace, E.M.; McDonald, W.C. Human Neurobrucellosis with Intracerebral Granuloma Caused by a Marine Mammal Brucella Spp. Emerg. Infect. Dis. 2003, 9, 485–488. [Google Scholar] [CrossRef]
- McDonald, W.L.; Jamaludin, R.; Mackereth, G.; Hansen, M.; Humphrey, S.; Short, P.; Taylor, T.; Swingler, J.; Dawson, C.E.; Whatmore, A.M.; et al. Characterization of a Brucella Sp. Strain as a Marine-Mammal Type despite Isolation from a Patient with Spinal Osteomyelitis in New Zealand. J. Clin. Microbiol. 2006, 44, 4363–4370. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case ID | Histological | Immuno-Histochemical | Biomolecular | Serological | Brucella Infection Diagnosis | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| CeMV ° | T. gondii °° | HV °°° | CeMV * | T. gondii* | Isolation and Identification ** | PCR from Tissues | Serological (RBT) * and *** | |||
| 1 | x | x | x | X | ||||||
| 2 | x | x | x | X | ||||||
| 3 | x | x | x | x | x | X | X ^ | |||
| 4 | x | x | X | |||||||
| 5 | x | x | x | X | ||||||
| 6 | x | X (Morbillivirus; T. gondii) * | x | x | x | x | X | X ^ | x | |
| 7 | x | x | x | x | x | X | X ^ | x | ||
| 8 | x | x | x | X | X ^^^ | |||||
| 9 | x | x | x | X | X ^^^ | x | ||||
| 10 | x | x | x | X | X ^^ | |||||
| 11 | x | x | x | x | X | X ^ | ||||
| 12 | x | X (Morbillivirus; T. gondii) * | x | x | x | X | X ^ | x | ||
| 13 | x | x | x | X | X ^ | |||||
| 14 | x | x | x | x | X | X ^^ | x | |||
| 15 | x | x | x | x | x | X | X ^^ | x | ||
| 16 | x | x | x | X | X ^^ | |||||
| 17 | x | x | x | x | X | X ^ | ||||
| 18 | x | x | x | X | x | |||||
| 19 | x | x | x | X | x | |||||
| 20 | x | x | x | X | X ^^ | x | ||||
| 21 | x | X (Morbillivirus; T. gondii) * | x | x | x | X | X ^ | |||
| 22 | x | X (Morbillivirus) * | x | x | x | X | X ^ | |||
| 23 | x | x | x | X | x | |||||
| 24 | x | x | x | x | x | X ^^ | ||||
| Gross Lesions | Striped Dolphins Examined |
|---|---|
| Hyperemic meninges and/or brain | 17 of 23 (73.9%) |
| Reproductive tract inflammation | 4 of 23 (17.3%) |
| Hemorrhagic cerebrospinal fluid/increased volume | 3 of 8 (37.5%) |
| Microscopic lesions | |
| Non-suppurative encephalitis (±meningitis/myelitis) | 20 of 23 (86.9%) |
| Splenitis (±necrosis) | 4 of 14 (20%) |
| Hepatitis | 2 of 15 (13.3%) |
| Myocarditis (±necrosis) | 2 of 11 (18.1%) |
| Reproductive tract inflammation | 4 of 12 (33.3%) |
| Lymphadenitis (±necrosis) | 4 of 14 (33.3%) |
| Pneumonia | 3 of 19 (15.7%) |
| Mastitis | 1 of 3 (33.3%) |
| Case ID | Cerebrum | Other Nervous System Regions Different from the Cerebrum | Associated Lesions Related to Other Pathogens | Co-Infections (CNS) | ||
|---|---|---|---|---|---|---|
| M | E | Pl-C | ||||
| 1 | + | - | - | - | - | - |
| 2 | + | - | - | NS cerebellitis | - | |
| 3 | + * | + | - | - | scattered protozoan cysts with a few granulomatous foci | T. gondii |
| 5 | + * | + * | + | NS inflammation in medulla oblongata * | ||
| 6 | + | + | + | granulomatous cerebellitis, with malacia and protozoan cysts | protozoan cysts | CeMV— T. gondii |
| 7 | + | + | - | - | - | CeMV |
| 8 | + * | + * | + | NS myelitis | - | - |
| 9 | + | + | - | NS cerebellitis and myelitis | - | CeMV |
| 10 | + | - | - | NS cerebellitis, NS perineuritis and periganglioneuritis at spinal cord cervical level | CeMV | |
| 14 | + | + * | + | NS cerebellitis, myelitis. | - | |
| 15 | + * | - | - | NS inflammation in medulla oblongata and perineuritis | - | |
| 16 | + | - | - | NS inflammation in medulla oblongata | ||
| 17 | + * | - | - | - | - | - |
| 18 | + | + | + | NS cerebellitis, inflammation in medulla oblongata | - | |
| 19 | + | + | + | NS inflammation in medulla oblongata and myelitis | - | - |
| 20 | + | + | - | NS inflammation in medulla oblongata and myelitis | - | - |
| 21 | + | + | + | T. gondii-HV | ||
| 22 | + | + * | + | - | ||
| 23 | + | + | + | NS cerebellitis | - | - |
| 24 | + | + | + | - | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grattarola, C.; Petrella, A.; Lucifora, G.; Di Francesco, G.; Di Nocera, F.; Pintore, A.; Cocumelli, C.; Terracciano, G.; Battisti, A.; Di Renzo, L.; et al. Brucella ceti Infection in Striped Dolphins from Italian Seas: Associated Lesions and Epidemiological Data. Pathogens 2023, 12, 1034. https://doi.org/10.3390/pathogens12081034
Grattarola C, Petrella A, Lucifora G, Di Francesco G, Di Nocera F, Pintore A, Cocumelli C, Terracciano G, Battisti A, Di Renzo L, et al. Brucella ceti Infection in Striped Dolphins from Italian Seas: Associated Lesions and Epidemiological Data. Pathogens. 2023; 12(8):1034. https://doi.org/10.3390/pathogens12081034
Chicago/Turabian StyleGrattarola, Carla, Antonio Petrella, Giuseppe Lucifora, Gabriella Di Francesco, Fabio Di Nocera, Antonio Pintore, Cristiano Cocumelli, Giuliana Terracciano, Antonio Battisti, Ludovica Di Renzo, and et al. 2023. "Brucella ceti Infection in Striped Dolphins from Italian Seas: Associated Lesions and Epidemiological Data" Pathogens 12, no. 8: 1034. https://doi.org/10.3390/pathogens12081034