


Natural and Engineered Resistance Mechanisms in Plants against Phytoviruses

1
Department of Plant Pathology, College of Agriculture, Punjab Agricultural University, Ludhiana 141004, Punjab, India
2
Department of Vegetable Science, College of Horticulture and Forestry, Punjab Agricultural University, Ludhiana 141004, Punjab, India
*
Author to whom correspondence should be addressed.
Pathogens 2023, 12(4), 619; https://doi.org/10.3390/pathogens12040619
Submission received: 9 March 2023
/
Revised: 14 April 2023
/
Accepted: 16 April 2023
/
Published: 19 April 2023
(This article belongs to the Special Issue Emerging and Re-emerging Plant Viruses in a Context of Global Change)
Abstract
:Plant viruses, as obligate intracellular parasites, rely exclusively on host machinery to complete their life cycle. Whether a virus is pathogenic or not depends on the balance between the mechanisms used by both plants and viruses during the intense encounter. Antiviral defence mechanisms in plants can be of two types, i.e., natural resistance and engineered resistance. Innate immunity, RNA silencing, translational repression, autophagy-mediated degradation, and resistance to virus movement are the possible natural defence mechanisms against viruses in plants, whereas engineered resistance includes pathogen-derived resistance along with gene editing technologies. The incorporation of various resistance genes through breeding programmes, along with gene editing tools such as CRISPR/Cas technologies, holds great promise in developing virus-resistant plants. In this review, different resistance mechanisms against viruses in plants along with reported resistance genes in major vegetable crops are discussed.
1. Introduction
Plant viruses cause significant yield loss in the cultivation of many agricultural and horticultural crops around the world. Conventional methods of virus control have been used for a long time, including crop rotation, other types of cultivation, removal of infected plant debris, and chemical control of insect vectors. Since these methods have very little impact in the mitigation of virus diseases, numerous inventive methods to manage viral infections in plants have been developed as a result of a growing understanding of the molecular genetics of plant viruses as well as of the hosts’ defence mechanisms. The primary defence mechanisms against viruses in plants include innate immunity, RNA silencing, translational repression, dominant resistance genes, and autophagy-mediated degradation [1]. The rise of gene editing technologies has also enabled researchers/scientists to engineer artificial resistance against viruses in plants. How do different antiviral mechanisms in plants operate to limit the successful pathogenesis of viruses? In this review, we summarise the recent advances in the plant defence mechanisms against phytoviruses along with some reported virus resistance genes in major vegetable crops.
2. Virus Infection Cycle within the Plants
The virus infection cycle within the plants can be narrated as described by Calil and Fontes [2]. Viruses enter plants through wounds created mechanically, vectors, pollen, seeds or vegetative propagation, pinocytosis, fungal parasites, and epidermal hairs. After entry, the uncoating of the virus capsid takes place in the cytoplasm. At this stage, the translation of viral proteins occurs using host translation machinery. Translation events are further divided into two parts, i.e., early translation (synthesis of proteins required for viral replication, e.g., RNA-dependent RNA polymerase) and late translation (synthesis of coat proteins). Using these replication-associated proteins, virus genome replication occurs either in the cytoplasm or in the nucleus using different mechanisms. The progeny genomic strands are then encapsidated with newly synthesised coat proteins, leading to numerous progeny virion particles. These progeny virion particles move from one cell to another via plasmodesmata and/or long-distance movement occurs through phloem with the help of virus movement proteins (MP). Finally, they get released outside the host cell and transmitted to new hosts by means of different mechanisms, and this cycle of infection continues.
This infection process is targeted by several host defence systems which help the host to counter the virus attack. There are several points in the infection cycle which can be targeted by host defences such as the uncoating of the virus capsid, translation events, encapsidation, and cell-to-cell movement (Figure 1).
3. Antiviral Defence Mechanisms in Plants
This can be divided into two types: Natural resistance to plant viruses and Engineered resistance to plant viruses.
3.1. Natural Resistance to Plant Viruses
Possible natural resistance mechanisms against viruses include innate antiviral immunity, translation repression, small RNA-mediated antiviral defence, dominant viral resistance genes, resistance to virus movement, autophagy, and cross protection.
3.1.1. Innate Antiviral Immunity
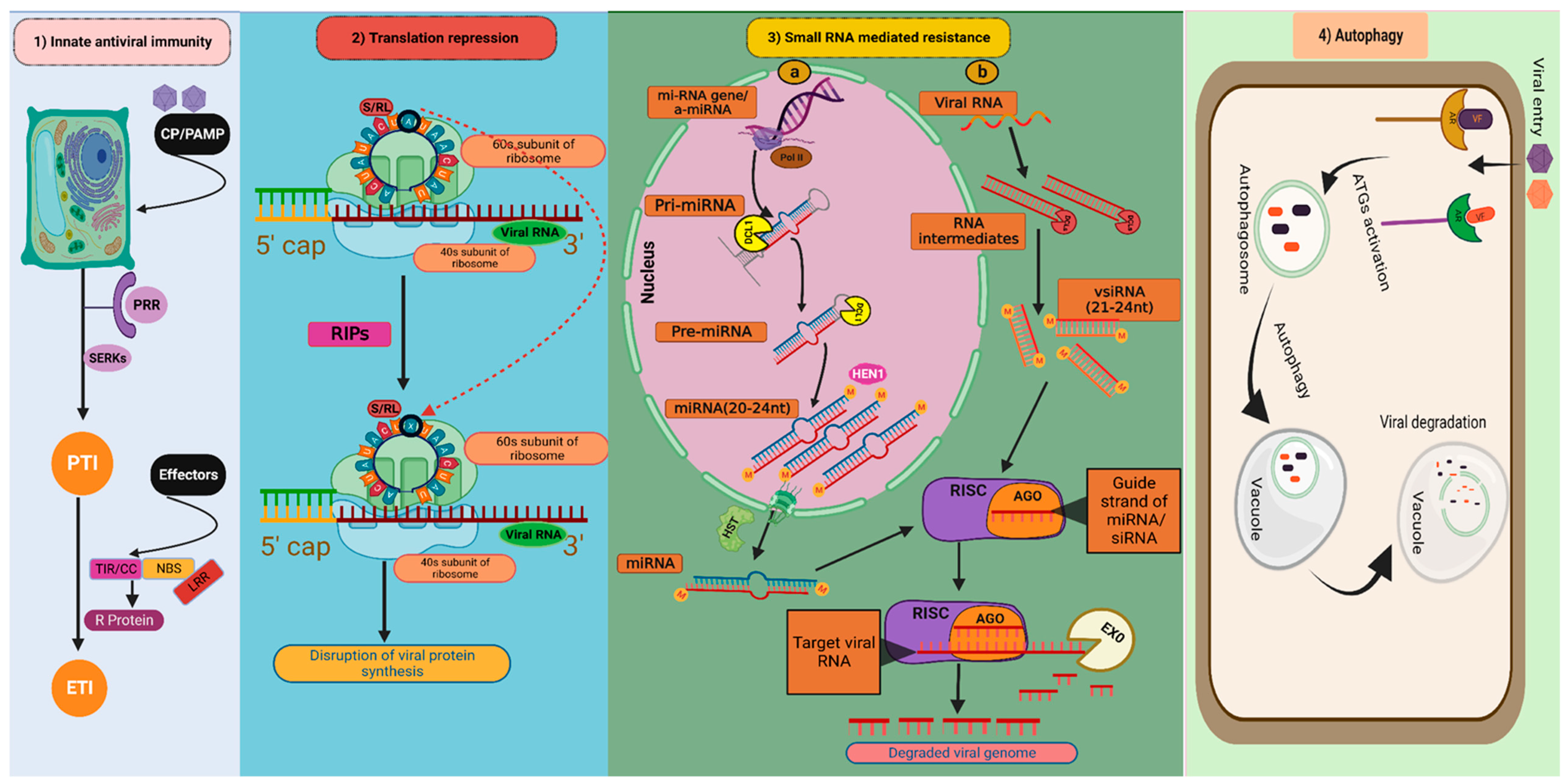
There are two layers of plant immune responses against microbial pathogens, i.e., PAMP (pathogen-associated molecular pattern) triggered immunity (PTI) and Effector-triggered immunity (ETI) (Figure 2 (1)). PTI is the initial mechanism by which plants detect the microbes at the cell membrane through detection of conserved PAMPs by extracellular pattern recognition receptors (PRRs) [3]. PRRs begin to dimerize as soon as the PAMPs are detected and associate with cofactors such as somatic embryogenesis receptor-like kinases (SERKs). This sets off a cascade of intracellular signalling events, such as the generation of reactive oxygen species (ROS), ion influx, increase in the production of defence hormones, and mitogen-activated protein kinases (MAPKs) activation. All these events result in the expression of the pathogenesis-related (PR) proteins, synthesis and deposition of callose at the plasmodesmata, and cell wall strengthening, leading to the generation of resistance response. [4]. Activation of PTI sometimes leads to hypersensitive response (HR) causing programmed cell death (PCD) that causes necrotic spots at the infection site [5]. Although plant PTI against other phytopathogens is well understood, plant viruses are traditionally known as non-PAMP coding pathogens [6]. However, some evidence implies that PTI also has a significant influence on both susceptible and resistant plant-virus interactions. For instance, exogenous application of double-strand RNA (dsRNA) caused SERK-1-dependent PTI responses in Arabidopsis [7]. In another instance, tobacco and Arabidopsis showed PTI-like responses due to the coat proteins of tobacco mosaic virus and potato virus X, respectively [8,9].
