
Characterisation of a Staphylococcus aureus Isolate Carrying Phage-Borne Enterotoxin E from a European Badger (Meles meles)
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Case Report
2.2. Amplification, Labelling, and Microarray Analyses
2.3. Whole Genome Sequencing by Illumina and Oxford Nanopore Technology (ONT)
2.4. MLST and Spa Typing
2.5. Phage Induction and Phage DNA Preparation
2.6. Sequencing of Phage DNA Applying ONT
2.7. Bioinformatic Analysis of Sequencing Data
2.8. Phage Detection by Transmission Electron Microscopy (TEM)
3. Results
3.1. Typing and Strain Characteristics
3.2. Comparison to Other CC425 Sequences and Strains
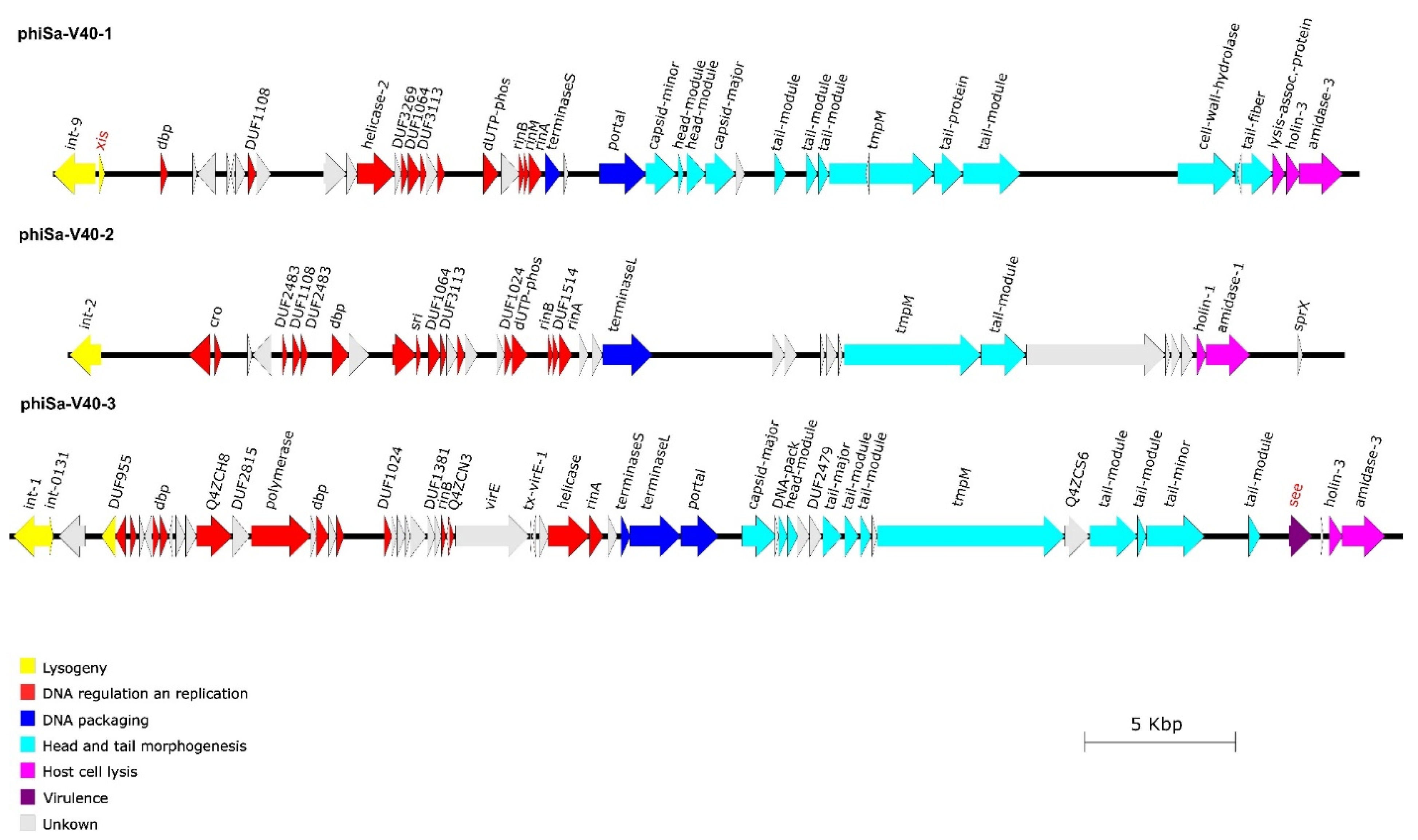
3.3. Phage Integration Sites and Sequences
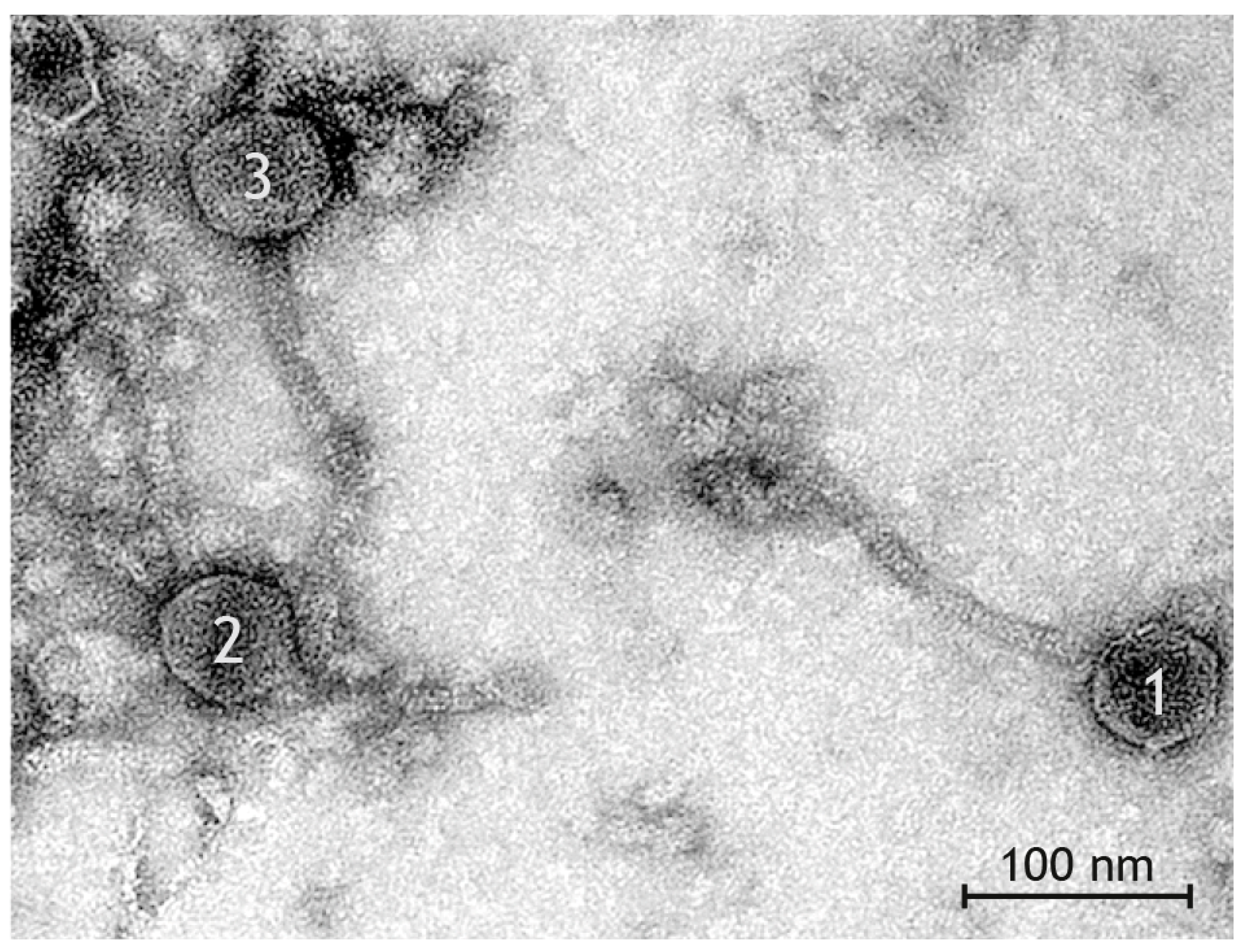
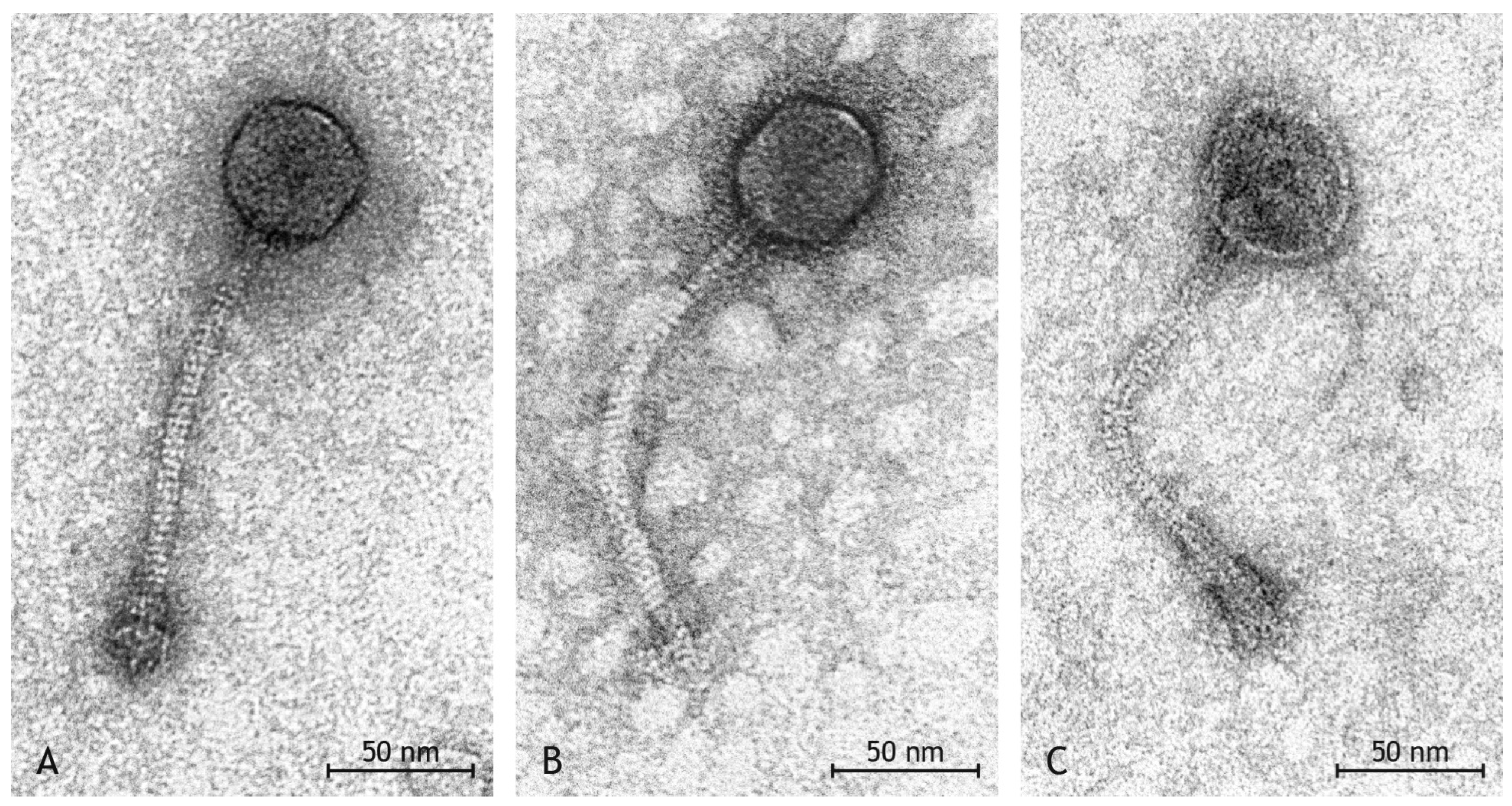
3.4. Phage Morphology
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waldvogel, F.A. Staphylococcus aureus (Including Toxic Shock Syndrome); Churchill Livingstone: New York, NY, USA; Edinburgh, Scotland; London, UK; Melbourne, Australia; Tokyo, Japan, 1995; pp. 1754–1777. [Google Scholar]
- Becker, K.; Skov, R.L.; von Eiff, C. Staphylococcus, Micrococcus and other catalase-positive cocci. In Manual of Clinical Microbiology, 11th ed.; Jorgensen, J.H., Pfaller, M.A., Carroll, K.C., Funke, G., Landry, M.L., Richter, S.S., Warnock, D.W., Eds.; ASM Press: Washington, DC, USA, 2015; Volume 1, pp. 354–382. [Google Scholar]
- Nowakiewicz, A.; Ziolkowska, G.; Zieba, P.; Gnat, S.; Wojtanowicz-Markiewicz, K.; Troscianczyk, A. Coagulase-positive Staphylococcus isolated from wildlife: Identification, molecular characterization and evaluation of resistance profiles with focus on a methicillin-resistant strain. Comp. Immunol. Microbiol. Infect. Dis. 2016, 44, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Loncaric, I.; Kubber-Heiss, A.; Posautz, A.; Stalder, G.L.; Hoffmann, D.; Rosengarten, R.; Walzer, C. Characterization of methicillin-resistant Staphylococcus spp. carrying the mecC gene, isolated from wildlife. J. Antimicrob. Chemother. 2013, 68, 2222–2225. [Google Scholar] [CrossRef] [PubMed]
- Loncaric, I.; Kubber-Heiss, A.; Posautz, A.; Stalder, G.L.; Hoffmann, D.; Rosengarten, R.; Walzer, C. mecC- and mecA-positive meticillin-resistant Staphylococcus aureus (MRSA) isolated from livestock sharing habitat with wildlife previously tested positive for mecC-positive MRSA. Vet. Dermatol. 2014, 25, 147–148. [Google Scholar] [CrossRef] [PubMed]
- Monecke, S.; Gavier-Widén, D.; Hotzel, H.; Peters, M.; Guenther, S.; Lazaris, A.; Loncaric, I.; Müller, E.; Reissig, A.; Ruppelt-Lorz, A.; et al. Diversity of Staphylococcus aureus Isolates in European Wildlife. PLoS ONE 2016, 11, e0168433. [Google Scholar] [CrossRef] [PubMed]
- Wardyn, S.E.; Kauffman, L.K.; Smith, T.C. Methicillin-resistant Staphylococcus aureus in central Iowa wildlife. J. Wildl. Dis. 2012, 48, 1069–1073. [Google Scholar] [CrossRef] [PubMed]
