
Infection with Seasonal H1N1 Influenza Results in Comparable Disease Kinetics and Host Immune Responses in Ferrets and Golden Syrian Hamsters

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Study Design
3.2. Comparison of Disease Induced by A/Guangdong-Maonan/SW1536/2019 (GM19)
3.3. Comparison of the Host Response Induced by A/Guangdong-Maonan/SW1536/2019 (GM19) Infection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Influenza (Seasonal): WHO; 2023 [updated 12/01/2023; cited 2023 23/02/2023]. Available online: https://www.who.int/news-room/fact-sheets/detail/influenza-(seasonal) (accessed on 23 February 2023).
- Bouvier, N.M.; Lowen, A.C. Animal Models for Influenza Virus Pathogenesis and Transmission. Viruses 2010, 2, 1530. [Google Scholar] [CrossRef]
- Park, M.S.; Kim, J.I.; Bae, J.-Y.; Park, M.-S. Animal models for the risk assessment of viral pandemic potential. Lab. Anim. Res. 2020, 36, 11. [Google Scholar] [CrossRef] [PubMed]
- Iwatsuki-Horimoto, K.; Nakajima, N.; Ichiko, Y.; Sakai-Tagawa, Y.; Noda, T.; Hasegawa, H.; Kawaoka, Y. Syrian Hamster as an Animal Model for the Study of Human Influenza Virus Infection. J. Virol. 2018, 92, e01693-17. [Google Scholar] [CrossRef]
- Fan, S.; Gu, C.; Kong, H.; Guan, L.; Neumann, G.; Kawaoka, Y. Influenza Viruses Suitable for Studies in Syrian Hamsters. Viruses 2022, 14, 1629. [Google Scholar] [CrossRef]
- Roubidoux, E.K.; Schultz-Cherry, S. Animal Models Utilized for the Development of Influenza Virus Vaccines. Vaccines 2021, 9, 787. [Google Scholar] [CrossRef]
- Couzens, L.; Gao, J.; Westgeest, K.; Sandbulte, M.; Lugovtsev, V.; Fouchier, R.; Eichelberger, M. An optimized enzyme-linked lectin assay to measure influenza A virus neuraminidase inhibition antibody titers in human sera. J. Virol. Methods 2014, 210, 7–14. [Google Scholar] [CrossRef]