Pathogens in response to PTI introduce particular proteins called effectors in plant cells to weaken PTI-mediated defence. In response to these effectors, host plants depend on specific intracellular receptors known as R gene proteins which cause direct or indirect identification of the pathogen effector molecules. These R gene products can block the effectors and activate the effector-triggered immunity (ETI) [10]. R genes mediating resistance against various plant viruses have been extensively cloned over the last decade due to their apparent practical value. Functional R proteins are composed of three domains: a central nucleotide-binding site (NBS) domain, a leucine-rich repeat (LRR) domain, and an N terminal Toll Interleukin-1 receptor (TIR) or coiled-coil (CC) domain. Systemic acquired resistance (SAR) is a condition that can occur as a result of both PTI and ETI and is characterised by the development of resistance in the tissues that are distal to the infection [11]. While the non-expressor of PR1 (NPR1) is a protein with ankyrin domains, which is necessary for triggering salicylic acid (SA) signalling and establishing SAR, salicylic acid is the principal plant hormone responsible for establishing SAR [12]. SAR, together with pathogenesis-related proteins, confers resistance to the host plants against various pathogens [13].
3.1.2. Translation Repression as Virus Resistance
The initiation of translation is a critical step in protein synthesis that demands an array of eukaryotic initiation factors (eIFs). The translation of eukaryotic mRNA hinges on the association between the translation eukaryotic initiation factor 4E (eIF4E) and their 5′ m7 G cap structure, and the 3′ polyA tail’s interaction with the polyA-binding protein (PABP) also heightens the process. Although translation factors are not typically encoded by viruses, they have developed several techniques to hijack translation factors from their hosts, resulting in the promotion of viral RNA translation while compromising the translation of endogenous mRNAs. Mutated isoforms of the translation IFs, i.e., eIF4E and eIF4G have been found linked to many plant recessive resistance genes. These mutations typically prevent host factors from interacting with viral RNAs or proteins to suppress viral protein translation, an endogenous antiviral mechanism that has emerged recently [14]. Recessive resistance occurs when a component (or components) of the translation machinery recruited by viruses ceases to operate, resulting in a ‘loss of susceptibility’ to viruses. For instance, a family of proteins known as ribosome-inactivating proteins (RIPs) can inhibit the synthesis of new proteins by depurinating the sarcin/ricin loop (SRL) of rRNA [15] (Figure 2 (2)). The most thoroughly studied RIP with antiviral activity is the pokeweed antiviral protein (PAP) from Phytolacca americana. PAP slows down the spread of various plant viruses, including the cauliflower mosaic virus, potato virus X, and cucumber mosaic virus [16]. In the interaction between plants and viruses, small RNA-related translation suppression can be quite significant. For instance, in tomato plants infected with the tomato ringspot virus, recovery of symptoms is associated with the AGO1-dependent translation repression of viral RNA2. [17]. Additionally, the inability of one strain of plum pox virus to recruit translation initiation factors conferred resistance against this strain in wild-type Arabidopsis thaliana and Chenopodium foetidum [18].
3.1.3. Small RNA-Mediated Antiviral Defence
Small RNAs (sRNA) are essential for the epigenetic and post-transcriptional control of gene expression in plants throughout growth, developmental, and biotic/abiotic stress responses. Recent research has highlighted the role of two distinct families of small RNAs, known as small interfering RNA (siRNA) and micro RNA (miRNA) with respect to biotic stress responses in plants. By regulating the gene expression of the modulators of host defence pathways, these sRNAs activate antiviral defence during virus infection. RNA silencing, commonly referred to as RNA interference (RNAi), is a conserved evolutionary method for regulating endogenous expressions of genes and preventing the entry of foreign nucleic acids such as viruses and transposons [19]. The plant type III endoribonucleases or dicer-like (DCL) proteins detect and cleave the virus-derived double-stranded RNA (dsRNA) into small 20–24 nucleotide RNA duplexes known as virus-derived short interfering small RNAs (vsiRNAs) [20]. The vsiRNAs incorporated into argonaute proteins (AGOs) form the core component of the RNA-induced silencing complex (RISC), which cleaves homologous viral RNAs and/or suppresses translation of viral protein synthesis [21,22] (Figure 2 (3b))
Unlike siRNAs, miRNAs are endogenously produced non-coding short RNAs by RNA polymerase II from ssRNA precursors with a hairpin structure. [23]. Along with the well-established function of vsiRNAs, research findings suggest that miRNAs play a significant role in plant antiviral defence. At the initiation of miRNA synthesis, a primary miRNA transcript, or Pri-miRNA is synthesised after transcription and is made up of an incomplete stem-like structure of 100–120 nucleotides. Then, the precursor miRNA (Pre-miRNA) which is about 70 nucleotides long, formed after processing of Pri-mRNA by the DCL1 complex in the nucleus. Finally, the cleaving of 20–24 nucleotides from the initial cleavage point results in the formation and release of the miRNA/miRNA duplex from the stem-like structure. HUA Enhancer 1 (HEN1) methylates miRNA-miRNA duplex to protect it from degradation in the cytosol. Finally, they are transported to the cytosol by exportin-5 homolog HASTY (HST). In the cytosol, mature miRNAs are incorporated into RISC complexes having AGO proteins. The majority of the animal miRNAs that have been studied so far have miRNA and mRNA base pairing, which hinders the target mRNA translation. This is one of two ways that miRNAs guide RISC to down-regulate target mRNAs. Contrarily, the majority of plant miRNAs form substantial base pairing and directly cleave their target mRNAs. [24] (Figure 2 (3a)). Recently, the roles of various miRNAs, such as miR168, miR528, miR319, and miR444 in rice antiviral immunity have been identified. By modifying jasmonic acid (JA) signalling, MiR319 has been reported to confer antiviral resistance to rice against the rice-ragged stunt virus and to wheat against the rice black-streaked dwarf virus. [25].
