

The Involvement of Neutrophil in the Immune Dysfunction Associated with BVDV Infection

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal
2.2. Virus
2.3. Isolation of Monocytes from Peripheral Blood Mononuclear Cells (PBMCs)
2.4. Development of Monocytes-Derived Macrophages (MDM)
2.5. BVDV Infection of MDM and Supernatant Collection & Irradiation
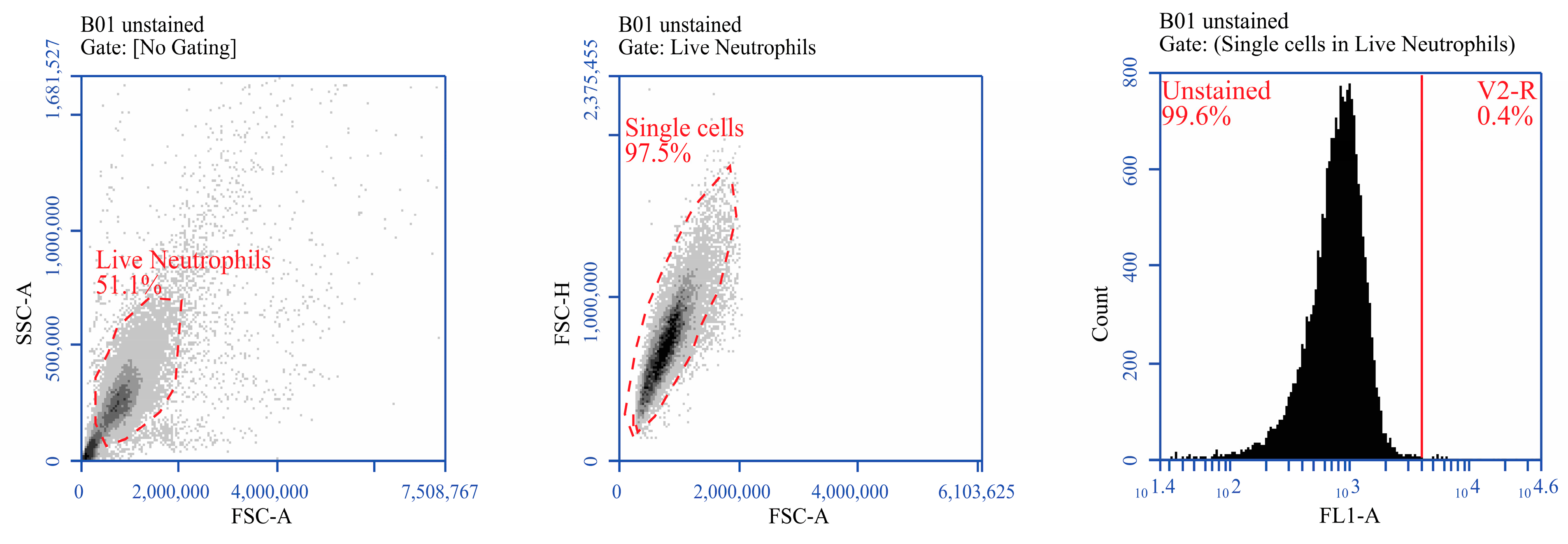
2.6. Neutrophil Isolation
2.7. Neutrophil Treatment with Infected MDM Supernatant and Viability Testing
2.8. Immunostaining and Flow Cytometry of Neutrophil Surface Marker in Response to Infected MDM Supernatant
2.9. Phagocytosis Activity of Neutrophils in Response to Infected MDM Supernatant
2.10. Neutrophil Oxidative Burst Activity in Response to Infected MDM Supernatant
2.11. Nitric Oxide Activity of Neutrophils in Response to Infected MDM Supernatant
2.12. Neutrophil Extracellular Traps (NETs) Formation in Response to Infected MDM Supernatant
2.13. Statistical Analysis
3. Results
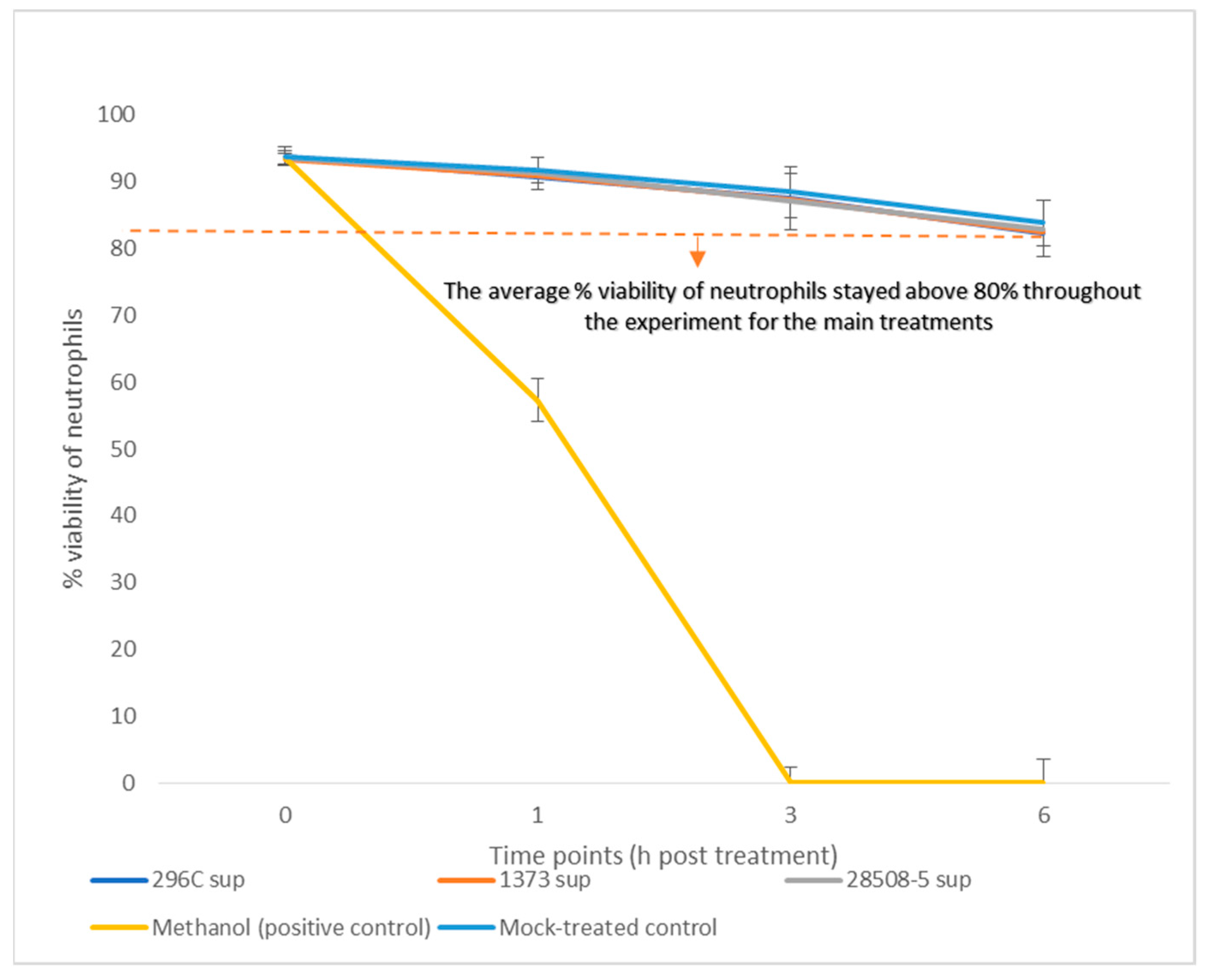
3.1. Effect of BVDV-Infected MDM Supernatant on Neutrophil Viability
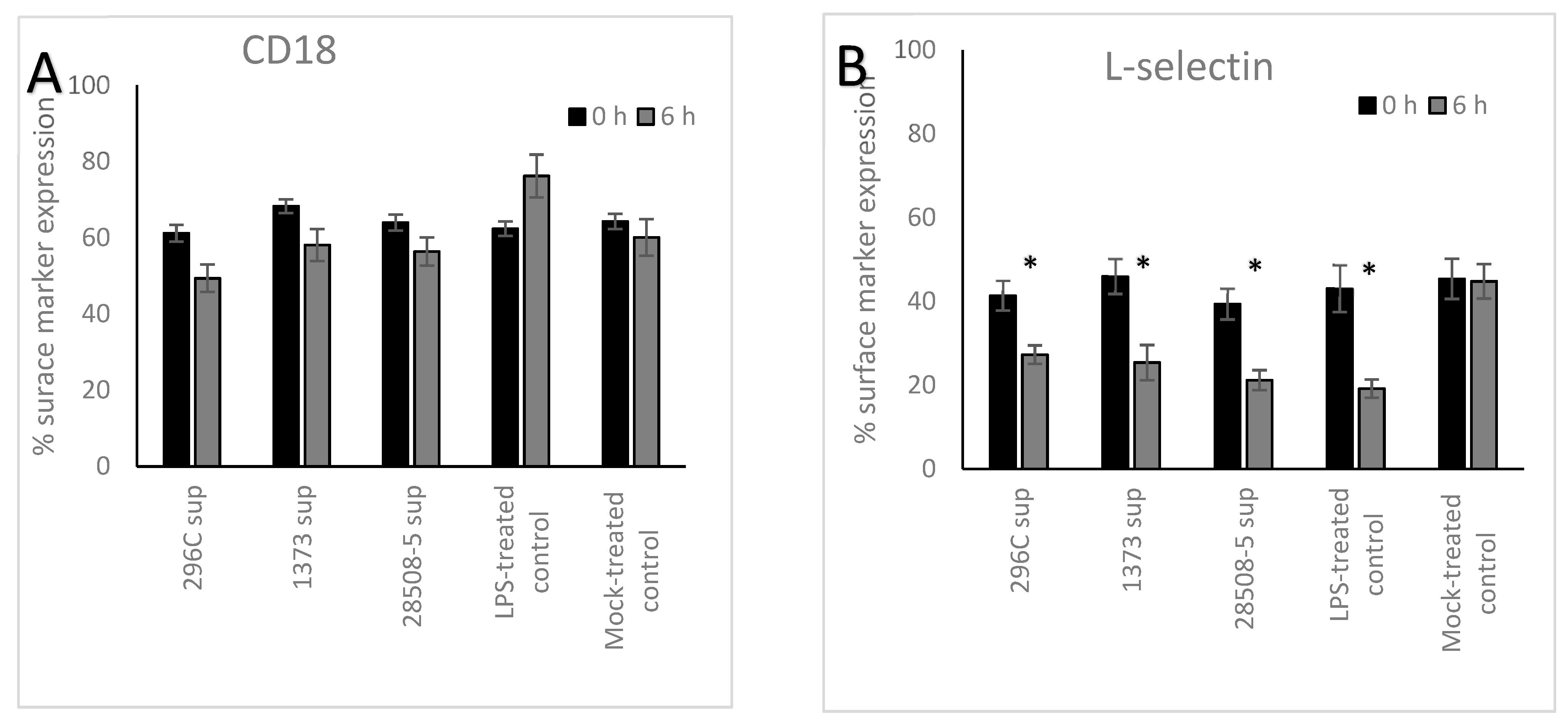