- Ryan, K.A.; Slack, G.S.; Marriott, A.; Kane, J.A.; Whittaker, C.J.; Silman, N.J.; Carroll, M.W.; Gooch, K.E. Cellular immune response to human influenza viruses differs between H1N1 and H3N2 subtypes in the ferret lung. PLoS ONE 2018, 13, e0202675. [Google Scholar] [CrossRef]
- Morgan, S.B.; Hemmink, J.D.; Porter, E.; Harley, R.; Shelton, H.; Aramouni, M.; Everett, H.E.; Brookes, S.M.; Bailey, M.; Townsend, A.M.; et al. Aerosol Delivery of a Candidate Universal Influenza Vaccine Reduces Viral Load in Pigs Challenged with Pandemic H1N1 Virus. J. Immunol. 2016, 196, 5014–5023. [Google Scholar] [CrossRef]
- Gauger, P.C.; Vincent, A.L.; Loving, C.L.; Henningson, J.N.; Lager, K.M.; Janke, B.H.; Kehrli, M.E.; Roth, J.A. Kinetics of Lung Lesion Development and Pro-Inflammatory Cytokine Response in Pigs with Vaccine-Associated Enhanced Respiratory Disease Induced by Challenge with Pandemic (2009) A/H1N1 Influenza Virus. Veter. Pathol. 2012, 49, 900–912. [Google Scholar] [CrossRef] [PubMed]
- Belser, J.A.; Johnson, A.; Pulit-Penaloza, J.A.; Pappas, C.; Pearce, M.B.; Tzeng, W.-P.; Hossain, M.J.; Ridenour, C.; Wang, L.; Chen, L.-M.; et al. Pathogenicity testing of influenza candidate vaccine viruses in the ferret model. Virology 2017, 511, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Chiba, S.; Hatta, M.; Pattinson, D.; Yasuhara, A.; Neumann, G.; Kawaoka, Y. Ferret model to mimic the sequential exposure of humans to historical H3N2 influenza viruses. Vaccine 2023, 41, 590–597. [Google Scholar] [CrossRef]
- Bodewes, R.; Kreijtz, J.H.C.M.; Geelhoed-Mieras, M.M.; van Amerongen, G.; Verburgh, R.J.; van Trierum, S.E.; Kuiken, T.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Rimmelzwaan, G.F. Vaccination against Seasonal Influenza A/H3N2 Virus Reduces the Induction of Heterosubtypic Immunity against Influenza A/H5N1 Virus Infection in Ferrets. J. Virol. 2011, 85, 2695–2702. [Google Scholar] [CrossRef]