3.1.4. Dominant Viral Resistance Genes
A few dominant resistance genes that function independently of the traditional innate immune signalling pathway have also been discovered in the past ten years. These dominant resistance genes express proteins that cannot be incorporated into the so-called plant innate immunity because they differ structurally from conventional R proteins. The majority of these dominant resistance genes inhibit the action of viral proteins by interacting with them in a direct manner. These dominant resistance gene products are referred to as atypical dominant viral resistance proteins (ADVRPs). The majority of ADVRPs that have been identified so far are from a protein family called lectin. Restricted TEV movement (RTM) 1, a member of the lectin-protein family is found in Arabidopsis, and it particularly provides resistance to a number of potyviruses, including TEV, lettuce mosaic virus (LMV), and plum pox virus by inhibiting their long-distance movement [26]. A lectin-like ADVRP of Arabidopsis called Jacaline-type lectin required for potexvirus resistance 1 (JAX1) imparts broad spectrum resistance to potexviruses at the initial infection stage by suppressing the activity of viral RNA-dependent RNA polymerase (RdRp) [27]. BanLec-1, another lectin from Musa paradisiaca, attaches to the TMV CP and inhibits virus infection in plants [28].
3.1.5. Resistance to Virus Movement within and between the Cells
For plant viruses to infect the entire plant system, they must spread from the initially infected cells to nearby ones. This must occur after the virus multiplication has begun within the cytoplasm and/or nucleus of a plant cell in a susceptible host. Host resistance to viral infection is evident when the virus only appears to affect one or a small number of cells but is unable to move past this initial focus of infection. At this point, resistance can be brought on by active host defence systems that swiftly thwart virus spread or by a breakdown in the connections between plant and viral components necessary for cell-to-cell movement. It is known that a variety of host gene alterations hinder the cell-to-cell movement of plant viruses. The cum1 and cum2 mutations in Arabidopsis cause reduced multiplication of CMV, thus rendering its movement in adjacent cells [29]. Pvr11 and Sbm1 were found to be mutated at a locus expressing eIF4E in pepper and pea, respectively [30,31]. eIF4E is thought to play a role in viral RNA replication or translation, although it may also be involved in cell-to-cell movement. The restricted-TEV-movement (RTM) genes RTM1, RTM2, and RTM3 prevent systemic tobacco etch virus (TEV) movement between plant cells. These genes interact with the virus CP and are expressed in phloem sieve elements [32]. Similar to this, BTR1 is a ribonucleoprotein K-homology RNA binding protein that binds to ToMV (tomato mosaic virus) genomic RNA and limits its movement between cells [33]. By inducing cell death at the sites of infection and restricting the movement between cells, the Ny-1 gene confers potato virus Y (PVY) resistance in potatoes [34].
3.1.6. Autophagy as Antiviral Mechanism against Plant Viruses
Proteins and defective organelles are transferred to vacuoles or lysosomes for destruction through autophagy, which is an evolutionarily conserved intracellular degradation mechanism [35]. Autophagy also contributes to antiviral defence mechanisms in plants. Virus particles upon entry are recognised by specific autophagy receptors (AR). After recognition, autophagy-related genes (ATGs) get activated and initiate the formation of autophagosome, which ultimately fuse with the vacuole leading to the vacuolar degradation of autophagosome contents which also harbour viral particles (Figure 2 (4)). As TMV aggregates at infection sites in Beclin-1 or ATG7 silenced plants, it has been hypothesised that autophagy contributes to plant defence against viruses. It is interesting to note that induction of autophagy extends beyond the TMV infection sites to uninfected neighbouring tissues, where it prevents cell death [36]. Several reports have recently emphasised the significance of this mechanism in restricting virus multiplication. To prevent the replication of the cotton leaf curl Multan virus (CLCuMuV), ATG8 specifically interacts with the beta-satellite (βC1) of the CLCuMuV [37]. Not only DNA viruses but RNA viruses are also targeted by the antiviral effects of autophagy. Nuclear inclusion protein B (NIb) of turnip mosaic virus (TuMV), interacts with autophagy-related gene 6 (ATG6, also known as Beclin1) to prevent viral replication [38]. Additionally, it was reported that TuMV infection triggered autophagy, which reduced viral RNA accumulation in Arabidopsis. [39].
3.1.7. Cross Protection
Cross protection has been utilised to manage viral diseases together with protein and RNA-mediated resistance. By infecting the host plant with a mild strain of the virus, the resistance is conferred effectively. The infection caused by attenuated strain makes the affected plant immune to further infections by a viral strain that is closely related to the inoculated virus. After the attenuated strain has been inoculated, it may trigger strain or sequence-specific resistance against the challenger virus. In a number of experiments, the effect of coat protein on cross-protection reactions has been examined. When systemically expressed using PVX as a virus vector, various TMV CP mutants with altered CP aggregation demonstrated that the CP mutants with high assembly capacity offered effective cross protection against TMV infections. TMV cross protection and CP-mediated resistance may therefore depend on CP’s capacity to prevent the challenging virus from uncoating its capsid [40]. Numerous researchers have also discovered that mutations to virus silencing suppressors can reduce the severity of symptoms. For instance, mutations in the 126 kD replicase, a suppressor of silencing from the pepper mild mottle virus (PMMoV), resulted in milder symptoms as well as offering pepper plants resistance through cross protection [41].
3.2. Engineered Resistance to Plant Viruses
Engineered resistance against plant viruses can be divided into two parts: pathogen-derived resistance (PDR) and gene editing technologies.