- Simpson, V.R.; Davison, N.J.; Kearns, A.M.; Pichon, B.; Hudson, L.O.; Koylass, M.; Blackett, T.; Butler, H.; Rasigade, J.P.; Whatmore, A.M. Association of a lukM-positive clone of Staphylococcus aureus with fatal exudative dermatitis in red squirrels (Sciurus vulgaris). Vet. Microbiol. 2013, 162, 987–991. [Google Scholar] [CrossRef]
- Simpson, V.R.; Hargreaves, J.; Everest, D.J.; Baker, A.S.; Booth, P.A.; Butler, H.M.; Blackett, T. Mortality in red squirrels (Sciurus vulgaris) associated with exudative dermatitis. Vet. Rec. 2010, 167, 59–62. [Google Scholar] [CrossRef]
- Sousa, M.; Silva, N.; Igrejas, G.; Silva, F.; Sargo, R.; Alegria, N.; Benito, D.; Gomez, P.; Lozano, C.; Gomez-Sanz, E.; et al. Antimicrobial resistance determinants in Staphylococcus spp. recovered from birds of prey in Portugal. Vet. Microbiol. 2014, 171, 436–440. [Google Scholar] [CrossRef]
- Gómez, P.; González-Barrio, D.; Benito, D.; García, J.T.; Viñuela, J.; Zarazaga, M.; Ruiz-Fons, F.; Torres, C. Detection of methicillin-resistant Staphylococcus aureus (MRSA) carrying the mecC gene in wild small mammals in Spain. J. Antimicrob. Chemother. 2014, 69, 2061–2064. [Google Scholar] [CrossRef]
- Price, L.B.; Stegger, M.; Hasman, H.; Aziz, M.; Larsen, J.; Andersen, P.S.; Pearson, T.; Waters, A.E.; Foster, J.; Schupp, J.; et al. Staphylococcus aureus CC398: Host adaptation and emergence of methicillin resistance in livestock. MBio, 2012; in press. [Google Scholar]
- Wagenaar, J.; Yue, H.; Pritchard, J.; Broekhuizen-Stins, M.; Huijsdens, X.; Mevius, D. Unexpected sequence types in livestock associated methicillin-resistant Staphylococcus aureus (MRSA): MRSA ST9 and a single locus variant of ST9 in pig farming in China. Vet. Microbiol. 2009, 139, 405–409. [Google Scholar] [CrossRef]
- Wulf, M.; Voss, A. MRSA in livestock animals-an epidemic waiting to happen? Clin. Microbiol. Infect. 2008, 14, 519–521. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Ballhausen, B.; Kahl, B.C.; Kock, R. The clinical impact of livestock-associated methicillin-resistant Staphylococcus aureus of the clonal complex 398 for humans. Vet. Microbiol. 2017, 200, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Feßler, A.T.; Kadlec, K.; Hassel, M.; Hauschild, T.; Eidam, C.; Ehricht, R.; Monecke, S.; Schwarz, S. Characterization of methicillin-resistant Staphylococcus aureus isolates from food and food products of poultry origin in Germany. Appl. Environ. Microbiol. 2011, 77, 7151–7157. [Google Scholar] [CrossRef]
- Lowder, B.V.; Guinane, C.M.; Ben Zakour, N.L.; Weinert, L.A.; Conway-Morris, A.; Cartwright, R.A.; Simpson, A.J.; Rambaut, A.; Nübel, U.; Fitzgerald, J.R. Recent human-to-poultry host jump, adaptation, and pandemic spread of Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2009, 106, 19545–19550. [Google Scholar] [CrossRef]
- Feßler, A.T.; Thomas, P.; Mühldorfer, K.; Grobbel, M.; Brombach, J.; Eichhorn, I.; Monecke, S.; Ehricht, R.; Schwarz, S. Phenotypic and genotypic characteristics of Staphylococcus aureus isolates from zoo and wild animals. Vet. Microbiol. 2018, 218, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Gnat, S.; Troscianczyk, A.; Nowakiewicz, A.; Majer-Dziedzic, B.; Ziolkowska, G.; Dziedzic, R.; Zieba, P.; Teodorowski, O. Experimental studies of microbial populations and incidence of zoonotic pathogens in the faeces of red deer (Cervus elaphus). Lett. Appl. Microbiol. 2015, 61, 446–452. [Google Scholar] [CrossRef]