- Barman, S.; Krylov, P.S.; Fabrizio, T.; Franks, J.; Turner, J.C.; Seiler, P.; Wang, D.; Rehg, J.E.; Erickson, G.A.; Gramer, M.; et al. Pathogenicity and Transmissibility of North American Triple Reassortant Swine Influenza A Viruses in Ferrets. PLoS Pathog. 2012, 8, e1002791. [Google Scholar] [CrossRef]
- Marriott, A.C.; Dove, B.K.; Whittaker, C.J.; Bruce, C.; Ryan, K.A.; Bean, T.J.; Rayner, E.; Pearson, G.; Taylor, I.; Dowall, S.; et al. Low Dose Influenza Virus Challenge in the Ferret Leads to Increased Virus Shedding and Greater Sensitivity to Oseltamivir. PLoS ONE 2014, 9, e94090. [Google Scholar] [CrossRef]
- Gooch, K.E.; Marriott, A.C.; Ryan, K.A.; Yeates, P.; Slack, G.S.; Brown, P.J.; Fothergill, R.; Whittaker, C.J.; Carroll, M.W. Heterosubtypic cross-protection correlates with cross-reactive interferon-gamma-secreting lymphocytes in the ferret model of influenza. Sci. Rep. 2019, 9, 2617. [Google Scholar] [CrossRef]
- Otte, A.; Marriott, A.C.; Dreier, C.; Dove, B.; Mooren, K.; Klingen, T.R.; Sauter, M.; Thompson, K.-A.; Bennett, A.; Klingel, K.; et al. Evolution of 2009 H1N1 influenza viruses during the pandemic correlates with increased viral pathogenicity and transmissibility in the ferret model. Sci. Rep. 2016, 6, 28583. [Google Scholar] [CrossRef]
- Shinya, K.; Makino, A.; Tanaka, H.; Hatta, M.; Watanabe, T.; Le, M.Q.; Imai, H.; Kawaoka, Y. Systemic Dissemination of H5N1 Influenza A Viruses in Ferrets and Hamsters after Direct Intragastric Inoculation. J. Virol. 2011, 85, 4673–4678. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, T.; Watanabe, K.; Sakurai, Y.; Nishi, K.; Yoshikawa, R.; Yasuda, J. Co-infection of SARS-CoV-2 and influenza virus causes more severe and prolonged pneumonia in hamsters. Sci. Rep. 2021, 11, 21259. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Kang, J.; Lyoo, K.; Le, T.B.; Yeo, Y.H.; Wong, S.; Na, W.; Song, D.; Webby, R.J.; Zanin, M.; et al. Severe acute respiratory syndrome coronavirus 2 and influenza A virus co-infection alters viral tropism and haematological composition in Syrian hamsters. Transbound. Emerg. Dis. 2022, 69, e3297–e3304. [Google Scholar] [CrossRef] [PubMed]