3.2.1. Pathogen-Derived Resistance
The majority of the functions played by plant viruses during the replication cycle are well-characterised since they have short genomes and few genes. Viruses are suitable targets for developing artificial resistance based on the concept of pathogen-derived resistance (PDR). Sanford and Johnston were the ones who originally introduced this idea [42]. The most popular and effective method of obtaining PDR has been the exploitation of virus CP genes. High levels of resistance in transgenic plants have been shown to result from host plants expressing the CP of a number of RNA viruses including TMV, PVX, CMV, and TRV. This suggests that CP-mediated resistance is dependent on inhibiting the disassembly of the infecting virus. A noteworthy achievement is the use of transgenic papaya expressing the CP transgenes of papaya ring spot virus for the control of papaya ringspot disease in Hawaii [43]. Some reported cases of PDR in crop plants have been enlisted in Table 1.
3.2.2. Gene Editing Technologies
These are further divided into Engineering ZFN or TALEN-based resistance and CRISPR/Cas technology.
Engineering ZFN or TALEN-Based Resistance against Viruses
It only became possible a decade ago to manipulate the genetic material in many cellular organisms, thanks to a novel technique called genome editing. The first-generation genome editing technologies are zinc finger nucleases (ZFNs) and transcription activator-like effector nucleases (TALENs) [52]. TALENs and ZFNs are chimeric proteins formed by joining the nonspecific cleavage domain of the enzyme FokI to the DNA binding domain (DBD) from either a transcription activator-like effector or a zinc finger protein, respectively. The cleavage domain cuts the DNA to create double-strand breaks (DSB) at the target site once the DBD detects a particular recognition sequence in the target DNA. In addition to integrating, deleting, or mutating desired genes, these genome editing techniques also give a potent weapon in the armoury against plant viruses. An artificial zinc finger protein (AZP) was designed to target the intergenic region (IR) of the beet-severe curly top virus (BSCTV) in Arabidopsis [53]. This protein differs from ZFN in that it lacks the cleavage domain. The stem-loop structure found in the geminiviruses IR is critical for virus replication, which initiates with the binding of viral replication initiation protein (Rep) [54]. The BSCTV IR is efficiently bound by the transgenically produced AZP, prevents Rep binding and thereby inhibits virus infection. ZFN technology has been reported to target the Rep genes tobacco curly shoot virus (TbCSV) and tomato yellow leaf curl China virus (TYLCCNV) in tobacco plants, which showed a considerable suppression of viral genome replication [55].
Engineering CRISPR/CAS-Based Resistance against Plant Viruses
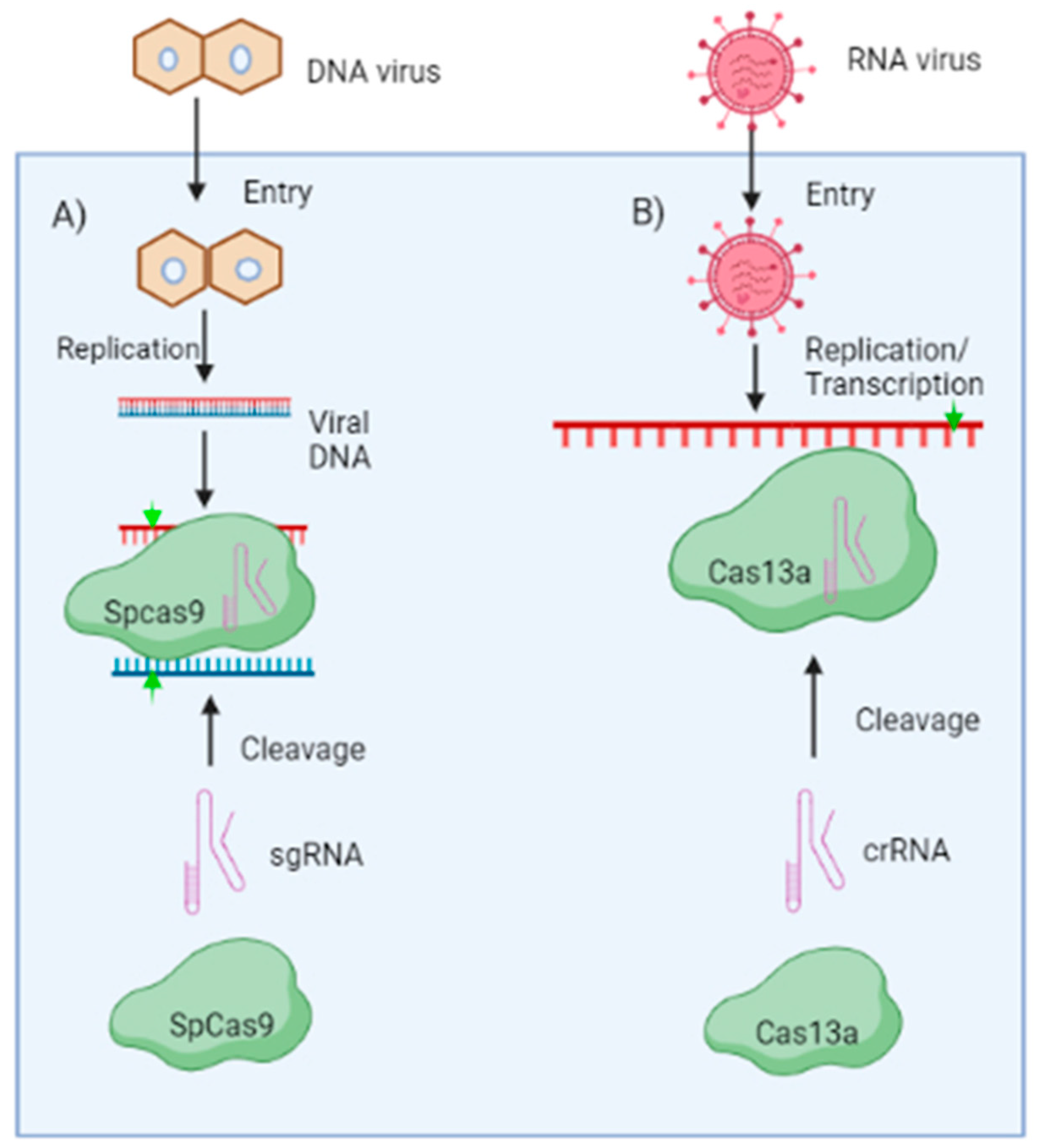
A promising tool for plant genome modification is the CRISPR (clustered regularly interspaced palindromic repeats)/CRISPR associated 9 (CRISPR/Cas9) system. The Cas9 nuclease and a single guide RNA (sgRNA) with a 20-nucleotide-long spacer sequence which guides the Cas protein to the DNA or RNA target make up the CRISPR/Cas machinery. Recently, various research has revealed the generation of virus-resistant plants with CRISPR/Cas9 technique having resistance durability [56]. By disrupting viral genes rather than silencing them at the RNA level, CRISPR/Cas9 technique provides an extra benefit to RNA interference (RNAi) or artificial microRNAs (amiRNAs) for generating virus-resistant plants [57]. The first system to be used as a platform for genetic engineering was the Class II type II CRISPR/Cas9 technology, having a single Cas9 nuclease. The Cas9 is directed by two small RNAs, transactivating CRISPR RNA (tracrRNA) and CRISPR RNA (crRNA). sgRNA is generated by the fusion of the tracrRNA and crRNA, which guides Cas9 to identify and degrade the target sequence [58] (Figure 3). This two-component system’s effectiveness and simplicity allow for developing virus resistance in different plant genera (Table 2).