3.2. Effect of BVDV-Infected MDM Supernatant on Neutrophil’s Surface Marker Expression
3.3. Effect of BVDV-Infected MDM Supernatant on Neutrophil Phagocytic Activity
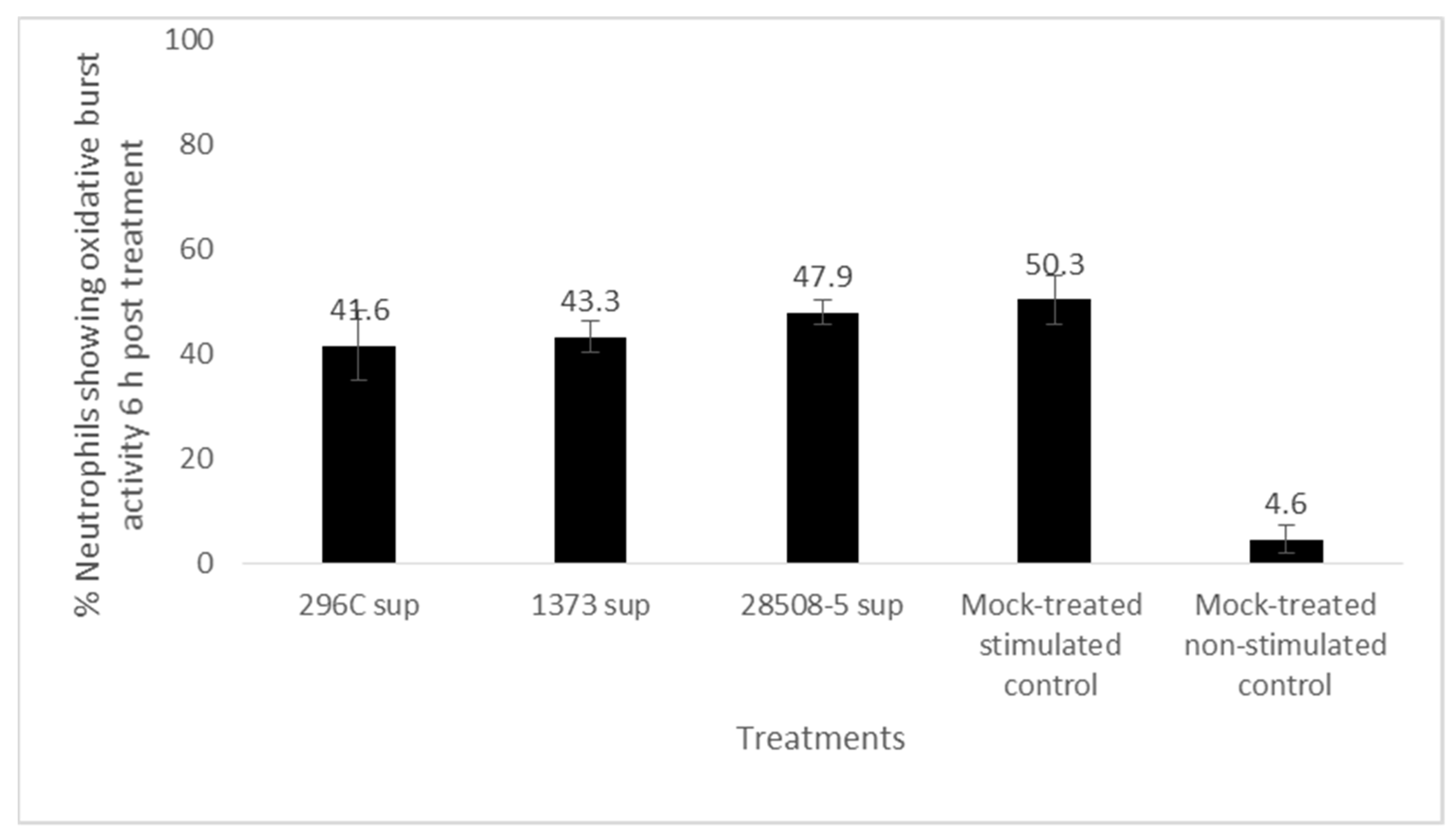
3.4. Effect of BVDV-Infected MDM Supernatant on Neutrophil Oxidative Burst
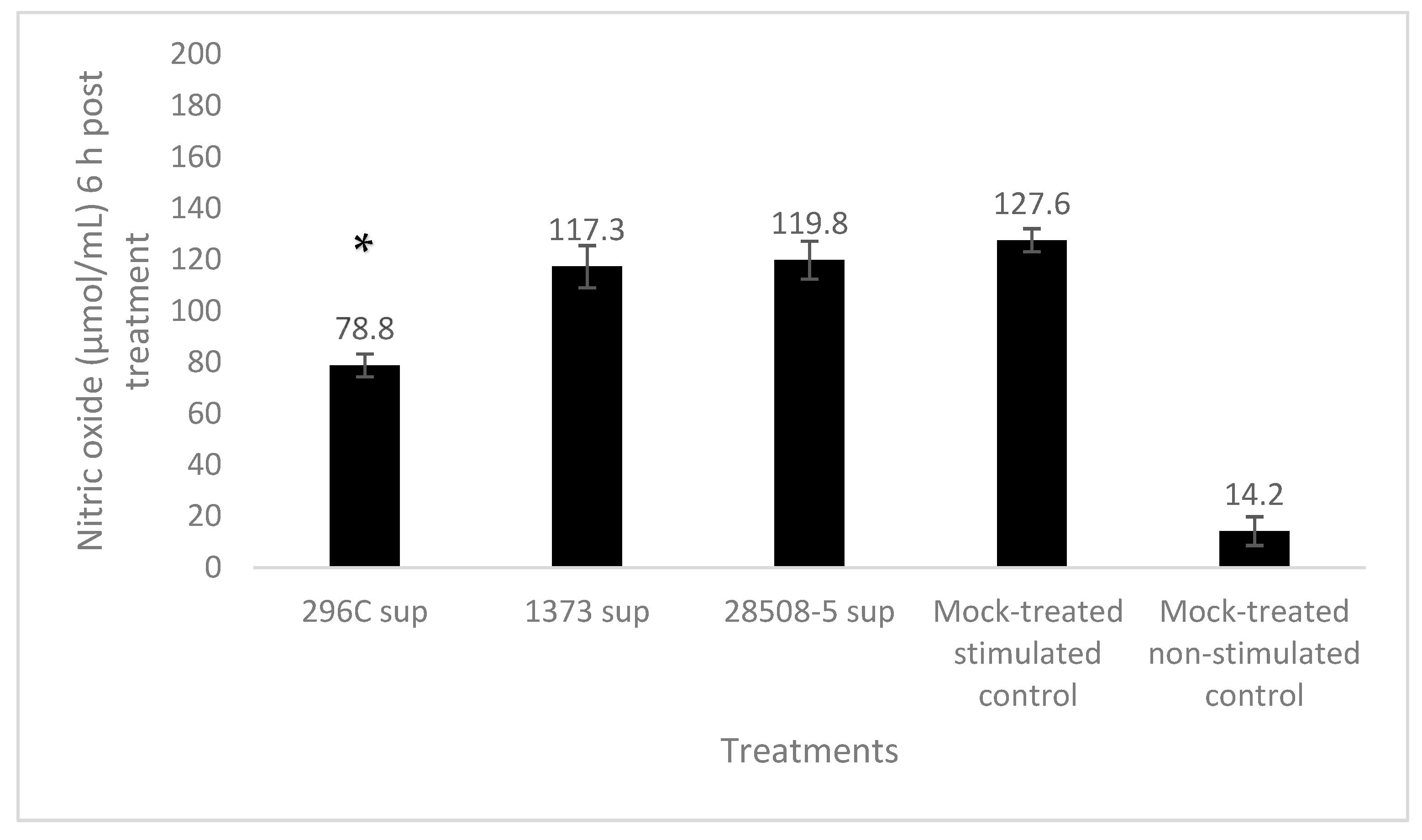
3.5. Effect of BVDV-Infected MDM Supernatant on Neutrophil Nitric Oxide (NO) Activity
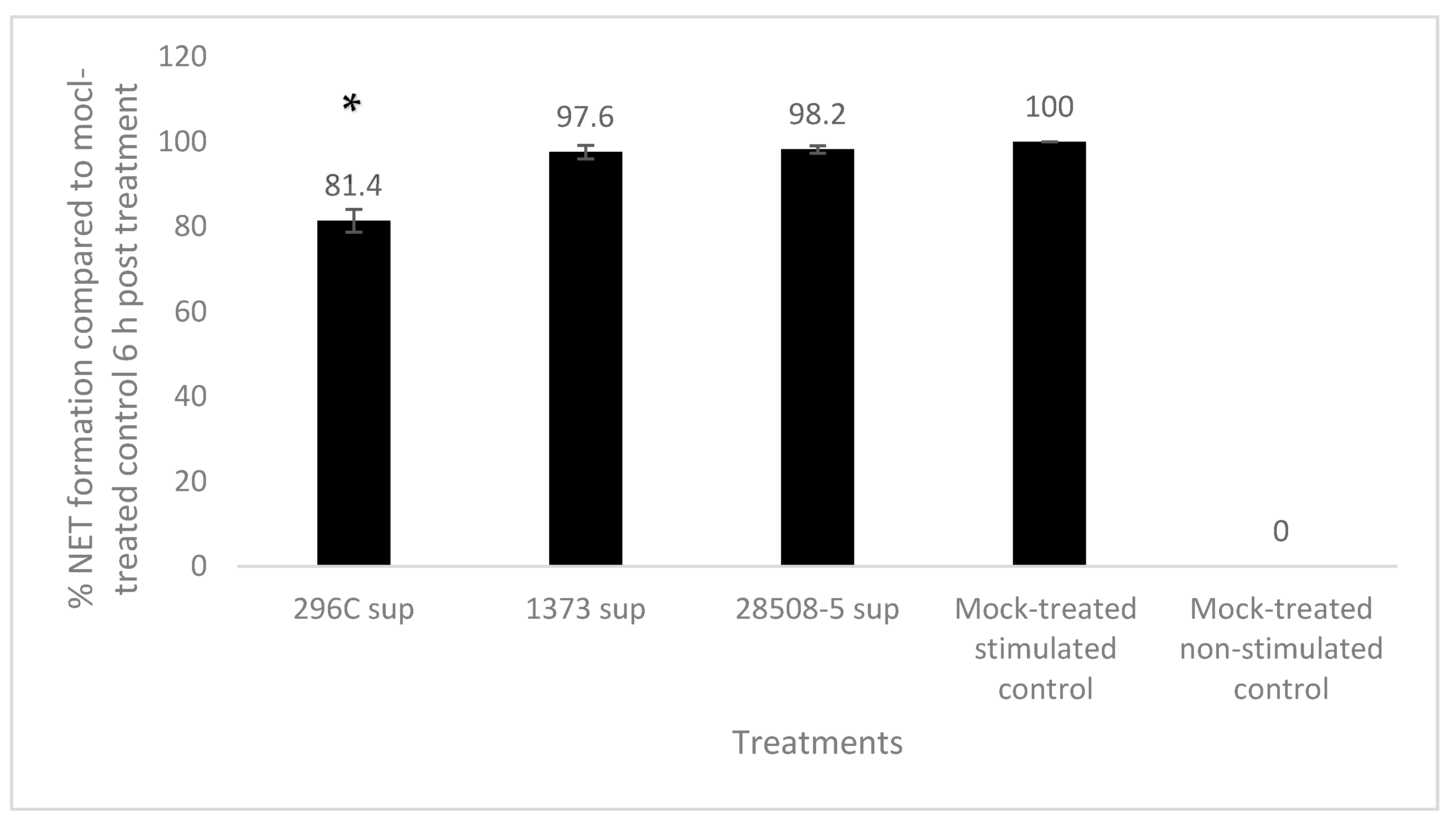
3.6. Effect of BVDV-Infected MDM Supernatant on Neutrophil Extracellular Trap (NET) Formation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Houe, H. Economic impact of BVDV infection in dairies. Biologicals 2003, 31, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Fourichon, C.; Beaudeau, F.; Bareille, N.; Seegers, H. Quantification of economic losses consecutive to infection of a dairy herd with bovine viral diarrhoea virus. Prev. Vet. Med. 2005, 72, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Ridpath, J. Preventive strategy for BVDV infection in North America. Jpn. J. Vet. Res. 2012, 60, S41–S49. [Google Scholar] [PubMed]