- Frere, J.J.; Serafini, R.A.; Pryce, K.D.; Zazhytska, M.; Oishi, K.; Golynker, I.; Panis, M.; Zimering, J.; Horiuchi, S.; Hoagland, D.A.; et al. SARS-CoV-2 infection in hamsters and humans results in lasting and unique systemic perturbations after recovery. Sci. Transl. Med. 2022, 14, eabq3059. [Google Scholar] [CrossRef]
- Roberts, K.L.; Shelton, H.; Stilwell, P.; Barclay, W.S. Transmission of a 2009 H1N1 Pandemic Influenza Virus Occurs before Fever Is Detected, in the Ferret Model. PLoS ONE 2012, 7, e43303. [Google Scholar] [CrossRef] [PubMed]
- Chrun, T.; Maze, E.A.; Vatzia, E.; Martini, V.; Paudyal, B.; Edmans, M.D.; McNee, A.; Manjegowda, T.; Salguero, F.J.; Wanasen, N.; et al. Simultaneous Infection with Porcine Reproductive and Respiratory Syndrome and Influenza Viruses Abrogates Clinical Protection Induced by Live Attenuated Porcine Reproductive and Respiratory Syndrome Vaccination. Front. Immunol. 2021, 12, 758368. [Google Scholar] [CrossRef]
- Holzer, B.; Morgan, S.B.; Martini, V.; Sharma, R.; Clark, B.; Chiu, C.; Salguero, F.J.; Tchilian, E. Immunogenicity and Protective Efficacy of Seasonal Human Live Attenuated Cold-Adapted Influenza Virus Vaccine in Pigs. Front. Immunol. 2019, 10, 2625. [Google Scholar] [CrossRef]
- Marriott, A.C.; Gooch, K.E.; Brown, P.J.; Ryan, K.A.; Jones, N.J.; Merredew, N.; Wiblin, N.; Dibben, O.; Bright, H.; Hallis, B.; et al. Severity of heterosubtypic influenza virus infection in ferrets is reduced by live attenuated influenza vaccine. npj Vaccines 2021, 6, 43. [Google Scholar] [CrossRef]
- Ryan, K.A.; Bewley, K.R.; Watson, R.J.; Burton, C.; Carnell, O.; Cavell, B.E.; Challis, A.; Coombes, N.S.; Davies, E.R.; Edun-Huges, J.; et al. Syrian hamster convalescence from prototype SARS-CoV-2 confers measurable protection against the attenuated disease caused by the Omicron variant. PLOS Pathog. 2023, 19, e1011293. [Google Scholar] [CrossRef]
- Yuan, L.; Zhu, H.; Zhou, M.; Ma, J.; Chen, R.; Chen, Y.; Chen, L.; Wu, K.; Cai, M.; Hong, J.; et al. Gender associates with both susceptibility to infection and pathogenesis of SARS-CoV-2 in Syrian hamster. Signal Transduct. Target. Ther. 2021, 6, 136. [Google Scholar] [CrossRef]
- Dhakal, S.; Ruiz-Bedoya, C.A.; Zhou, R.; Creisher, P.S.; Villano, J.S.; Littlefield, K.; Castillo, J.R.; Marinho, P.; Jedlicka, A.E.; Ordonez, A.A.; et al. Sex Differences in Lung Imaging and SARS-CoV-2 Antibody Responses in a COVID-19 Golden Syrian Hamster Model. mBio 2021, 12, e0097421. [Google Scholar] [CrossRef]
- Newby, C.M.; Rowe, R.; Pekosz, A. Influenza A virus infection of primary differentiated airway epithelial cell cultures derived from Syrian golden hamsters. Virology 2006, 354, 80–90. [Google Scholar] [CrossRef] [PubMed]
- van Riel, D.; Munster, V.J.; de Wit, E.; Rimmelzwaan, G.F.; Fouchier, R.A.M.; Osterhaus, A.D.; Kuiken, T. H5N1 Virus Attachment to Lower Respiratory Tract. Science 2006, 312, 399. [Google Scholar] [CrossRef]
- Shinya, K.; Ebina, M.; Yamada, S.; Ono, M.; Kasai, N.; Kawaoka, Y. Influenza virus receptors in the human airway. Nature 2006, 440, 435–436. [Google Scholar] [CrossRef] [PubMed]
- Shiwa-Sudo, N.; Sakai, Y.; Iwata-Yoshikawa, N.; Watanabe, S.; Yamada, S.; Kuroda, Y.; Yamamoto, T.; Shirakura, M.; Fujisaki, S.; Miyazaki, K.; et al. Impact of Reinfection with SARS-CoV-2 Omicron Variants in Previously Infected Hamsters. J. Virol. 2023, 97, e0136622. [Google Scholar] [CrossRef]
- Mohandas, S.; Yadav, P.D.; Shete, A.; Nyayanit, D.; Sapkal, G.; Lole, K.; Gupta, N. SARS-CoV-2 Delta Variant Pathogenesis and Host Response in Syrian Hamsters. Viruses 2021, 13, 1773. [Google Scholar] [CrossRef] [PubMed]
- Yinda, C.K.; Port, J.R.; Bushmaker, T.; Fischer, R.J.; Schulz, J.E.; Holbrook, M.G.; Shaia, C.; de Wit, E.; van Doremalen, N.; Munster, V.J. Prior aerosol infection with lineage A SARS-CoV-2 variant protects hamsters from disease, but not reinfection with B.1.351 SARS-CoV-2 variant. Emerg. Microbes Infect. 2021, 10, 1284–1292. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.J.; van Doremalen, N.; Adney, D.R.; Yinda, C.K.; Port, J.R.; Holbrook, M.G.; Schulz, J.E.; Williamson, B.N.; Thomas, T.; Barbian, K.; et al. ChAdOx1 nCoV-19 (AZD1222) protects Syrian hamsters against SARS-CoV-2 B.1.351 and B.1.1.7. Nat. Commun. 2021, 12, 5868. [Google Scholar] [CrossRef]
- Tostanoski, L.H.; Yu, J.; Mercado, N.B.; McMahan, K.; Jacob-Dolan, C.; Martinot, A.J.; Piedra-Mora, C.; Anioke, T.; Chang, A.; Giffin, V.M.; et al. Immunity elicited by natural infection or Ad26.COV2.S vaccination protects hamsters against SARS-CoV-2 variants of concern. Sci. Transl. Med. 2021, 13, eabj3789. [Google Scholar] [CrossRef] [PubMed]
- Huo, J.; Mikolajek, H.; Le Bas, A.; Clark, J.J.; Sharma, P.; Kipar, A.; Dormon, J.; Norman, C.; Weckener, M.; Clare, D.K.; et al. A potent SARS-CoV-2 neutralising nanobody shows therapeutic efficacy in the Syrian golden hamster model of COVID-19. Nat. Commun. 2021, 12, 5469. [Google Scholar] [CrossRef]
- Halfmann, P.J.; Iida, S.; Iwatsuki-Horimoto, K.; Maemura, T.; Kiso, M.; Scheaffer, S.M.; Darling, T.L.; Joshi, A.; Loeber, S.; Singh, G.; et al. SARS-CoV-2 Omicron virus causes attenuated disease in mice and hamsters. Nature 2022, 603, 687–692. [Google Scholar] [CrossRef]
- Svyatchenko, V.A.; Ternovoi, V.A.; Lutkovskiy, R.Y.; Protopopova, E.V.; Gudymo, A.S.; Danilchenko, N.V.; Susloparov, I.M.; Kolosova, N.P.; Ryzhikov, A.B.; Taranov, O.S.; et al. Human Adenovirus and Influenza A Virus Exacerbate SARS-CoV-2 Infection in Animal Models. Microorganisms 2023, 11, 180. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ferret (Mustela putorius furo) | Golden Syrian hamster (Mesocricetus auratus) | ||
|---|---|---|---|
| Age | 15–19 weeks | 5 weeks (‘young’) | >20 weeks (‘older’) |
| Infection Dose | 1 × 106 PFU in 200 μL (‘high’) and 1 × 102 PFU in 200 μL (‘low’) | 1 × 106 PFU in 200 μL | |
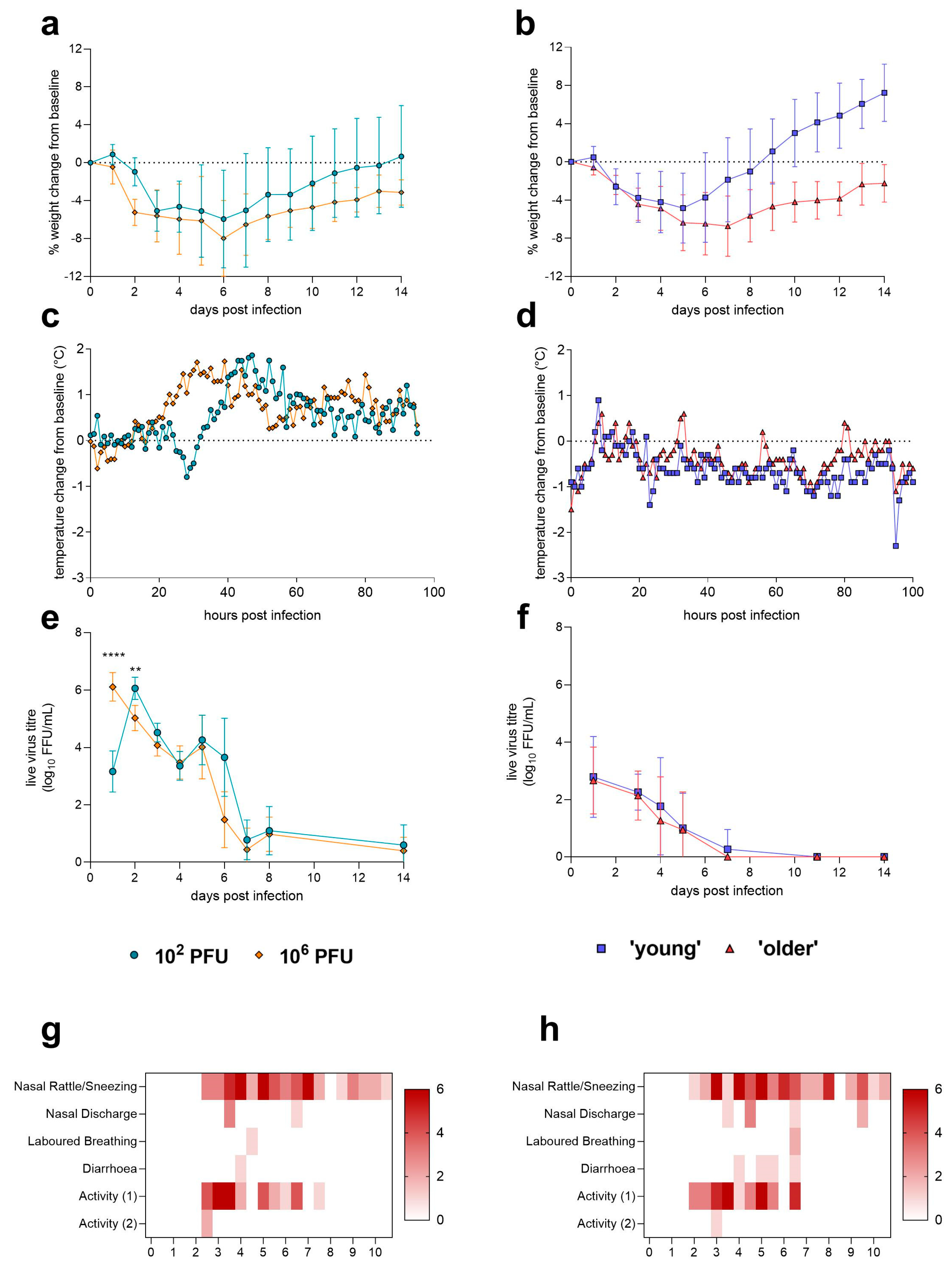
| Weight loss | Yes, average peak – 6 dpi | Yes, average peak – 5 dpi (young) or 7 dpi (older) | |
| Clinical signs | Nasal rattles/sneezing, nasal discharge, laboured breathing, diarrhoea, reduced activity. No differences observed between doses. | None | |
| Temperature | Fever peak at ~35–45 hpi, depending on dose. | Temperature drop observed following infection as well as disruption to diurnal rhythm. Differences observed between male and female hamsters—male hamster temperatures fell significantly following infection. | |
| Shedding of live virus from the URT | Yes, peak observed at 1–2 dpi depending on dose, shedding was detectable until 8 dpi. | Yes, peak observed mainly at 1 dpi; shedding detected until 5 dpi. Differences observed between male and female hamsters at 1 dpi—male hamsters shedding significantly more live virus. | |
| Viral load in LRT | Live virus detected in lung at 4 dpi. Virus only detected in one (high dose) animal at 14 dpi. | Live virus detected in lung of 2/12 (male) at 4 dpi. Assay failure suspected. Method requires further optimisation. | |
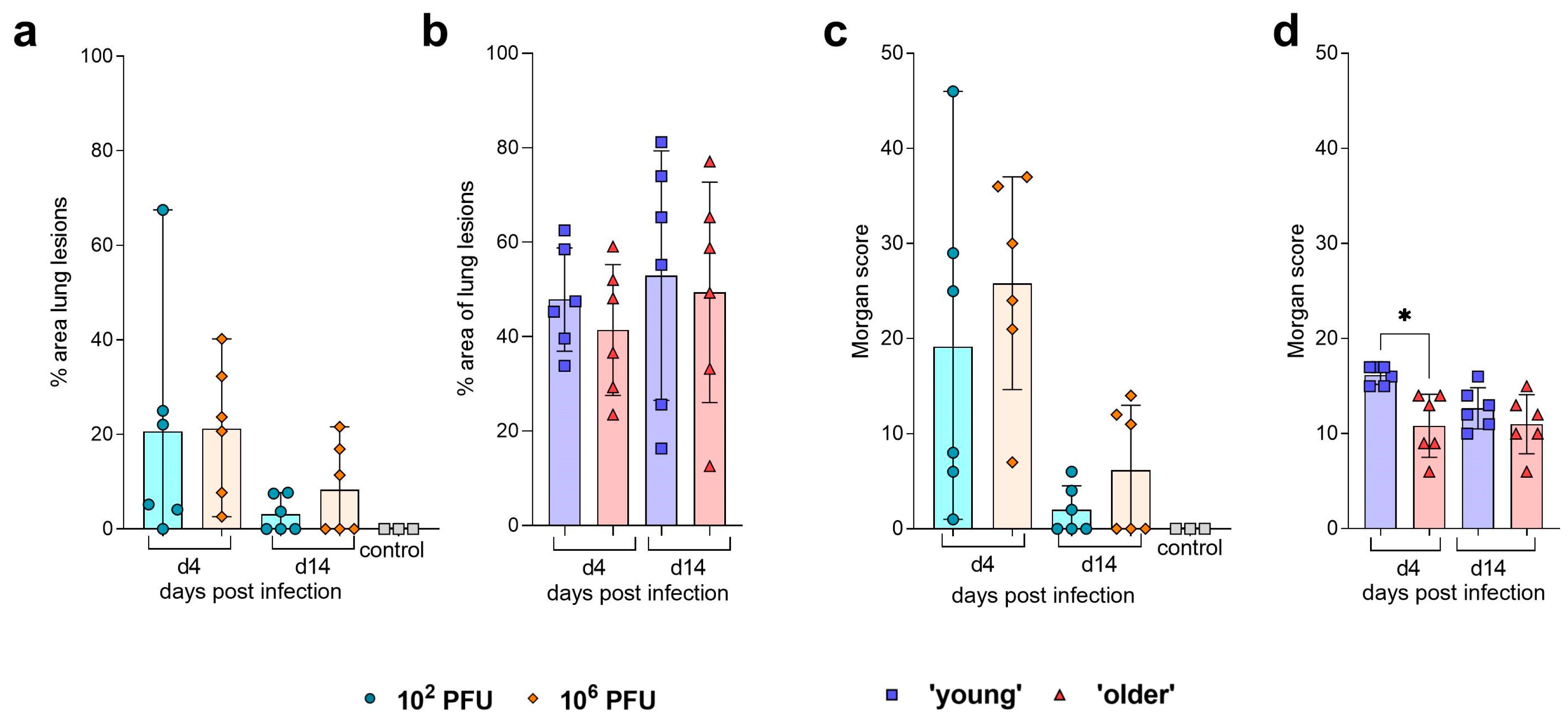
| Pathology | No differences observed in % lesions in the lung. No significant differences observed in assigned ‘Morgan score’ [9] at 4 or 14 dpi. Viral NP staining was detectable at 4 dpi in the NT and lung. | No differences observed in % lesions in the lung. ‘Young’ hamsters were assigned a significantly higher ‘Morgan score’ [9] at 4 dpi. Viral NP staining was detectable at 4 dpi in the NT and lung. | |
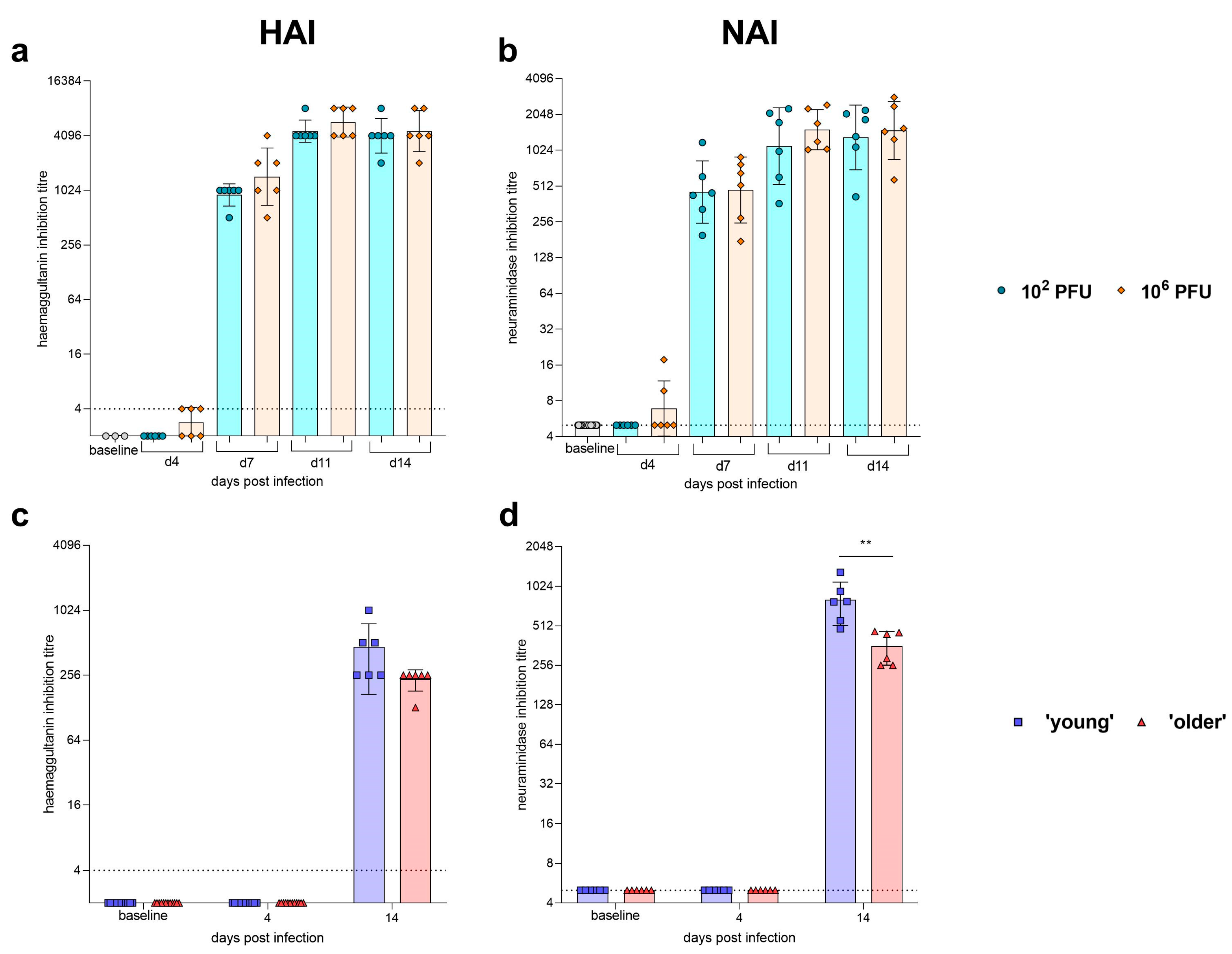
| Humoral Response | Seroconversion detected by HAI and NAI at 7 dpi. No difference between dose observed in HAI and NAI titres. | Seroconversion detected by HAI and NAI at 14 dpi. No differences between ages observed in HAI titres. Young hamsters had significantly higher NAI titres at 14 dpi. | |
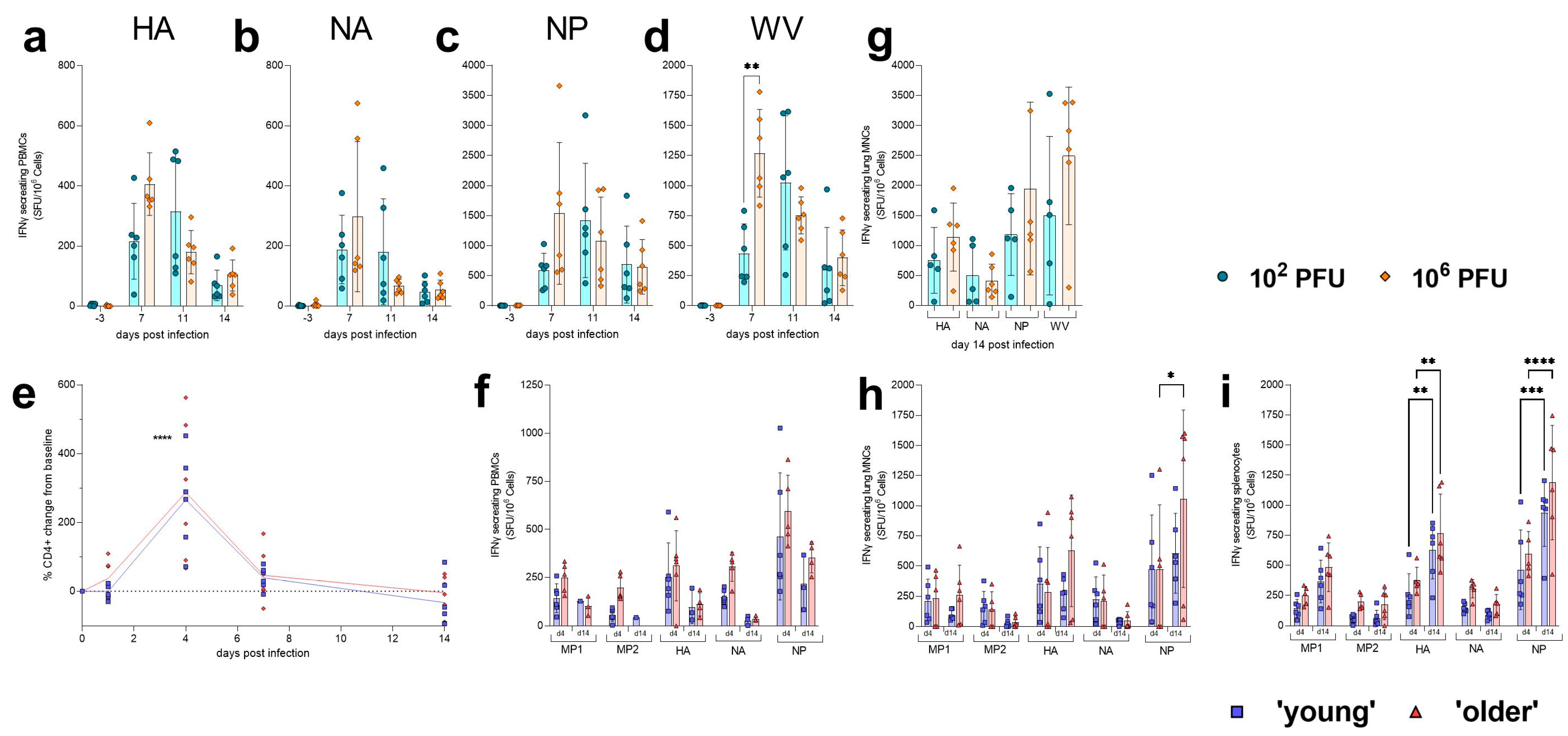
| Cellular Response | Peak of response in PBMCs observed at 7 dpi (high dose) or 11 dpi (low dose). Higher influenza-specific IFNγ responses observed in the lung of high-dose ferrets when compared to low-dose ferrets. NP-specific IFNγ responses were highest in both compartments measured. | No differences observed between ‘young’ and ‘older’ hamsters in amount of CD4+ cells in peripheral blood following infection. No differences in influenza-specific IFNγ responses observed between ‘young’ or ‘older’ hamsters. In general, higher responses were observed at 14 dpi when compared to 4 dpi. NP-specific IFNγ responses were highest in all compartments measured. | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paterson, J.; Ryan, K.A.; Morley, D.; Jones, N.J.; Yeates, P.; Hall, Y.; Whittaker, C.J.; Salguero, F.J.; Marriott, A.C. Infection with Seasonal H1N1 Influenza Results in Comparable Disease Kinetics and Host Immune Responses in Ferrets and Golden Syrian Hamsters. Pathogens 2023, 12, 668. https://doi.org/10.3390/pathogens12050668
Paterson J, Ryan KA, Morley D, Jones NJ, Yeates P, Hall Y, Whittaker CJ, Salguero FJ, Marriott AC. Infection with Seasonal H1N1 Influenza Results in Comparable Disease Kinetics and Host Immune Responses in Ferrets and Golden Syrian Hamsters. Pathogens. 2023; 12(5):668. https://doi.org/10.3390/pathogens12050668
Chicago/Turabian StylePaterson, Jemma, Kathryn A. Ryan, Daniel Morley, Nicola J. Jones, Paul Yeates, Yper Hall, Catherine J. Whittaker, Francisco J. Salguero, and Anthony C. Marriott. 2023. "Infection with Seasonal H1N1 Influenza Results in Comparable Disease Kinetics and Host Immune Responses in Ferrets and Golden Syrian Hamsters" Pathogens 12, no. 5: 668. https://doi.org/10.3390/pathogens12050668