CRISPR/CAS-Based Resistance against DNA Viruses
DNA is the target molecule for the CRISPR/Cas9 technology derived from Streptococcus pyrogenes (Figure 3A). In order to inhibit the geminiviruses, the CRISPR/Cas9 technology was initially used to target its genomic DNA during the replication stage. It was demonstrated that numerous geminiviruses might be simultaneously targeted by a sgRNA, which targets the conserved sequence (TAATATTAC) in the IR, which is an essential site for replication initiation [59]. In tobacco and Arabidopsis, CRISPR/Cas9 was successfully used by three different research teams to develop geminivirus resistance. sgRNAs that specifically target the IR, Rep, or CP loci were designed, and these greatly diminished geminivirus disease symptoms in plants [59,60,61]. To provide resistance to the sense and anti-sense regions of the tomato yellow leaf curl virus (TYLCV), CRISPR/Cas9 cassettes were developed for Nicotiana benthamiana. It is noteworthy that the virus was hindered by the CRISPR/Cas9 system, which also significantly lowered viral titre in leaves and caused reduced viral symptoms. The virus genome was modified by CRISPR/Cas9 through the error-prone non-homologous end joining repair (NHEJ) after it targeted the viral dsDNA for cleavage [62]. A similar technique was recently employed in barley as well, establishing highly effective resistance against the wheat dwarf virus (WDV). In order to develop resistance against multiple virus strains, the genome sequences of two barley and two wheat WDV strains were examined to identify potential sgRNA target sites situated in conserved regions. Transgenic barley lines harbouring sgRNA constructs 1,3 and 4 exhibited no symptoms and the presence of the virus was neither detected by northern blotting nor by polymerised chain reaction (PCR) [63]. CRISPR/Cas9 has also been found efficient in reducing the virulence of the cauliflower mosaic virus in Arabidopsis. It was reported that Arabidopsis transgenic plants involve the expression of several sgRNAs that target the CaMV CP coding sequences. Moreover, it was found that siRNAs with 21–24 nucleotides (nt) were produced from sgRNAs, most of which were mapped to 3’ end of the sgRNA backbone region and less frequently to the spacer region, which binds to the CaMV coat protein-coding sequences. This finding proved CRISPR/Cas9 effectiveness against double-stranded DNA viruses as well [64].
CRISPR/CAS-Based Resistance against RNA Viruses
Many viruses that infect both humans and plants have RNA genomes, which lack any DNA intermediates in their life cycle. As a result, they are immune to the traditional DNA targeting CRISPR/Cas9 mechanism. Francisella novicida, a pathogenic bacterium, whose FnCas9 orthologue targets RNA has been exploited to neutralise various human and/or plant viruses. Numerous Cas protein variants, including the Cas9 from Francisella novicida (FnCas9) and Cas13a from Leptotrichia shahii (LshCas13a) or Leptotrichia wadei (LwaCas13a), have been reported to target RNA in vivo, which shows promising results against RNA viruses (Figure 3B). For instance, CMV and TMV were the targets for modified FnCas9 and its sgRNA, in tobacco and Arabidopsis having the antiviral mechanisms, respectively. It caused lesser virus accumulation and resulted in reduced symptoms in tobacco and Arabidopsis [65]. It is interesting to note that virus inhibition by FnCas9 requires RNA-binding abilities rather than its cleavage mechanism. The LshCas13a system was designed to destroy the genomic RNA of southern rice black-streaked dwarf virus (SRBSDV) and rice stripe mosaic virus (RSMV) in rice, as well as to cleave the genomic RNA of TMV in tobacco [66]. Although these systems might result in non-specific RNA cleavage, RNA genomes as target sites are preferable because they would not cause any heritable off-target effects on the host genomic DNA. Furthermore, the RNAi system of plants will destroy the ‘cleaved off’ genomic RNAs, thus giving RNA viruses very less chance to avoid the CRISPR/Cas system.

{kind=link}
{kind=link}
{kind=link}
Table 2.
CRISPR/Cas-mediated resistance against viruses in different plant genera. Cas9 obtained from Streptococcus pyrogenes (SpCas9), FnCas9 from Francisella novicida, and Cas13a from Leptotrichia shahii (LshCas13a).
Table 2.
CRISPR/Cas-mediated resistance against viruses in different plant genera. Cas9 obtained from Streptococcus pyrogenes (SpCas9), FnCas9 from Francisella novicida, and Cas13a from Leptotrichia shahii (LshCas13a).
| CRISPR/Cas System | Host Plant | Virus | References |
|---|---|---|---|
| SpCas9 | Tobacco and Arabidopsis | Beet-severe curly top virus, Beet curly top virus | [59,61] |
| Tobacco | Bean yellow dwarf virus | [60] | |
| Barley | Wheat dwarf virus | [63,64] | |
| FnCas9 | Tobacco | Tobacco mosaic virus | [65] |
| Tobacco and Arabidopsis | Cucumber mosaic virus | [65] | |
| Tobacco | Tobacco mosaic virus | [66] | |
| LshCas13a | Rice | Southern rice blacked streaked dwarf virus | [66] |
| Tobacco | Turnip mosaic virus | [67] |
4. Reported Resistance Genes (Dominant and/or Recessive) against Plant Viruses in Major Vegetable Crops
Among plant pathogens, viruses are known to cause major yield losses to most of the significant field and horticultural crops around the world. As a result, considerable effort has been put into breeding for virus resistance. The most recent developments in molecular biology techniques have made it possible to engineer virus resistance in plants. The initial step in a resistant breeding programme is to identify a resistance source of the specific virus. Thereafter, the breeding strategy for a crop is chosen to keep in view the biology of its reproduction, type of cultivar, and inheritance of the resistance (i.e., monogenic; oligogenic or polygenic; dominant or recessive).
It was found that around 80% of the host-virus combinations have monogenic resistance, with the remaining 20% being either oligo- or polygenic [68]. Genes that confer resistance to the virus are designated as either dominant or recessive. The nucleotide-binding site leucine-rich repeat (NBS-LRR) proteins that are coded by dominant resistance genes provide resistance by interacting with the avirulence (Avr) gene products of the pathogen [69]. Members of NBS-LRR proteins can be further divided based on whether they have an N terminal coiled-coil (CC) domain or a Toll interleukin-1 receptor (TIR) homology domain. The use of dominant resistance genes conferring total resistance is an interesting option for plant breeders and they have been used widely in the breeding programme. Although the resistance activation by pathogen Avr determinants is dependent on precise molecular interactions, virus resistance genes appear to have more specificity than one might anticipate [70]. While affording a wider spectrum of protection than dominant resistance genes, recessive resistance is defined as incompatible interactions between a plant virus and its host caused by loss of or mutations in host components [15]. As viruses totally depend on their hosts to complete their life cycle, disruptions of host-virus interactions caused by mutations or deletions in host components essential for the virus infection cycle may result in recessive resistance. Host proteins necessary for intracellular and intercellular transport as well as virus replication will be the possible targets for recessive resistance genes [71]. In the following section, some of the reported dominant and/or recessive virus resistance genes in major vegetable crops have been enlisted (Table 3, Table 4, Table 5, Table 6, Table 7 and Table 8).