- Abdullahi, I.N.; Fernández-Fernández, R.; Juárez-Fernández, G.; Martínez-Álvarez, S.; Eguizábal, P.; Zarazaga, M.; Lozano, C.; Torres, C. Wild Animals Are Reservoirs and Sentinels of Staphylococcus aureus and MRSA Clones: A Problem with “One Health” Concern. Antibiotics 2021, 10, 1556. [Google Scholar] [CrossRef]
- Bengtsson, B.; Persson, L.; Ekström, K.; Unnerstad, H.E.; Uhlhorn, H.; Börjesson, S. High occurrence of mecC-MRSA in wild hedgehogs (Erinaceus europaeus) in Sweden. Vet. Microbiol. 2017, 207, 103–107. [Google Scholar] [CrossRef]
- Monecke, S.; Gavier-Widen, D.; Mattsson, R.; Rangstrup-Christensen, L.; Lazaris, A.; Coleman, D.C.; Shore, A.C.; Ehricht, R. Detection of mecC-positive Staphylococcus aureus (CC130-MRSA-XI) in diseased European hedgehogs (Erinaceus europaeus) in Sweden. PLoS ONE 2013, 8, e66166. [Google Scholar] [CrossRef]
- Rasmussen, S.L.; Larsen, J.; van Wijk, R.E.; Jones, O.R.; Berg, T.B.; Angen, Ø.; Larsen, A.R. European hedgehogs (Erinaceus europaeus) as a natural reservoir of methicillin-resistant Staphylococcus aureus carrying mecC in Denmark. PLoS ONE 2019, 14, e0222031. [Google Scholar] [CrossRef]
- Smith, J.M.; Marples, M.J. Dermatophyte lesions in the hedgehog as a reservoir of penicillin-resistant staphylococci. J. Hyg. 1965, 63, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.; Raisen, C.L.; Ba, X.; Sadgrove, N.J.; Padilla-González, G.F.; Simmonds, M.S.J.; Loncaric, I.; Kerschner, H.; Apfalter, P.; Hartl, R.; et al. Emergence of methicillin resistance predates the clinical use of antibiotics. Nature 2022, 602, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Dube, F.; Söderlund, R.; Lampinen Salomonsson, M.; Troell, K.; Börjesson, S. Benzylpenicillin-producing Trichophyton erinacei and methicillin resistant Staphylococcus aureus carrying the mecC gene on European hedgehogs—A pilot-study. BMC Microbiol. 2021, 21, 212. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, J.; Kamio, Y. Bacterial two-component and hetero-heptameric pore-forming cytolytic toxins: Structures, pore-forming mechanism, and organization of the genes. Biosci. Biotechnol. Biochem. 2004, 68, 981–1003. [Google Scholar] [CrossRef]
- Dinges, M.M.; Orwin, P.M.; Schlievert, P.M. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 2000, 13, 16–34. [Google Scholar] [CrossRef]
- Monecke, S.; Feßler, A.T.; Burgold-Voigt, S.; Krüger, H.; Mühldorfer, K.; Wibbelt, G.; Liebler-Tenorio, E.M.; Reinicke, M.; Braun, S.D.; Hanke, D.; et al. Staphylococcus aureus isolates from Eurasian Beavers (Castor fiber) carry a novel phage-borne bicomponent leukocidin related to the Panton-Valentine leukocidin. Sci. Rep. 2021, 11, 24394. [Google Scholar] [CrossRef]
- Sabat, A.J.; Wouthuyzen-Bakker, M.; Rondags, A.; Hughes, L.; Akkerboom, V.; Koutsopetra, O.; Friedrich, A.W.; Bathoorn, E. Case Report: Necrotizing fasciitis caused by Staphylococcus aureus positive for a new sequence variant of exfoliative toxin E. Front. Genet. 2022, 13, 964358. [Google Scholar] [CrossRef]
- Kaneko, J.; Kimura, T.; Kawakami, Y.; Tomita, T.; Kamio, Y. Panton-valentine leukocidin genes in a phage-like particle isolated from mitomycin C-treated Staphylococcus aureus V8 (ATCC 49775). Biosci. Biotechnol. Biochem. 1997, 61, 1960–1962. [Google Scholar] [CrossRef]
- Kaneko, J.; Kimura, T.; Narita, S.; Tomita, T.; Kamio, Y. Complete nucleotide sequence and molecular characterization of the temperate staphylococcal bacteriophage phiPVL carrying Panton-Valentine leukocidin genes. Gene 1998, 215, 57–67. [Google Scholar] [CrossRef]
- Coleman, D.C.; Sullivan, D.J.; Russell, R.J.; Arbuthnott, J.P.; Carey, B.F.; Pomeroy, H.M. Staphylococcus aureus bacteriophages mediating the simultaneous lysogenic conversion of beta-lysin, staphylokinase and enterotoxin A: Molecular mechanism of triple conversion. J. Gen. Microbiol. 1989, 135, 1679–1697. [Google Scholar] [CrossRef]
- Skippington, E.; Ragan, M.A. Lateral genetic transfer and the construction of genetic exchange communities. FEMS Microbiol. Rev. 2011, 35, 707–735. [Google Scholar] [CrossRef] [PubMed]
- McNally, L.; Viana, M.; Brown, S.P. Cooperative secretions facilitate host range expansion in bacteria. Nat. Commun. 2014, 5, 4594. [Google Scholar] [CrossRef] [PubMed]
- Haaber, J.; Penadés, J.R.; Ingmer, H. Transfer of Antibiotic Resistance in Staphylococcus aureus. Trends Microbiol. 2017, 25, 893–905. [Google Scholar] [CrossRef] [PubMed]
- Koop, G.; Vrieling, M.; Storisteanu, D.M.; Lok, L.S.; Monie, T.; van Wigcheren, G.; Raisen, C.; Ba, X.; Gleadall, N.; Hadjirin, N.; et al. Identification of LukPQ, a novel, equid-adapted leukocidin of Staphylococcus aureus. Sci. Rep. 2017, 7, 40660. [Google Scholar] [CrossRef]
- Vancraeynest, D.; Haesebrouck, F.; Deplano, A.; Denis, O.; Godard, C.; Wildemauwe, C.; Hermans, K. International dissemination of a high virulence rabbit Staphylococcus aureus clone. J. Vet. Med. B Infect. Dis. Vet. Public Health 2006, 53, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Alvarez, L.; Holden, M.T.; Lindsay, H.; Webb, C.R.; Brown, D.F.; Curran, M.D.; Walpole, E.; Brooks, K.; Pickard, D.J.; Teale, C.; et al. Meticillin-resistant Staphylococcus aureus with a novel mecA homologue in human and bovine populations in the UK and Denmark: A descriptive study. Lancet Infect. Dis. 2011, 11, 595–603. [Google Scholar] [CrossRef]
- Meemken, D.; Blaha, T.; Hotzel, H.; Strommenger, B.; Klein, G.; Ehricht, R.; Monecke, S.; Kehrenberg, C. Genotypic and Phenotypic Characterization of Staphylococcus aureus Isolates from Wild Boars. Appl. Environ. Microbiol. 2013, 79, 1739–1742. [Google Scholar] [CrossRef]
- Concepcion Porrero, M.; Harrison, E.M.; Fernandez-Garayzabal, J.F.; Paterson, G.K.; Diez-Guerrier, A.; Holmes, M.A.; Dominguez, L. Detection of mecC-Methicillin-resistant Staphylococcus aureus isolates in river water: A potential role for water in the environmental dissemination. Environ. Microbiol. Rep. 2014, 6, 705–708. [Google Scholar] [CrossRef] [PubMed]
- Porrero, M.C.; Mentaberre, G.; Sánchez, S.; Fernández-Llario, P.; Casas-Díaz, E.; Mateos, A.; Vidal, D.; Lavín, S.; Fernández-Garayzábal, J.-F.; Domínguez, L. Carriage of Staphylococcus aureus by Free-Living Wild Animals in Spain. Appl. Environ. Microbiol. 2014, 80, 4865–4870. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ripa, L.; Gómez, P.; Alonso, C.A.; Camacho, M.C.; de la Puente, J.; Fernández-Fernández, R.; Ramiro, Y.; Quevedo, M.A.; Blanco, J.M.; Zarazaga, M.; et al. Detection of MRSA of Lineages CC130-mecC and CC398-mecA and Staphylococcus delphini-lnu(A) in Magpies and Cinereous Vultures in Spain. Microb. Ecol. 2019, 78, 409–415. [Google Scholar] [CrossRef]
- Heaton, C.J.; Gerbig, G.R.; Sensius, L.D.; Patel, V.; Smith, T.C. Staphylococcus aureus Epidemiology in Wildlife: A Systematic Review. Antibiotics 2020, 9, 89. [Google Scholar] [CrossRef] [PubMed]
- Luzzago, C.; Lauzi, S.; Ehricht, R.; Monecke, S.; Corlatti, L.; Pedrotti, L.; Piccinini, R. Survey of Staphylococcus aureus carriage by free-living red deer (Cervus elaphus): Evidence of human and domestic animal lineages. Transbound. Emerg. Dis. 2022, 69, e1659–e1669. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, Q.; Zhang, H.; Li, X.; Huang, W.; Fu, Q.; Li, M. Molecular Characteristics of Community-Associated Staphylococcus aureus Isolates From Pediatric Patients With Bloodstream Infections Between 2012 and 2017 in Shanghai, China. Front. Microbiol. 2018, 9, 1211. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, G.; Monecke, S.; Brus, O.; Ehricht, R.; Soderquist, B. Long term molecular epidemiology of methicillin-susceptible Staphylococcus aureus bacteremia isolates in Sweden. PLoS ONE 2014, 9, e114276. [Google Scholar] [CrossRef]
- Damasco, P.V.; Chamon, R.C.; Barbosa, A.T.; da Cunha, S.; Aquino, J.H.; Cavalcante, F.S.; Dos Santos, K.R. Involvement of methicillin-susceptible Staphylococcus aureus related to sequence type 25 and harboring pvl genes in a case of carotid cavernous fistula after community-associated sepsis. J. Clin. Microbiol. 2012, 50, 196–198. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Seijas, C.; Mascarós, P.; Lizana, V.; Martí-Marco, A.; Arnau-Bonachera, A.; Chillida-Martínez, E.; Cardells, J.; Selva, L.; Viana, D.; Corpa, J.M. Genomic Characterization of Staphylococcus aureus in Wildlife. Animals 2023, 13, 1064. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, M.J.H.; Pascual-Linaza, A.V.; Couzens, C.; Holmes, C.; Bell, C.; Spence, N.; Huey, R.J.; Murphy, J.A.; Devaney, R.; Lahuerta-Marin, A. Estimation of the Prevalence of Antimicrobial Resistance in Badgers (Meles meles) and Foxes (Vulpes vulpes) in Northern Ireland. Front. Microbiol. 2021, 12, 596891. [Google Scholar] [CrossRef]
- Cheeseman, C.L.; Wilesmith, J.W.; Stuart, F.A. Tuberculosis: The disease and its epidemiology in the badger, a review. Epidemiol. Infect. 1989, 103, 113–125. [Google Scholar] [CrossRef]
- Bennett, R.M. The political economy of bovine tuberculosis in Great Britain. Rev. Sci. Tech. 2017, 36, 105–114. [Google Scholar] [CrossRef]
- Otsuji, N.; Sekiguchi, M.; Iijima, T.; Takagi, Y. Induction of phage formation in the lysogenic Escherichia coli K-12 by mitomycin C. Nature 1959, 184 (Suppl. 14), 1079–1080. [Google Scholar] [CrossRef]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, D.; Claus, H.; Witte, W.; Rothgänger, J.; Claus, H.; Turnwald, D.; Vogel, U. Typing of methicillin-mesistant Staphylococcus aureus in a university hospital setting by using novel software for spa repeat determination and database management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef] [PubMed]
- Raya, R.R.; H’Bert, E.M. Isolation of Phage via Induction of Lysogens. Methods Mol. Biol. 2009, 501, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Burgold-Voigt, S.; Monecke, S.; Simbeck, A.; Holzmann, T.; Kieninger, B.; Liebler-Tenorio, E.M.; Braun, S.D.; Collatz, M.; Diezel, C.; Müller, E.; et al. Characterisation and molecular analysis of an unusual chimeric methicillin resistant Staphylococcus aureus strain and its bacteriophages. Front. Genet. 2021, 12, 723958. [Google Scholar] [CrossRef] [PubMed]
- Hagemann, R. Extraktion der Lambda-DNA. In Gentechnologische Arbeitsmethoden—Ein Handbuch; Hagemann, R., Ed.; Akademie-Verlag: Berlin, Germany, 1990; p. 51. [Google Scholar]
- Crovadore, J.; Calmin, G.; Tonacini, J.; Chablais, R.; Baumgartner, A.; Schnyder, B.; Hodille, E.; Lefort, F. Whole-Genome Sequences of 15 Strains of Staphylococcus aureus subsp. aureus Isolated from Foodstuff and Human Clinical Samples. Genome Announc. 2015, 3, e00684-15. [Google Scholar] [CrossRef]
- Luzzago, C.; Lauzi, S.; Ehricht, R.; Monecke, S.; Trogu, T.; Pedrotti, L.; Lanfranchi, P.; Piccinini, R. Staphylococcus aureus nasal and intestinal carriage by free-ranging red deer: Evidence of human, domestic and wild animal lineages. International. J. Infect. Dis. 2019, 79, 21–22. [Google Scholar] [CrossRef]
- Shore, A.C.; Deasy, E.C.; Slickers, P.; Brennan, G.; O’Connell, B.; Monecke, S.; Ehricht, R.; Coleman, D.C. Detection of staphylococcal cassette chromosome mec type XI carrying highly divergent mecA, mecI, mecR1, blaZ, and ccr genes in human clinical isolates of clonal complex 130 methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 3765–3773. [Google Scholar] [CrossRef]
- Deghorain, M.; Van Melderen, L. The Staphylococci phages family: An overview. Viruses 2012, 4, 3316–3335. [Google Scholar] [CrossRef]
- Coleman, D.; Knights, J.; Russell, R.; Shanley, D.; Birkbeck, T.H.; Dougan, G.; Charles, I. Insertional inactivation of the Staphylococcus aureus beta-toxin by bacteriophage phi 13 occurs by site- and orientation-specific integration of the phi 13 genome. Mol. Microbiol. 1991, 5, 933–939. [Google Scholar] [CrossRef]
- van Wamel, W.J.B.; Rooijakkers, S.H.M.; Ruyken, M.; van Kessel, K.P.M.; van Strijp, J.A.G. The Innate Immune Modulators Staphylococcal Complement Inhibitor and Chemotaxis Inhibitory Protein of Staphylococcus aureus Are Located on {beta}-Hemolysin-Converting Bacteriophages. J. Bacteriol. 2006, 188, 1310–1315. [Google Scholar] [CrossRef]
- Sanchini, A.; Del Grosso, M.; Villa, L.; Ammendolia, M.G.; Superti, F.; Monaco, M.; Pantosti, A. Typing of Panton-Valentine leukocidin-encoding phages carried by methicillin-susceptible and methicillin-resistant Staphylococcus aureus from Italy. Clin. Microbiol. Infect. 2014, 20, O840–O846. [Google Scholar] [CrossRef] [PubMed]
- Dokland, T. Molecular Piracy: Redirection of Bacteriophage Capsid Assembly by Mobile Genetic Elements. Viruses 2019, 11, 1003. [Google Scholar] [CrossRef]
- Bar-On, Y.M.; Phillips, R.; Milo, R. The biomass distribution on Earth. Proc. Natl. Acad. Sci. USA 2018, 115, 6506–6511. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.H.; Banyard, A.C.; Hussein, A.; Laurenson, M.K.; Malcolm, J.R.; Marino, J.; Regassa, F.; Stewart, A.M.; Fooks, A.R.; Sillero-Zubiri, C. Canine distemper in endangered Ethiopian wolves. Emerg. Infect. Dis. 2015, 21, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Katale, B.Z.; Mbugi, E.V.; Siame, K.K.; Keyyu, J.D.; Kendall, S.; Kazwala, R.R.; Dockrell, H.M.; Fyumagwa, R.D.; Michel, A.L.; Rweyemamu, M.; et al. Isolation and Potential for Transmission of Mycobacterium bovis at Human-livestock-wildlife Interface of the Serengeti Ecosystem, Northern Tanzania. Transbound. Emerg. Dis. 2017, 64, 815–825. [Google Scholar] [CrossRef]
- Michel, A.L.; Bengis, R.G.; Keet, D.F.; Hofmeyr, M.; Klerk, L.M.; Cross, P.C.; Jolles, A.E.; Cooper, D.; Whyte, I.J.; Buss, P.; et al. Wildlife tuberculosis in South African conservation areas: Implications and challenges. Vet. Microbiol. 2006, 112, 91–100. [Google Scholar] [CrossRef]
- Bergdoll, M.S.; Borja, C.R.; Robbins, R.N.; Weiss, K.F. Identification of enterotoxin E. Infect. Immun. 1971, 4, 593–595. [Google Scholar] [CrossRef]
- Ostyn, A.; De Buyser, M.L.; Guillier, F.; Groult, J.; Felix, B.; Salah, S.; Delmas, G.; Hennekinne, J.A. First evidence of a food poisoning outbreak due to staphylococcal enterotoxin type E, France, 2009. Eurosurveillance 2010, 15, 19528. [Google Scholar] [CrossRef]
- Morris, C.A.; Conway, H.D.; Everall, P.H. Food-poisoning due to staphylococcal enterotoxin E. Lancet 1972, 2, 1375–1376. [Google Scholar] [CrossRef]
- Monecke, S.; Coombs, G.; Shore, A.C.; Coleman, D.C.; Akpaka, P.; Borg, M.; Chow, H.; Ip, M.; Jatzwauk, L.; Jonas, D.; et al. A field guide to pandemic, epidemic and sporadic clones of methicillin-resistant Staphylococcus aureus. PLoS ONE 2011, 6, e17936. [Google Scholar] [CrossRef]
- Grispoldi, L.; Massetti, L.; Sechi, P.; Iulietto, M.F.; Ceccarelli, M.; Karama, M.; Popescu, P.A.; Pandolfi, F.; Cenci-Goga, B.T. Short communication: Characterization of enterotoxin-producing Staphylococcus aureus isolated from mastitic cows. J. Dairy Sci. 2019, 102, 1059–1065. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhang, M.; Li, H.; Yang, H.; Li, X.; Song, X.; Wang, Z. Prevalence and molecular characterization of Staphylococcus aureus isolated from goats in Chongqing, China. BMC Vet. Res. 2017, 13, 352. [Google Scholar] [CrossRef] [PubMed]
- Bobay, L.M.; Touchon, M.; Rocha, E.P. Pervasive domestication of defective prophages by bacteria. Proc. Natl. Acad. Sci. USA 2014, 111, 12127–12132. [Google Scholar] [CrossRef] [PubMed]
- Dawkins, R. Das Egoistische Gen (The Selfish Gene, German Ed.); Spektrum Verlag: Berlin/Heidelberg, Germany, 1994. [Google Scholar]
- Peacock, S.J.; Moore, C.E.; Justice, A.; Kantzanou, M.; Story, L.; Mackie, K.; O’Neill, G.; Day, N.P. Virulent combinations of adhesin and toxin genes in natural populations of Staphylococcus aureus. Infect. Immun. 2002, 70, 4987–4996. [Google Scholar] [CrossRef] [PubMed]
- Goerke, C.; Köller, J.; Wolz, C. Ciprofloxacin and trimethoprim cause phage induction and virulence modulation in Staphylococcus aureus. Antimicrob. Agents Chemother. 2006, 50, 171–177. [Google Scholar] [CrossRef]
- Zeman, M.; Mašlaňová, I.; Indráková, A.; Šiborová, M.; Mikulášek, K.; Bendíčková, K.; Plevka, P.; Vrbovská, V.; Zdráhal, Z.; Doškař, J.; et al. Staphylococcus sciuri bacteriophages double-convert for staphylokinase and phospholipase, mediate interspecies plasmid transduction, and package mecA gene. Sci. Rep. 2017, 7, 46319. [Google Scholar] [CrossRef]
- Sutcliffe, S.G.; Shamash, M.; Hynes, A.P.; Maurice, C.F. Common Oral Medications Lead to Prophage Induction in Bacterial Isolates from the Human Gut. Viruses 2021, 13, 455. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Group | Head Shape | Head Size | Tail Length | Tail Diameter | Base Plate Size | Number in Preparation |
|---|---|---|---|---|---|---|
| I | icosahedral | Ø 57 nm × 56 nm | Ø 176 nm | Ø 10 nm | Ø 31 nm × 24 nm | 24 |
| II | elongated | Ø 62 nm × 52 nm | Ø 195 nm | Ø 10 nm | Ø 32 nm × 24 nm | 6 |
| III | broad | Ø 57 nm × 66 nm | Ø 157 nm | Ø 9 nm | Ø 30 nm × 20 nm | 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burgold-Voigt, S.; Monecke, S.; Busch, A.; Bocklisch, H.; Braun, S.D.; Diezel, C.; Hotzel, H.; Liebler-Tenorio, E.M.; Müller, E.; Reinicke, M.; et al. Characterisation of a Staphylococcus aureus Isolate Carrying Phage-Borne Enterotoxin E from a European Badger (Meles meles). Pathogens 2023, 12, 704. https://doi.org/10.3390/pathogens12050704
Burgold-Voigt S, Monecke S, Busch A, Bocklisch H, Braun SD, Diezel C, Hotzel H, Liebler-Tenorio EM, Müller E, Reinicke M, et al. Characterisation of a Staphylococcus aureus Isolate Carrying Phage-Borne Enterotoxin E from a European Badger (Meles meles). Pathogens. 2023; 12(5):704. https://doi.org/10.3390/pathogens12050704
Chicago/Turabian StyleBurgold-Voigt, Sindy, Stefan Monecke, Anne Busch, Herbert Bocklisch, Sascha D. Braun, Celia Diezel, Helmut Hotzel, Elisabeth M. Liebler-Tenorio, Elke Müller, Martin Reinicke, and et al. 2023. "Characterisation of a Staphylococcus aureus Isolate Carrying Phage-Borne Enterotoxin E from a European Badger (Meles meles)" Pathogens 12, no. 5: 704. https://doi.org/10.3390/pathogens12050704