- Chase, C.C.; Elmowalid, G.; Yousif, A.A. The immune response to bovine viral diarrhea virus: A constantly changing picture. Vet. Clin. Food Anim. Pract. 2004, 20, 95–114. [Google Scholar] [CrossRef]
- Kobayashi, S.D.; DeLeo, F.R. Role of neutrophils in innate immunity: A systems biology-level approach. Wiley Interdiscip. Rev. Syst. Biol. Med. 2009, 1, 309–333. [Google Scholar] [CrossRef]
- Jaillon, S.; Galdiero, M.R.; Del Prete, D.; Cassatella, M.A.; Garlanda, C.; Mantovani, A. Neutrophils in innate and adaptive immunity. In Seminars Immunopathol; Springer: Berlin/Heidelberg, Germany, 2013; Volume 35, pp. 377–394. [Google Scholar]
- McAvoy, E.F.; McDonald, B.; Parsons, S.A.; Wong, C.H.; Landmann, R.; Kubes, P. The role of CD14 in neutrophil recruitment within the liver microcirculation during endotoxemia. J. Immunol. 2011, 186, 2592–2601. [Google Scholar] [CrossRef]
- Gao, J.X.; Issekutz, A.C. Mac-1 (CD11b/CD18) is the predominant β2 (CD18) integrin mediating human neutrophil migration through synovial and dermal fibroblast barriers. Immunology 1996, 88, 463–470. [Google Scholar] [CrossRef]
- Gomez, J.C.; Doerschuk, C.M. The role of CD18 in the production and release of neutrophils from the bone marrow. Lab. Investig. 2010, 90, 599–610. [Google Scholar] [CrossRef]
- Von Andrian, U.H.; Berger, E.M.; Ramezani, L.; Chambers, J.D.; Ochs, H.D.; Harlan, J.M.; Paulson, J.C.; Etzioni, A.; Arfors, K.E. In vivo behavior of neutrophils from two patients with distinct inherited leukocyte adhesion deficiency syndromes. J. Clin. Investig. 1993, 91, 2893–2897. [Google Scholar] [CrossRef]
- Thakur, N.; Evans, H.; Abdelsalam, K.; Farr, A.; Rajput, M.K.; Young, A.J.; Chase, C.C. Bovine viral diarrhea virus compromises neutrophil’s functions in strain dependent manner. Microb. Pathog. 2020, 149, 104515. [Google Scholar] [CrossRef]
- Rajput, M.K.; Darweesh, M.F.; Park, K.; Braun, L.J.; Mwangi, W.; Young, A.J.; Chase, C.C. The effect of bovine viral diarrhea virus (BVDV) strains on bovine monocyte-derived dendritic cells (Mo-DC) phenotype and capacity to produce BVDV. Virol. J. 2014, 11, 44. [Google Scholar] [CrossRef] [PubMed]
- Abdelsalam, K.; Rajput, M.; Elmowalid, G.; Sobraske, J.; Thakur, N.; Abdallah, H.; Ali, A.A.; Chase, C.C. The effect of bovine viral diarrhea virus (BVDV) strains and the corresponding infected-macrophages’ supernatant on macrophage inflammatory function and lymphocyte apoptosis. Viruses 2020, 12, 701. [Google Scholar] [CrossRef] [PubMed]
- Potgieter, L.N. Immunology of bovine viral diarrhea virus. Vet. Clin. N. Am. Food Anim. Pract. 1995, 11, 501–520. [Google Scholar] [CrossRef] [PubMed]
- Byers, S.R.; Evermann, J.F.; Bradway, D.S.; Parish, S.M.; Barrington, G.M. Evaluation of a commercial bovine viral diarrhea virus vaccine in nonpregnant female alpacas (Vicugna pacos). Vaccine 2010, 28, 591–593. [Google Scholar] [CrossRef]
- Fulton, R.W.; Saliki, J.T.; Burge, L.J.; d’Offay, J.M.; Bolin, S.R.; Maes, R.K.; Baker, J.C.; Frey, M.L. Neutralizing antibodies to type 1 and 2 bovine viral diarrhea viruses: Detection by inhibition of viral cytopathology and infectivity by immunoperoxidase assay. Clin. Diagn. Lab. Immunol. 1997, 4, 380–383. [Google Scholar] [CrossRef]
- Schaut, R.G.; Ridpath, J.F.; Sacco, R.E. Bovine viral diarrhea virus type 2 impairs macrophage responsiveness to toll-like receptor ligation with the exception of toll-like receptor 7. PLoS ONE 2016, 11, e0159491. [Google Scholar] [CrossRef]
- Strober, W. Trypan blue exclusion test of cell viability. Curr. Protoc. Immunol. 1997, 21, A–3B. [Google Scholar]
- Abd, G.A.; Elmowalid, A. Unmasking the Effect of Bovine Viral Diarrhea Virus on Macrophage Inflammatory Functions. Ph.D. Thesis, South Dakota State University, Brookings, South Dakota, 2003. [Google Scholar]
- Jungmann, A.; Nieper, H.; Müller, H. Apoptosis is induced by infectious bursal disease virus replication in productively infected cells as well as in antigen-negative cells in their vicinity. J. Gen. Virol. 2001, 82, 1107–1115. [Google Scholar] [CrossRef]
- Blond, D.; Raoul, H.; Le Grand, R.; Dormont, D. Nitric oxide synthesis enhances human immunodeficiency virus replication in primary human macrophages. J. Virol. 2000, 74, 8904–8912. [Google Scholar] [CrossRef]
- Fulton, R.W. Impact of species and subgenotypes of bovine viral diarrhea virus on control by vaccination. Anim. Health Res. Rev. 2015, 16, 40–54. [Google Scholar] [CrossRef]
- Roth, J.A.; Kaeberle, M.L.; Griffith, R.W. Effects of bovine viral diarrhea virus infection on bovine polymorphonuclear leukocyte function. Am. J. Vet. Res. 1981, 42, 244–250. [Google Scholar] [PubMed]
- Walzog, B.; Scharffetter-Kochanek, K.; Gaehtgens, P. Impairment of neutrophil emigration in CD18-null mice. Am. J. Physiol.-Gastrointest. Liver Physiol. 1999, 276, G1125–G1130. [Google Scholar] [CrossRef] [PubMed]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Al-Kubati, A.A.; Hussen, J.; Kandeel, M.; Al-Mubarak, A.I.; Hemida, M.G. Recent advances on the bovine viral diarrhea virus molecular pathogenesis, immune response, and vaccines development. Front. Vet. Sci. 2021, 8, 665128. [Google Scholar] [CrossRef] [PubMed]
- Jirillo, E.; Greco, B.; Caradonna, L.; Satalino, R.; Pugliese, V.; Cozzolongo, R.; Cuppone, R.; Manghisi, O.G. Evaluation of cellular immune responses and soluble mediators in patients with chronic hepatitis C virus (cHCV) infection. Immunopharmacol. Immunotoxicol. 1995, 17, 347–364. [Google Scholar] [CrossRef]
- Roth, J.A.; Kaeberle, M.L. Suppression of neutrophil and lymphocyte function induced by a vaccinal strain of bovine viral diarrhea virus with and without the administration of ACTH. Am. J. Vet. Res. 1983, 44, 2366–2372. [Google Scholar]
- Lasarte, J.J.; Sarobe, P.; Boya, P.; Casares, N.; Arribillaga, L.; de Cerio, A.L.; Gorraiz, M.; Borrás-Cuesta, F.; Prieto, J. A recombinant adenovirus encoding hepatitis C virus core and E1 proteins protects mice against cytokine-induced liver damage. Hepatology 2003, 37, 461–470. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BVDV Strains | Biotype | Virulence | Titer |
|---|---|---|---|
| 1373 | ncp, BVDV-2 | High virulence | 5.8 × 105 |
| 28508-5 | ncp, BVDV-2 | Low virulence | 7.2 × 106 |
| 296C | cp, BVDV-2 | Virulent | 5.2 × 105 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelsalam, K.; Kaushik, R.S.; Chase, C. The Involvement of Neutrophil in the Immune Dysfunction Associated with BVDV Infection. Pathogens 2023, 12, 737. https://doi.org/10.3390/pathogens12050737
Abdelsalam K, Kaushik RS, Chase C. The Involvement of Neutrophil in the Immune Dysfunction Associated with BVDV Infection. Pathogens. 2023; 12(5):737. https://doi.org/10.3390/pathogens12050737
Chicago/Turabian StyleAbdelsalam, Karim, Radhey S Kaushik, and Christopher Chase. 2023. "The Involvement of Neutrophil in the Immune Dysfunction Associated with BVDV Infection" Pathogens 12, no. 5: 737. https://doi.org/10.3390/pathogens12050737