5. Conclusions
Despite great progress over the past decade, many questions remain unanswered, such as the molecular mechanisms controlling incompatible virus-plant interactions, the existence of PAMPs from viruses and their associated PRRs, and the mechanisms controlling the mixed infection-assisted resistance loss in host plants. Increased understanding of the natural resistance mechanisms of plants against viruses has been attained through in-depth investigations on virus/plant interactions employing innovative technologies. Numerous antiviral defence strategies have also been identified by utilising the growing understanding of the molecular interactions between viruses and their host plants. The use of well characterised natural resistance genes that can be inserted by marker-assisted breeding techniques is very promising along with genome editing tools, especially CRISPR/Cas-mediated technologies which stand out for their simplicity, robustness, and versatility in developing virus-resistant plants.
6. Future Directions
A promising approach to building genetic resistance to viruses in plants is to use host genes with antiviral activity. However, viruses are rapidly evolving and have a remarkable ability for mutation. Although significant progress has been made over the past few years in the understanding of the structure and function of the plant-virus resistance genes and in the deployment of these genes in various fields and horticultural crops, more work still needs to be carried out. Viral disease threats are emerging, particularly in developing countries, due to the expansion of monoculturing of specific crops for which the resistance resources are still unknown. Interpretation of the techniques which are significantly promising in virus resistance, and the different pathways by which these mechanisms can be deployed in resistance breeding are definitely going to be a centre of attraction for research in the coming future.
Author Contributions
A.M. and R.B. wrote the manuscript and drew figures. A.S. gave critical suggestions throughout the review and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
Different financial grants received from ICAR, DBT, and DST are acknowledged. Financial support from ICAR to authors A.M. and R.B. in terms of fellowships (Order No. Ag. Edn. 6(27)/2014-HRD); grants from Department of Science and Technology under PURSE scheme (SR/PURSE/Phase 2/25); FIST scheme (SR/FST/LS-II/2019/561) is acknowledged.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
All authors declare no conflict of interest.
References
- Soosaar, J.L.; Burch-Smith, T.M.; Dinesh-Kumar, S.P. Mechanisms of plant resistance to viruses. Nat. Rev. Microbiol. 2005, 3, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Calil, I.P.; Fontes, E.P. Plant immunity against viruses: Antiviral immune receptors in focus. Ann. Bot. 2017, 119, 711–723. [Google Scholar] [CrossRef]
- Schwessinger, B.; Zipfel, C. News from the frontline: Recent insights into PAMP-triggered immunity in plants. Curr. Opin. Plant Biol. 2008, 11, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Wang, G.; Zhou, J.M. Receptor kinases in plant-pathogen interactions: More than pattern recognition. Plant Cell 2017, 29, 618–637. [Google Scholar] [CrossRef]
- Coll, N.S.; Epple, P.; Dangl, J.L. Programmed cell death in the plant immune system. Cell Death Differ. 2011, 18, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Mandadi, K.K.; Scholthof, K.B.G. Plant immune responses against viruses: How does a virus cause disease? Plant Cell 2013, 25, 1489–1505. [Google Scholar] [CrossRef] [PubMed]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R. Recognition of double-stranded RNA and activation of NF-κB by Toll-like receptor 3. Nature 2001, 413, 732–738. [Google Scholar] [CrossRef]
- Allan, A.C.; Lapidot, M.; Culver, J.N.; Fluhr, R. An early tobacco mosaic virus-induced oxidative burst in tobacco indicates extracellular perception of the virus coat protein. Plant Physiol. 2001, 126, 97–108. [Google Scholar] [CrossRef]
- Perraki, A.; Gronnier, J.; Gouguet, P.; Boudsocq, M.; Deroubaix, A.F.; Simon, V.; Germain, V. REM1. 3’s phospho-status defines its plasma membrane nanodomain organization and activity in restricting PVX cell-to-cell movement. PLoS Pathog. 2018, 14, e1007378. [Google Scholar] [CrossRef]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant–pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef]
- Fu, Z.Q.; Dong, X. Systemic acquired resistance: Turning local infection into global defense. Annu. Rev. Plant Biol. 2013, 64, 839–863. [Google Scholar] [CrossRef] [PubMed]
- Luna, E.; Bruce, T.J.; Roberts, M.R.; Flors, V.; Ton, J. Next-generation systemic acquired resistance. Plant Physiol. 2012, 158, 844–853. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D. Plant immune response strategies against pathogens. Plant Arch. 2020, 20, 1169–1174. [Google Scholar]
- Sanfaçon, H. Plant translation factors and virus resistance. Viruses 2015, 7, 3392–3419. [Google Scholar] [CrossRef] [PubMed]
- Truniger, V.; Aranda, M.A. Recessive resistance to plant viruses. Adv. Virus Res. 2009, 75, 119–231. [Google Scholar] [PubMed]
- Domashevskiy, A.V.; Cheng, S.Y. Translation Initiation Complex eIFiso4F Targets Pokeweed Antiviral Protein (PAP) to Selectively Depurinate Uncapped Tobacco Etch Virus (TEV) RNA. Biophys. J. 2018, 114, 442. [Google Scholar] [CrossRef]
- Pantaleo, V.; Szittya, G.; Burgyán, J. Molecular bases of viral RNA targeting by viral small interfering RNA-programmed RISC. J. Virol. 2007, 81, 3797–3806. [Google Scholar] [CrossRef]
- Calvo, M.; Martínez-Turiño, S.; García, J.A. Resistance to Plum pox virus strain C in Arabidopsis thaliana and Chenopodium foetidum involves genome-linked viral protein and other viral determinants and might depend on compatibility with host translation initiation factors. Mol. Plant-Microbe Interact. 2014, 27, 1291–1301. [Google Scholar] [CrossRef]
- Ding, S.W.; Han, Q.; Wang, J.; Li, W.X. Antiviral RNA interference in mammals. Curr. Opin. Immunol. 2018, 54, 109–114. [Google Scholar] [CrossRef]
- Baulcombe, D. RNA silencing in plants. Nature. 2004, 431, 356–363. [Google Scholar] [CrossRef]
- Llave, C. Virus-derived small interfering RNAs at the core of plant–virus interactions. Trends Plant Sci. 2010, 15, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Ghoshal, B.; Sanfaçon, H. Temperature-dependent symptom recovery in Nicotiana benthamiana plants infected with tomato ringspot virus is associated with reduced translation of viral RNA2 and requires ARGONAUTE 1. Virology 2014, 456, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Jia, T.; Chen, X. The ‘how’and ‘where’of plant micro RNAs. New Phytol. 2017, 216, 1002–1017. [Google Scholar] [CrossRef] [PubMed]
- Rhoades, M.W.; Bartel, D.P.; Bartel, B. MicroRNAs and their regulatory roles in plants. Annu. Rev. Plant Biol. 2006, 57, 19–53. [Google Scholar] [CrossRef]
- Zhang, C.; Ding, Z.; Wu, K.; Yang, L.; Li, Y.; Yang, Z.; Wu, J. Suppression of jasmonic acid-mediated defense by viral-inducible microRNA319 facilitates virus infection in rice. Mol. Plant. 2016, 9, 1302–1314. [Google Scholar] [CrossRef]
- Ranf, S.; Gisch, N.; Schäffer, M.; Illig, T.; Westphal, L.; Knirel, Y.A.; Scheel, D. A lectin S-domain receptor kinase mediates lipopolysaccharide sensing in Arabidopsis thaliana. Nat. Immunol. 2015, 16, 426–433. [Google Scholar] [CrossRef]
- Sugawara, K.; Shiraishi, T.; Yoshida, T.; Fujita, N.; Netsu, O.; Yamaji, Y.; Namba, S. A replicase of potato virus X acts as the resistance-breaking determinant for JAX1-mediated resistance. Mol. Plant-Microbe Interact. 2013, 26, 1106–1112. [Google Scholar] [CrossRef]
- Yoshida, T.; Shiraishi, T.; Hagiwara-Komoda, Y.; Komatsu, K.; Maejima, K.; Okano, Y.; Namba, S. The plant noncanonical antiviral resistance protein JAX1 inhibits potexviral replication by targeting the viral RNA-dependent RNA polymerase. J. Virol. 2019, 93, e01506-18. [Google Scholar] [CrossRef]
- Yoshii, M.; Yoshioka, N.; Ishikawa, M.; Naito, S. Isolation of an Arabidopsis thaliana mutant in which accumulation of cucumber mosaic virus coat protein is delayed. Plant J. 1998, 13, 211–219. [Google Scholar] [CrossRef]
- Gao, Z.; Johansen, E.; Eyers, S.; Thomas, C.L.; Noel Ellis, T.H.; Maule, A.J. The potyvirus recessive resistance gene, sbm1, identifies a novel role for translation initiation factor eIF4E in cell-to-cell trafficking. Plant J. 2004, 40, 376–385. [Google Scholar] [CrossRef]
- Ruffel, S.; Dussault, M.H.; Palloix, A.; Moury, B.; Bendahmane, A.; Robaglia, C.; Caranta, C. A natural recessive resistance gene against potato virus Y in pepper corresponds to the eukaryotic initiation factor 4E (eIF4E). Plant J. 2002, 32, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, S.T.; Parra, M.A.; Anderberg, R.J.; Carrington, J.C. Arabidopsis RTM1 and RTM2 genes function in phloem to restrict long-distance movement of tobacco etch virus. Plant Physiol. 2001, 127, 1667–1675. [Google Scholar] [CrossRef] [PubMed]
- Fujisaki, K.; Ishikawa, M. Identification of an Arabidopsis thaliana protein that binds to tomato mosaic virus genomic RNA and inhibits its multiplication. Virology 2008, 380, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Lukan, T.; Baebler, Š.; Pompe-Novak, M.; Guček, K.; Zagorščak, M.; Coll, A.; Gruden, K. Cell death is not sufficient for the restriction of potato virus Y spread in hypersensitive response-conferred resistance in potato. Front. Plant Sci. 2018, 9, 168. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Codogno, P. The mechanism and physiological function of macroautophagy. J. Innate Immun. 2013, 5, 427–433. [Google Scholar] [CrossRef]
- Espert, L.; Codogno, P.; Biard-Piechaczyk, M. Involvement of autophagy in viral infections: Antiviral function and subversion by viruses. J. Mol. Med. 2007, 85, 811–823. [Google Scholar] [CrossRef]
- Haxim, Y.; Ismayil, A.; Jia, Q.; Wang, Y.; Zheng, X.; Chen, T.; Liu, Y. Autophagy functions as an antiviral mechanism against geminiviruses in plants. eLife 2017, 6, e23897. [Google Scholar] [CrossRef]
- Li, F.; Zhang, C.; Li, Y.; Wu, G.; Hou, X.; Zhou, X.; Wang, A. Beclin1 restricts RNA virus infection in plants through suppression and degradation of the viral polymerase. Nat. Commun. 2018, 9, 1268. [Google Scholar] [CrossRef]
- Hafrén, A.; Üstün, S.; Hochmuth, A.; Svenning, S.; Johansen, T.; Hofius, D. Turnip mosaic virus counteracts selective autophagy of the viral silencing suppressor HCpro. Plant Physiol. 2018, 176, 649–662. [Google Scholar] [CrossRef]
- Bazzini, A.A.; Asurmendi, S.; Hopp, H.E.; Beachy, R.N. Tobacco mosaic virus (TMV) and potato virus X (PVX) coat proteins confer heterologous interference to PVX and TMV infection, respectively. J. Gen. Virol. 2006, 87, 1005–1012. [Google Scholar] [CrossRef]
- Yoon, J.Y.; Ahn, H.I.; Kim, M.; Tsuda, S.; Ryu, K.H. Pepper mild mottle virus pathogenicity determinants and cross protection effect of attenuated mutants in pepper. Virus Res. 2006, 118, 23–30. [Google Scholar] [CrossRef]
- Sanford, J.C.; Johnston, S.A. The concept of parasite-derived resistance-deriving resistance genes from the parasite’s own genome. J. Theor. Biol. 1985, 113, 395–405. [Google Scholar] [CrossRef]
- Ferreira, S.A.; Pitz, K.Y.; Manshardt, R.; Zee, F.; Fitch, M.; Gonsalves, D.E. Virus coat protein transgenic papaya provides practical control of papaya ringspot virus in Hawaii. Plant Dis. 2002, 86, 101–105. [Google Scholar] [CrossRef]
- Yang, R.; Xu, H.; Long, M.; Yu, W.; Lu, C.; Wu, G.C.; Chen, Z. Transgenic tomato plants expressing cucumber mosaic virus coat protein and their resistance to CMV. Jiangsu Nong Ye Xue Bao 1995, 11, 40–44. [Google Scholar]
- Zhu, Y.X.; Ou Yang, W.J.; Zhang, Y.F.; Chen, Z.L. Transgenic sweet pepper plants from 1103 Agrobacterium mediated transformation. Plant Cell Rep. 1996, 16, 71–75. [Google Scholar] [CrossRef]
- Kaniewski, W.K.; Thomas, P.E. The potato story. AgBioForum 2004, 7, 41–46. [Google Scholar]
- Davis, M.J.; Ying, Z. Development of papaya breeding lines with transgenic resistance to Papaya ringspot virus. Plant Dis. 2004, 88, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Malinowski, T.; Cambra, M.; Capote, N.; Zawadzka, B.; Gorris, M.T.; Scorza, R.; Ravelonandro, M. Field trials of plum clones transformed with the Plum pox virus coat protein (PPV-CP) gene. Plant Dis. 2006, 90, 1012–1018. [Google Scholar] [CrossRef]
- Faria, J.C.; Albino, M.M.; Dias, B.B.; Cançado, L.J.; da Cunha, N.B.; Silva, L.D.M.; Aragão, F.J. Partial resistance to Bean golden mosaic virus in a transgenic common bean (Phaseolus vulgaris L.) line expressing a mutated rep gene. Plant Sci. 2006, 171, 565–571. [Google Scholar] [CrossRef]
- Guo, Y.; Xu, Q.; Canzio, D.; Shou, J.; Li, J.; Gorkin, D.U.; Wu, Q. CRISPR inversion of CTCF sites alters genome topology and enhancer/promoter function. Cell 2015, 162, 900–910. [Google Scholar] [CrossRef] [PubMed]
- Ilardi, V.; Tavazza, M. Biotechnological strategies and tools for Plum pox virus resistance: Trans-, intra-, cis-genesis, and beyond. Front. Plant Sci. 2015, 6, 379. [Google Scholar] [CrossRef] [PubMed]
- Boch, J.; Scholze, H.; Schornack, S.; Landgraf, A.; Hahn, S.; Kay, S.; Bonas, U. Breaking the code of DNA binding specificity of TAL-type III effectors. Science 2009, 326, 1509–1512. [Google Scholar] [CrossRef] [PubMed]
- Sera, T. Inhibition of virus DNA replication by artificial zinc finger proteins. J. Virol. 2005, 79, 2614–2619. [Google Scholar] [CrossRef] [PubMed]
- Hanley-Bowdoin, L.; Bejarano, E.R.; Robertson, D.; Mansoor, S. Geminiviruses: Masters at redirecting and reprogramming plant processes. Nat. Rev. Microbiol. 2013, 11, 777–788. [Google Scholar] [CrossRef]
- Chen, W.; Qian, Y.; Wu, X.; Sun, Y.; Wu, X.; Cheng, X. Inhibiting replication of begomoviruses using artificial zinc finger nucleases that target viral-conserved nucleotide motif. Virus Genes 2014, 48, 494–501. [Google Scholar] [CrossRef]
- Zaidi, S.S.E.A.; Tashkandi, M.; Mansoor, S.; Mahfouz, M.M. Engineering plant immunity: Using CRISPR/Cas9 to generate virus resistance. Front. Plant Sci. 2016, 7, 1673. [Google Scholar] [CrossRef]
- Unniyampurath, U.; Pilankatta, R.; Krishnan, M.N. RNA interference in the age of CRISPR: Will CRISPR interfere with RNAi? Int. J. Mol. Sci. 2016, 17, 291. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Ali, Z.; Abulfaraj, A.; Idris, A.; Ali, S.; Tashkandi, M.; Mahfouz, M.M. CRISPR/Cas9-mediated viral interference in plants. Genome Biol. 2015, 16, 238. [Google Scholar] [CrossRef]
- Baltes, N.J.; Hummel, A.W.; Konecna, E.; Cegan, R.; Bruns, A.N.; Bisaro, D.M.; Voytas, D.F. Conferring resistance to geminiviruses with the CRISPR–Cas prokaryotic immune system. Nat. Plants 2015, 1, 15145. [Google Scholar] [CrossRef]
- Ji, X.; Zhang, H.; Zhang, Y.; Wang, Y.; Gao, C. Establishing a CRISPR–Cas-like immune system conferring DNA virus resistance in plants. Nat. Plants 2015, 1, 15144. [Google Scholar] [CrossRef] [PubMed]
- Mahas, A.; Mahfouz, M. Engineering virus resistance via CRISPR–Cas systems. Curr. Opin. Virol. 2018, 32, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kis, A.; Hamar, É.; Tholt, G.; Bán, R.; Havelda, Z. Creating highly efficient resistance against wheat dwarf virus in barley by employing CRISPR/Cas9 system. Plant Biotechnol. J. 2019, 17, 1004. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Soyars, C.L.; Li, J.; Fei, Q.; He, G.; Peterson, B.A.; Wang, X. CRISPR/Cas9-mediated resistance to cauliflower mosaic virus. Plant Direct. 2018, 2, e00047. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zheng, Q.; Yi, X.; An, H.; Zhao, Y.; Ma, S.; Zhou, G. Establishing RNA virus resistance in plants by harnessing CRISPR immune system. Plant Biotechnol. J. 2018, 16, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhao, Y.; Ye, J.; Cao, X.; Xu, C.; Chen, B.; Zhou, G. Establishing CRISPR/Cas13a immune system conferring RNA virus resistance in both dicot and monocot plants. Plant Biotechnol. J. 2019, 17, 1185. [Google Scholar] [CrossRef]
- Aman, R.; Ali, Z.; Butt, H.; Mahas, A.; Aljedaani, F.; Khan, M.Z.; Mahfouz, M. RNA virus interference via CRISPR/Cas13a system in plants. Genome Biol. 2018, 19, 1–9. [Google Scholar] [CrossRef]
- Kang, B.C.; Yeam, I.; Jahn, M.M. Genetics of plant virus resistance. Annu. Rev. Phytopathol. 2005, 43, 581–621. [Google Scholar] [CrossRef]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Maule, A.J.; Caranta, C.; Boulton, M.I. Sources of natural resistance to plant viruses: Status and prospects. Mol. Plant Pathol. 2007, 8, 223–231. [Google Scholar] [CrossRef]
- Tatineni, S.; Hein, G.L. Plant Viruses of Agricultural Importance: Current and Future Perspectives of Virus Disease Management Strategies. Phytopathology 2023, 113, 117–141. [Google Scholar] [CrossRef] [PubMed]
- Zamir, D.; Ekstein-Michelson, I.; Zakay, Y.; Navot, N.; Zeidan, M.; Sarfatti, M.; Czosnek, H. Mapping and introgression of a tomato yellow leaf curl virus tolerance gene, Ty-1. Theor. Appl. Genet. 1994, 88, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Verlaan, M.G.; Hutton, S.F.; Ibrahem, R.M.; Kormelink, R.; Visser, R.G.; Scott, J.W.; Bai, Y. The tomato yellow leaf curl virus resistance genes Ty-1 and Ty-3 are allelic and code for DFDGD-class RNA–dependent RNA polymerases. PLoS Genet. 2011, 9, e1003399. [Google Scholar] [CrossRef]
- Ji, Y.; Schuster, D.J.; Scott, J.W. Ty-3, a begomovirus resistance locus near the Tomato yellow leaf curl virus resistance locus Ty-1 on chromosome 6 of tomato. Mol. Breed. 2007, 20, 271–284. [Google Scholar] [CrossRef]
- Ji, Y.; Scott, J.W.; Schuster, D.J.; Maxwell, D.P. Molecular mapping of Ty-4, a new Tomato yellow leaf curl virus resistance locus on chromosome 3 of tomato. J. Am. Soc. Hortic. Sci. 2009, 134, 281–288. [Google Scholar] [CrossRef]
- Anbinder, I.; Reuveni, M.; Azari, R.; Paran, I.; Nahon, S.; Shlomo, H.; Levin, I. Molecular dissection of Tomato leaf curl virus resistance in tomato line TY172 derived from Solanum peruvianum. Theor. Appl. Genet. 2009, 119, 519–530. [Google Scholar] [CrossRef]
- Hutton, S.F.; