



Comparative Analysis of Campylobacter jejuni and C. coli Isolated from Livestock Animals to C. jejuni and C. coli Isolated from Surface Water Using DNA Sequencing and MALDI-TOF

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collections of Strains
2.2. MLST and cgMLST
2.3. Source Attribution Using STRUCTURE
2.4. MALDI-Typing
3. Results
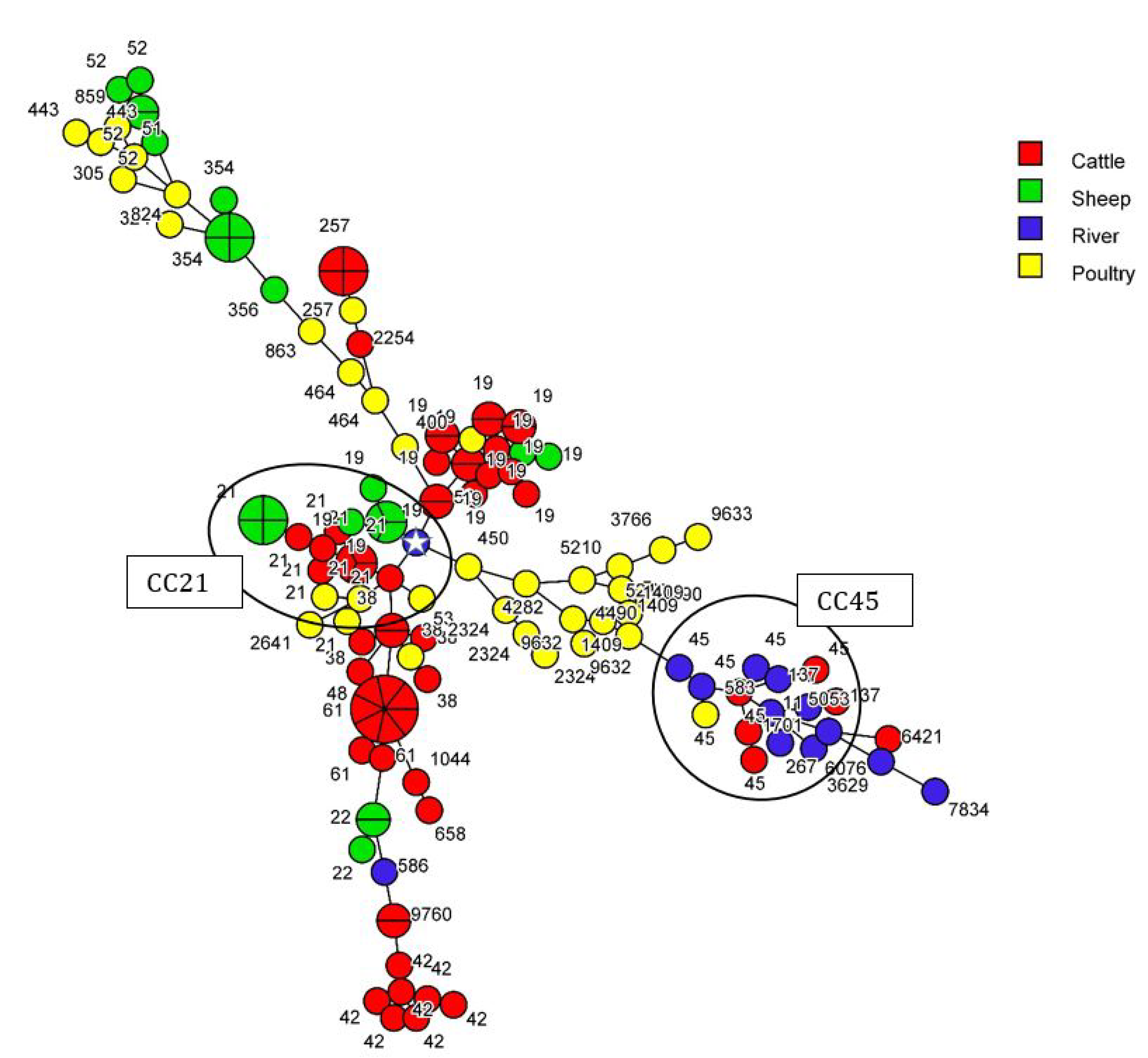
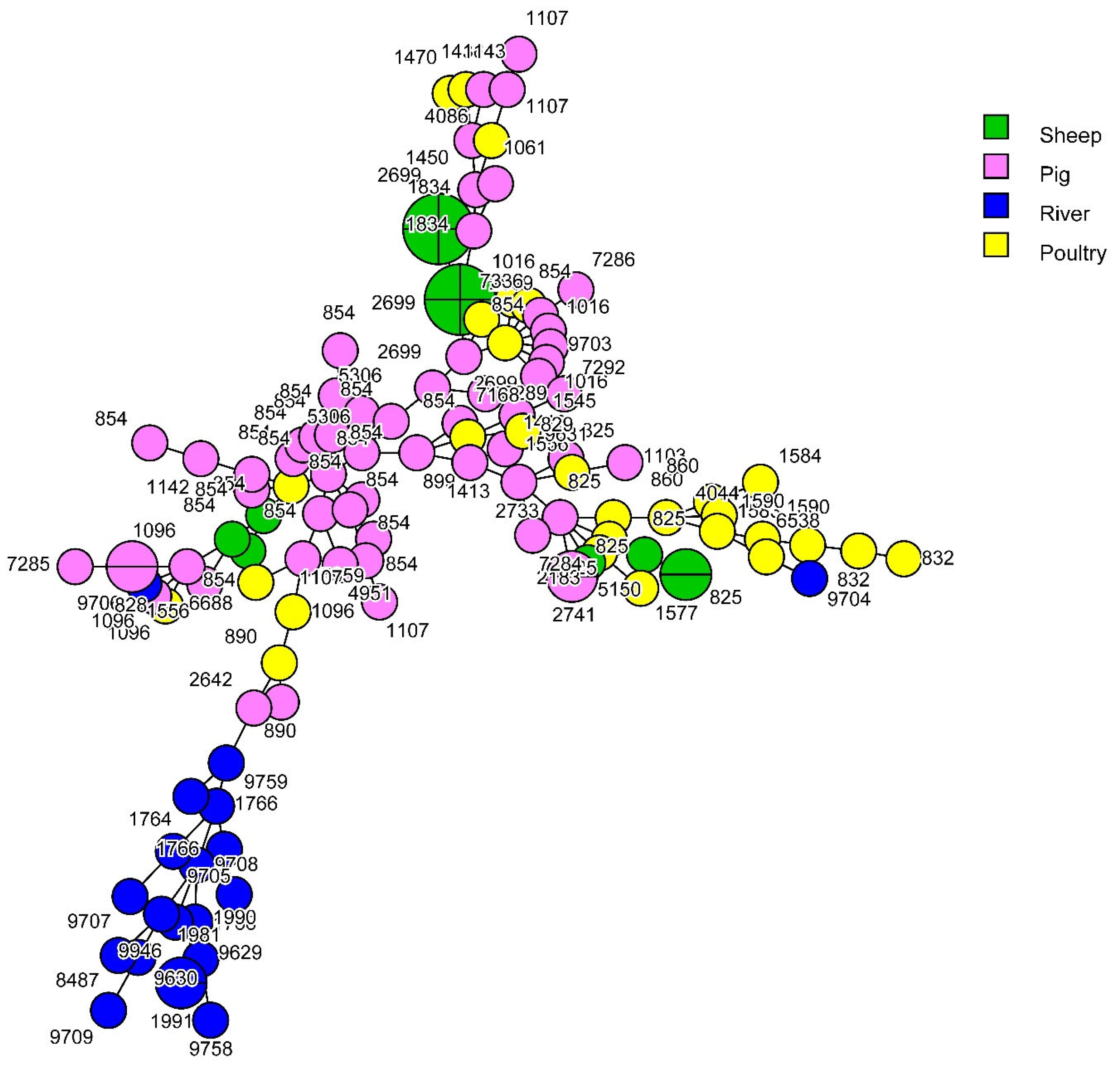
3.1. Contribution of Livestock Type to Surface Water Contamination with Genome Comparison
3.2. Source Attribution of C. jejuni and C. coli Strains Retrieved from Surface Water
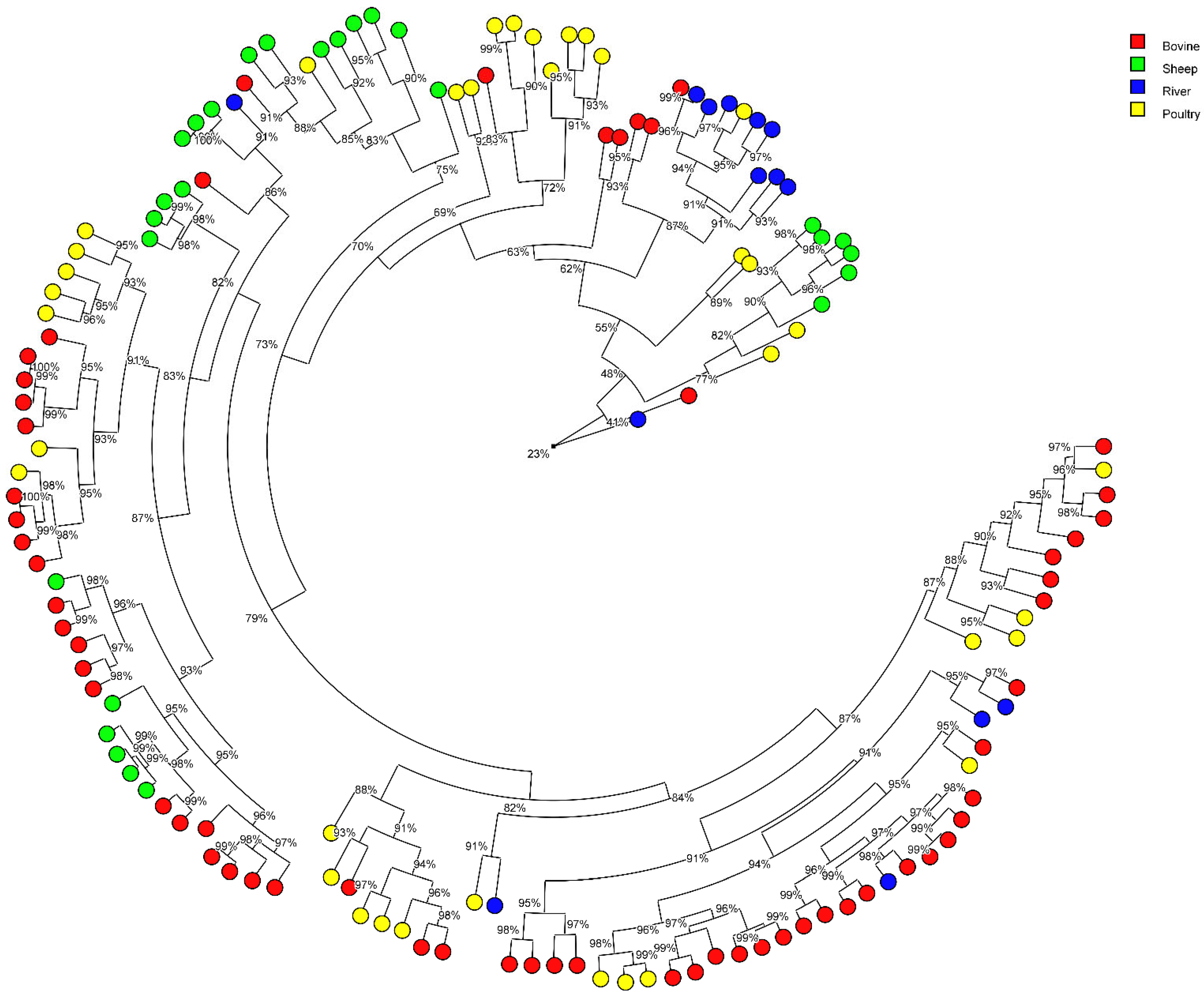
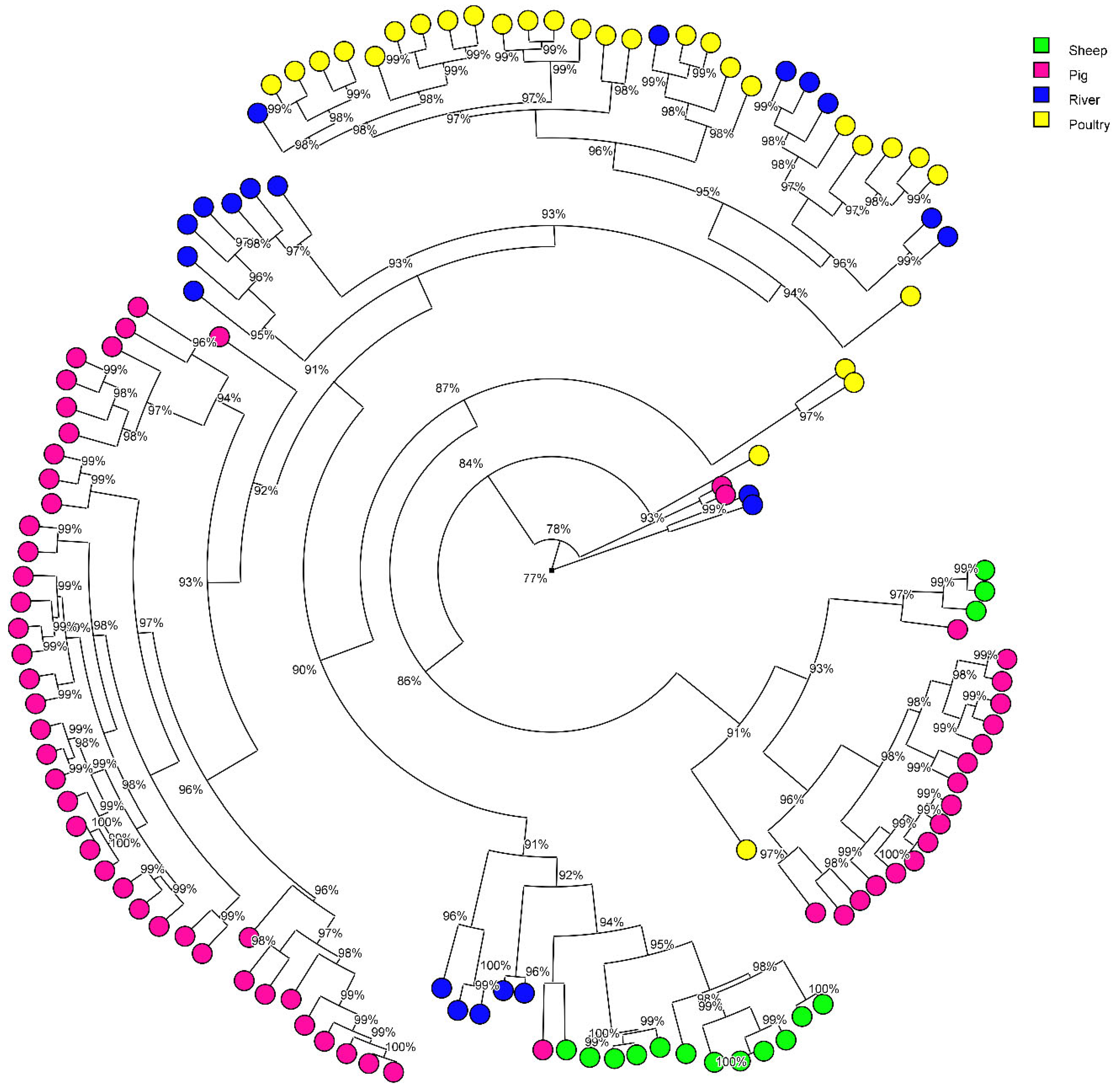
3.3. Contribution of Animal Production in the Surface Water Contamination with MALDI Typing Comparison
4. Discussion
4.1. Diversity of Sequence Types in Cattle, Sheep, Poultry and Pigs
4.2. Sources of Campylobacter Contamination of Rivers
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Cauteren, D.; De Valk, H.; Sommen, C.; King, L.A.; Jourdan-Da Silva, N.; Weill, F.X.; Le Hello, S.; Mégraud, F.; Vaillant, V.; Desenclos, J.C. Community Incidence of Campylobacteriosis and Nontyphoidal Salmonellosis, France, 2008–2013. Foodborne Pathog. Dis. 2015, 12, 664–669. [Google Scholar] [CrossRef] [PubMed]
- CNCH (Centre National de Référence des Campylobacters et des Helicobacters); SPF (Santé Publique France). Rapport de l’Année d’Exercice 2019; CNCH Report; CNCH: Paris, France, 2020; 64p. [Google Scholar]
- Denis, M.; Henrique, E.; Chidaine, B.; Tircot, A.; Bougeard, S.; Fravalo, P. Campylobacter from sows in farrow-to-finish pig farms: Risk indicators and genetic diversity. Vet. Microbiol. 2011, 154, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Allain, V.; Chemaly, M.; Laisney, M.J.; Rouxel, S.; Quesne, S.; Le Bouquin, S. Prevalence of and risk factors for Campylobacter colonisation in broiler flocks at the end of the rearing period in France. Br. Poult. Sci. 2014, 55, 452–459. [Google Scholar] [CrossRef]
- Thépault, A.; Méric, G.; Rivoal, K.; Pascoe, B.; Mageiros, L.; Touzain, F.; Rose, V.; Béven, V.; Chemaly, M.; Sheppard, S.K. Genome-wide identification of host-segregating epidemiological markers for source attribution in Campylobacter jejuni. Appl. Environ. Microbiol. 2017, 83, e03085–e030816. [Google Scholar] [CrossRef]
- Thépault, A.; Rose, V.; Queguiner, M.; Chemaly, M.; Rivoal, K. Dogs and Cats: Reservoirs for Highly Diverse Campylobacter jejuni and a Potential Source of Human Exposure. Animals 2020, 12, 838. [Google Scholar] [CrossRef]
- Hue, O.; Le Bouquin, S.; Laisney, M.J.; Allain, V.; Lalande, F.; Petetin, I.; Rouxel, S.; Quesne, S.; Gloaguen, P.Y.; Picherot, M.; et al. Prevalence of and risk factors for Campylobacter spp. contamination of broiler chicken carcasses at the slaughterhouse. Food Microbiol. 2010, 27, 992–999. [Google Scholar] [CrossRef] [PubMed]
- Fosse, J.; Seegers, H.; Magras, C. Prevalence and risk factors for bacterial food-borne zoonotic hazards in slaughter pigs: A review. Zoonosis Public Health 2009, 56, 429–454. [Google Scholar] [CrossRef] [PubMed]
- Thépault, A.; Poezevara, T.; Quesne, S.; Rose, V.; Chemaly, M.; Rivoal, K. Prevalence of Thermophilic Campylobacter in Cattle Production at Slaughterhouse Level in France and Link Between C. jejuni Bovine Strains and Campylobacteriosis. Front. Microbiol. 2018, 9, 471. [Google Scholar] [CrossRef] [PubMed]
- Cauvin, E.; Benoit, F.; Denis, M. Prevalence of Campylobacter spp. in cattle and sheep farms, in Normandy, France. In Proceedings of the IAFP Congress, Nantes, France, 24–26 April 2019. [Google Scholar]
- Yang, R.; Jacobson, C.; Gardner, G.; Carmichael, I.; Campbell, A.J.; Ryan, U. Longitudinal prevalence, faecal shedding and molecular characterisation of Campylobacter spp. and Salmonella enterica in sheep. Vet. J. 2014, 202, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Jones, K. Campylobacters in water, sewage and the environment. Symp. Ser. (Soc. Appl. Microbiol.) 2001, 30, 68S–79S. [Google Scholar] [CrossRef]
- Denis, M.; Tanguy, M.; Chidaine, B.; Laisney, M.J.; Mégraud, F.; Fravalo, P. Description and sources of contamination by Campylobacter spp. of river water destined for human consumption in Brittany, France. Pathol. Biol. 2011, 59, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Rincé, A.; Balière, C.; Hervio-Heath, D.; Cozien, J.; Lozach, S.; Parnaudeau, S.; Le Guyader, F.S.; Le Hello, S.; Giard, J.C.; Sauvageot, N.; et al. Occurrence of Bacterial Pathogens and Human Noroviruses in Shellfish-Harvesting Areas and Their Catchments in France. Front. Microbiol. 2018, 11, 2443. [Google Scholar] [CrossRef] [PubMed]
- Clark, C.G.; Taboada, E.; Grant, C.C.; Blakeston, C.; Pollari, F.; Marshall, B.; Rahn, K.; Mackinnon, J.; Daignault, D.; Pillai, D.; et al. Comparison of molecular typing methods useful for detecting clusters of Campylobacter jejuni and C. coli isolates through routine surveillance. J. Clin. Microbiol. 2012, 50, 798–809. [Google Scholar] [CrossRef]
- Jonas, R.; Kittl, S.; Overesch, G.; Kuhnert, P. Genotypes and antibiotic resistance of bovine Campylobacter and their contribution to human campylobacteriosis. Epidemiol. Infect. 2015, 143, 2373–2380. [Google Scholar] [CrossRef] [PubMed]
- Mughini-Gras, L.; Penny, C.; Ragimbeau, C.; Schets, F.M.; Blaak, H.; Duim, B.; Wagenaar, J.A.; de Boer, A.; Cauchie, H.M.; Mossong, J.; et al. Quantifying potential sources of surface water contamination with Campylobacter jejuni and Campylobacter coli. Water Res. 2016, 15, 36–45. [Google Scholar] [CrossRef]
- Mossong, J.; Mughini-Gras, L.; Penny, C.; Devaux, A.; Olinger, C.; Losch, S.; Cauchie, H.M.; van Pelt, W.; Ragimbeau, C. Human Campylobacteriosis in Luxembourg, 2010–2013: A Case-Control Study Combined with Multilocus Sequence Typing for Source Attribution and Risk Factor Analysis. Sci. Rep. 2016, 10, 20939. [Google Scholar] [CrossRef]
- Hsu, C.H.; Harrison, L.; Mukherjee, S.; Strain, E.; McDermott, P.; Zhang, Q.; Zhao, S. Core Genome Multilocus Sequence Typing for Food Animal Source Attribution of Human Campylobacter jejuni Infections. Pathogens 2020, 2, 532. [Google Scholar] [CrossRef]
- Mulder, A.C.; Franz, E.; de Rijk, S.; Versluis, M.A.J.; Coipan, C.; Buij, R.; Müskens, G.; Koene, M.; Pijnacker, R.; Duim, B.; et al. Tracing the animal sources of surface water contamination with Campylobacter jejuni and Campylobacter coli. Water Res. 2020, 15, 116421. [Google Scholar] [CrossRef]
- Zautner, A.E.; Masanta, W.O.; Tareen, A.M.; Weig, M.; Lugert, R.; Groß, U.; Bader, O. Discrimination of multilocus sequence typing-based Campylobacter jejuni subgroups by MALDI-TOF mass spectrometry. BMC Microbiol. 2013, 7, 247. [Google Scholar] [CrossRef]
- Feucherolles, M.; Nennig, M.; Becker, S.L.; Martiny, D.; Losch, S.; Penny, C.; Cauchie, H.M.; Ragimbeau, C. Investigation of MALDI-TOF Mass Spectrometry for Assessing the Molecular Diversity of Campylobacter jejuni and Comparison with MLST and cgMLST: A Luxembourg One-Health Study. Diagnostics 2021, 11, 1949. [Google Scholar] [CrossRef]
- Cody, A.J.; Bray, J.E.; Jolley, K.A.; McCarthy, N.D.; Maiden, M.C.J. Core Genome Multilocus Sequence Typing scheme for stable, comparative analyses of Campylobacter jejuni and C. coli human disease isolates. J. Clin. Microbiol. 2017, 55, 2086–2097. [Google Scholar] [CrossRef]
- Nennig, M.; Llarena, A.K.; Herold, M.; Mossong, J.; Penny, C.; Losch, S.; Tresse, O.; Ragimbeau, C. Investigating Major Recurring Campylobacter jejuni Lineages in Luxembourg Using Four Core or Whole Genome Sequencing Typing Schemes. Front. Cell. Infect. Microbiol. 2021, 8, 608020. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Wen, X.; Falush, D. Documentation for Structure Software, Version 2.3; University of Chicago: Chicago, IL, USA, 2010. [Google Scholar]
- Lévesque, S.; Fournier, E.; Carrier, N.; Frost, E.; Arbeit, R.D.; Michaud, S. Campylobacteriosis in urban versus rural areas: A case-case study integrated with molecular typing to validate risk factors and to attribute sources of infection. PLoS ONE 2013, 8, e83731. [Google Scholar] [CrossRef] [PubMed]
- McLure, A.; Smith, J.J.; Firestone, S.M.; Kirk, M.D.; French, N.; Fearnley, E.; Wallace, R.; Valcanis, M.; Bulach, D.; Moffatt, C.R.M.; et al. Source attribution of campylobacteriosis in Australia, 2017–2019. Risk Anal. 2023. online ahead of print. [Google Scholar] [CrossRef]
- Ogden, I.D.; Dallas, J.F.; MacRae, M.; Rotariu, O.; Reay, K.W.; Leitch, M.; Thomson, A.P.; Sheppard, S.K.; Maiden, M.; Forbes, K.J.; et al. Campylobacter excreted into the environment by animal sources: Prevalence, concentration shed, and host association. Foodborne Pathog. Dis. 2009, 6, 1161–1170. [Google Scholar] [CrossRef] [PubMed]
- Mughini-Gras, L.; Smid, J.H.; Wagenaar, J.A.; de Boer, A.G.; Havelaar, A.H.; Friesema, I.H.; French, N.P.; Busani, L.; van Pelt, W. Risk factors for campylobacteriosis of chicken, ruminant, and environmental origin: A combined case-control and source attribution analysis. PLoS ONE 2012, 7, e42599. [Google Scholar] [CrossRef] [PubMed]
- Dearlove, B.L.; Cody, A.J.; Pascoe, B.; Méric, G.; Wilson, D.J.; Sheppard, S.K. Rapid host switching in generalist Campylobacter strains erodes the signal for tracing human infections. ISME J. 2016, 10, 721–729. [Google Scholar] [CrossRef]
- Ocejo, M.; Oporto, B.; Hurtado, A. Occurrence of Campylobacter jejuni and Campylobacter coli in Cattle and Sheep in Northern Spain and Changes in Antimicrobial Resistance in Two Studies 10-years Apart. Pathogens 2019, 8, 98. [Google Scholar] [CrossRef]
- Ocejo, M.; Oporto, B.; Lavín, J.L.; Hurtado, A. Whole genome-based characterisation of antimicrobial resistance and genetic diversity in Campylobacter jejuni and Campylobacter coli from ruminants. Sci. Rep. 2021, 26, 8998. [Google Scholar] [CrossRef]
- Bianchini, V.; Borella, L.; Benedetti, V.; Parisi, A.; Miccolupo, A.; Santoro, E.; Recordati, C.; Luini, M. Prevalence in bulk tank milk and epidemiology of Campylobacter jejuni in dairy herds in Northern Italy. Appl. Environ. Microbiol. 2014, 80, 1832–1837. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Sippy, R.; Sahin, O.; Plummer, P.; Vidal, A.; Newell, D.; Zhang, Q. Genetic diversity and antimicrobial susceptibility of Campylobacter jejuni isolates associated with sheep abortion in the United States and Great Britain. J. Clin. Microbiol. 2014, 52, 1853–1861. [Google Scholar] [CrossRef]
- Yaeger, M.J.; Sahin, O.; Plummer, P.J.; Wu, Z.; Stasko, J.A.; Zhang, Q. The pathology of natural and experimentally induced Campylobacter jejuni abortion in sheep. J. Vet. Diagn. Investig. 2021, 33, 1096–1105. [Google Scholar] [CrossRef] [PubMed]
- Gülmez Sağlam, A.; Akça, D.; Çelebi, Ö.; Büyük, F.; Çelik, E.; Coşkun, M.R.; Şahin, M.; Otlu, S. Isolation and molecular identification of Campylobacter spp. from vaginal swab sample obtained from sheep herds with abort history. Kafkas Univ. Vet. Fak. Derg. 2019, 25, 697–701. [Google Scholar] [CrossRef]
- Guyard-Nicodème, M.; Rivoal, K.; Houard, E.; Rose, V.; Quesne, S.; Mourand, G.; Rouxel, S.; Kempf, I.; Guillier, L.; Gauchard, F.; et al. Prevalence and characterization of Campylobacter jejuni from chicken meat sold in French retail outlets. Int. J. Food Microbiol. 2015, 203, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Thépault, A.; Guyard-Nicodème, M.; Rose, V.; Quesne, S.; Queguiner, M.; Houard, E.; Mégraud, F.; Rivoal, K.; Chemaly, M. A representative overview of the genetic diversity and lipooligosaccharide sialylation in Campylobacter jejuni along the broiler production chain in France and its comparison with human isolates. Int. J. Food Microbiol. 2018, 2, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, S.K.; Dallas, J.F.; MacRae, M.; McCarthy, N.D.; Sproston, E.L.; Gormley, F.J.; Strachan, N.J.; Ogden, I.D.; Maiden, M.C.; Forbes, K.J. Campylobacter genotypes from food animals, environmental sources and clinical disease in Scotland 2005/6. Int. J. Food Microbiol. 2009, 31, 96–103. [Google Scholar] [CrossRef]
- Denis, M.; Nagard, B.; Rose, V.; Bourgouin, K.; Cutimbo, M.; Kerouanton, A. No clear differences between organic or conventional pig farms in the genetic diversity or virulence of Campylobacter coli isolates. Front. Microbiol. 2017, 8, 1016. [Google Scholar] [CrossRef] [PubMed]
- Thakur, S.; Morrow, W.E.; Funk, J.A.; Bahnson, P.B.; Gebreyes, W.A. Molecular epidemiologic investigation of Campylobacter coli in swine production systems, using multilocus sequence typing. Appl. Environ. Microbiol. 2006, 72, 5666–5669. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.G.; Englen, M.D.; Kathariou, S.; Wesley, I.V.; Wang, G.; Pittenger-Alley, L.; Siletz, R.M.; Muraoka, W.; Fedorka-Cray, P.J.; Mandrell, R.E. Identification of host-associated alleles by multilocus sequence typing of Campylobacter coli strains from food animals. Microbiology 2006, 152 Pt 1, 245–255. [Google Scholar] [CrossRef]
- Litrup, E.; Torpdahl, M.; Nielsen, E.M. Multilocus sequence typing performed on Campylobacter coli isolates from humans, broilers, pigs and cattle originating in Denmark. J. Appl. Microbiol. 2007, 103, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Egger, R.; Korczak, B.M.; Niederer, L.; Overesch, G.; Kuhnert, P. Genotypes and antibiotic resistance of Campylobacter coli in fattening pigs. Vet. Microbiol. 2012, 23, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Di Donato, G.; Marotta, F.; Nuvoloni, R.; Zilli, K.; Neri, D.; Di Sabatino, D.; Calistri, P.; Di Giannatale, E. Prevalence, Population Diversity and Antimicrobial Resistance of Campylobacter coli Isolated in Italian Swine at Slaughterhouse. Microorganisms 2020, 7, 222. [Google Scholar] [CrossRef] [PubMed]
- French, N.; Barrigas, M.; Brown, P.; Ribiero, P.; Williams, N.; Leatherbarrow, H.; Birtles, R.; Bolton, E.; Fearnhead, P.; Fox, A. Spatial epidemiology and natural population structure of Campylobacter jejuni colonizing a farmland ecosystem. Environ. Microbiol. 2005, 7, 1116–1126. [Google Scholar] [CrossRef] [PubMed]
- Sopwith, W.; Birtles, A.; Matthews, M.; Fox, A.; Gee, S.; Painter, M.; Regan, M.; Syed, Q.; Bolton, E. Identification of potential environmentally adapted Campylobacter jejuni strain, United Kingdom. Emerg. Infect. Dis. 2008, 14, 1769–1773. [Google Scholar] [CrossRef]
- Hock, L.; Herold, M.; Walczak, C.; Schoos, A.; Penny, C.; Cauchie, H.M.; Ragimbeau, C. Environmental dynamics of Campylobacter jejuni genotypes circulating in Luxembourg: What is the role of wild birds? Microb. Genom. 2023, 9, mgen001031. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sources | Pig | Cattle | Sheep | Poultry | River | Total |
|---|---|---|---|---|---|---|
| C. jejuni | 0 | 61 | 27 | 36 | 14 | 138 |
| C. coli | 60 | 0 | 15 | 29 | 21 | 125 |
| Total | 60 | 61 | 42 | 65 | 35 | 263 |
| Campylobacter jejuni | Campylobacter coli | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CC | ST | Cattle | Sheep | Poultry | River | Total | CC | ST | Pig | Sheep | Poultry | River | Total | |
| 21 | 19 | 14 | 7 | 1 | 1 | 23 | 828 | 825 | 2 | 3 | 2 | 7 | ||
| 21 | 8 | 5 | 3 | 16 | 828 | 2 | 2 | |||||||
| 50 | 2 | 2 | 829 | 1 | 1 | |||||||||
| 53 | 1 | 1 | 832 | 2 | 2 | |||||||||
| 22 | 22 | 3 | 3 | 854 | 15 | 3 | 4 | 22 | ||||||
| 42 | 42 | 7 | 7 | 860 | 2 | 2 | ||||||||
| 9760 | 2 | 2 | 890 | 1 | 1 | 2 | ||||||||
| 45 | 11 | 2 | 2 | 899 | 1 | 1 | ||||||||
| 45 | 3 | 1 | 3 | 7 | 1016 | 2 | 1 | 3 | ||||||
| 137 | 1 | 1 | 2 | 1061 | 1 | 1 | ||||||||
| 583 | 1 | 1 | 1096 | 2 | 2 | 4 | ||||||||
| 1701 | 1 | 1 | 1103 | 1 | 1 | |||||||||
| 6076 | 1 | 1 | 1107 | 4 | 4 | |||||||||
| 48 | 38 | 5 | 1 | 6 | 1142 | 1 | 1 | |||||||
| 48 | 1 | 1 | 1413 | 2 | 2 | |||||||||
| 52 | 52 | 5 | 5 | 1436 | 1 | 1 | ||||||||
| 61 | 61 | 9 | 9 | 1545 | 1 | 1 | ||||||||
| 179 | 6421 | 1 | 1 | 1556 | 2 | 2 | ||||||||
| 257 | 257 | 4 | 1 | 5 | 1583 | 1 | 1 | |||||||
| 824 | 1 | 1 | 1590 | 2 | 2 | |||||||||
| 2254 | 1 | 1 | 1834 | 2 | 2 | |||||||||
| 283 | 267 | 1 | 1 | 2183 | 1 | 1 | ||||||||
| 353 | 356 | 1 | 1 | 2642 | 1 | 1 | ||||||||
| 400 | 1 | 1 | 2699 | 2 | 8 | 1 | 11 | |||||||
| 2641 | 1 | 1 | 2733 | 1 | 1 | |||||||||
| 354 | 324 | 1 | 1 | 2741 | 2 | 2 | ||||||||
| 354 | 5 | 5 | 4044 | 1 | 1 | |||||||||
| 443 | 51 | 1 | 1 | 4086 | 1 | 1 | ||||||||
| 443 | 2 | 2 | 5306 | 2 | 2 | |||||||||
| 859 | 1 | 1 | 5959 | 1 | 1 | |||||||||
| 446 | 450 | 1 | 1 | 6538 | 1 | 1 | ||||||||
| 464 | 464 | 2 | 2 | 6688 | 1 | 1 | ||||||||
| 574 | 305 | 1 | 1 | 7168 | 1 | 1 | ||||||||
| 607 | 863 | 1 | 1 | 7285 | 1 | 1 | ||||||||
| 658 | 658 | 1 | 1 | 7286 | 1 | 1 | ||||||||
| 1044 | 1 | 1 | 7288 | 1 | 1 | |||||||||
| 952 | 7834 | 1 | 1 | 7289 | 1 | 1 | ||||||||
| 1275 | 3629 | 1 | 1 | 7292 | 1 | 1 | ||||||||
| NA | 586 | 1 | 1 | 7336 | 1 | 1 | ||||||||
| 1409 | 3 | 3 | 9631 | 1 | 1 | |||||||||
| 2274 | 1 | 1 | 9703 | 1 | 1 | |||||||||
| 2324 | 3 | 3 | 9704 | 1 | 1 | |||||||||
| 3766 | 1 | 1 | 9706 | 1 | 1 | |||||||||
| 4282 | 1 | 1 | NA | 1143 | 1 | 1 | ||||||||
| 4490 | 2 | 2 | 1450 | 1 | 1 | |||||||||
| 5053 | 1 | 1 | 1470 | 1 | 1 | |||||||||
| 5210 | 2 | 2 | 1577 | 1 | 1 | |||||||||
| 9632 | 2 | 2 | 1584 | 1 | 1 | |||||||||
| 9633 | 1 | 1 | 1764 | 2 | 2 | |||||||||
| Total | 61 | 27 | 36 | 14 | 138 | 1766 | 3 | 3 | ||||||
| ID | 0.89 | 0.84 | 0.97 | 0.95 | 0.94 | 1981 | 1 | 1 | ||||||
| 95% CI | 0.86–0.93 | 0.80–0.89 | 0.96–0.99 | 0.89–1.02 | 0.93–0.97 | 1990 | 1 | 1 | ||||||
| 1991 | 2 | 2 | ||||||||||||
| 1991 | 2 | 2 | ||||||||||||
| 4951 | 1 | 1 | ||||||||||||
| 5150 | 1 | 1 | ||||||||||||
| 7284 | 1 | 1 | ||||||||||||
| 8487 | 1 | 1 | ||||||||||||
| 9629 | 1 | 1 | ||||||||||||
| 9630 | 1 | 1 | ||||||||||||
| 9705 | 1 | 1 | ||||||||||||
| 9707 | 1 | 1 | ||||||||||||
| 9708 | 1 | 1 | ||||||||||||
| 9709 | 1 | 1 | ||||||||||||
| 9758 | 1 | 1 | ||||||||||||
| 9759 | 1 | 1 | ||||||||||||
| 9946 | 1 | 1 | ||||||||||||
| Total | 60 | 15 | 29 | 21 | 125 | |||||||||
| ID | 0.93 | 0.67 | 0.97 | 0.98 | 0.96 | |||||||||
| 95% CI | 0.88–0.98 | 0.49–0.86 | 0.95–1.00 | 0.95–1.01 | 0.93–0.98 | |||||||||
| (a) | MLST | MALDI ≥ 91% | MALDI ≥ 94% | MALDI ≥ 97% | ||||||||
| sources of C. jejuni strains | n. of isolates | n. of water strains with same ST as animal strains | % of attributed water strains | n. of water strains in same cluster ≥ 91% as animal strains | % of attributed water strains | n. of water strains in same cluster ≥ 94% as animal strains | % of attributed water strains | n. of water strains in same cluster ≥ 97% as animal strains | % of attributed water strains | |||
| Cattle | 61 | 5 | 35.7 | 11 | 78.6 | 6 | 42.8 | 2 | 14.3 | |||
| Sheep | 27 | 1 | 7.1 | 1 | 7.1 | 0 | 0 | 0 | 0 | |||
| Poultry | 36 | 4 | 28.5 | 12 | 85.7 | 4 | 14.3 | 1 | 7.1 | |||
| Water | 14 | 9 non-attributed | 64.3 | 1 non-attributed | 7.1 | 6 non-attributed | 42.8 | 11 non-attributed | 78.8 | |||
| (b) | MLST | MALDI ≥ 91% | MALDI ≥ 94% | MALDI ≥ 97% | ||||||||
| sources of C. coli strains | n. of isolates | n. of water strains with same ST as animal strains | % of attributed water strains | n. of water strains in same cluster ≥ 91% as animal strains | % of attributed water strains | n. of water strains in same cluster ≥ 94% as animal strains | % of attributed water strains | n. of water strains in same cluster ≥ 97% as animal strains | % of attributed water strains | |||
| Pig | 60 | 0 | 0 | 5 | 23.8 | 0 | 0 | 0 | 0 | |||
| Sheep | 15 | 0 | 0 | 5 | 23.8 | 0 | 0 | 0 | 0 | |||
| Poultry | 29 | 0 | 0 | 14 | 66.6 | 7 | 33.3 | 5 | 23.8 | |||
| Water | 21 | 21 non-attributed | 100 | 2 non-attributed | 9.52 | 14 non-attributed | 66.6 | 16 non-attributed | 76.2 | |||
| Sources | Cattle | Pig | Poultry | Sheep | Unsampled |
|---|---|---|---|---|---|
| C. jejuni (n = 14) | 0 | NA | 0.935 | 0.003 | 0.063 |
| C. coli (n = 21) | NA | 0 | 0.057 | 0 | 0.943 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Denis, M.; Rose, V.; Nagard, B.; Thépault, A.; Lucas, P.; Meunier, M.; Benoit, F.; Wilhem, A.; Gassilloud, B.; Cauvin, E.; et al. Comparative Analysis of Campylobacter jejuni and C. coli Isolated from Livestock Animals to C. jejuni and C. coli Isolated from Surface Water Using DNA Sequencing and MALDI-TOF. Pathogens 2023, 12, 1069. https://doi.org/10.3390/pathogens12091069
Denis M, Rose V, Nagard B, Thépault A, Lucas P, Meunier M, Benoit F, Wilhem A, Gassilloud B, Cauvin E, et al. Comparative Analysis of Campylobacter jejuni and C. coli Isolated from Livestock Animals to C. jejuni and C. coli Isolated from Surface Water Using DNA Sequencing and MALDI-TOF. Pathogens. 2023; 12(9):1069. https://doi.org/10.3390/pathogens12091069
Chicago/Turabian StyleDenis, Martine, Valérie Rose, Bérengère Nagard, Amandine Thépault, Pierrick Lucas, Meagan Meunier, Fabienne Benoit, Amandine Wilhem, Benoit Gassilloud, Elodie Cauvin, and et al. 2023. "Comparative Analysis of Campylobacter jejuni and C. coli Isolated from Livestock Animals to C. jejuni and C. coli Isolated from Surface Water Using DNA Sequencing and MALDI-TOF" Pathogens 12, no. 9: 1069. https://doi.org/10.3390/pathogens12091069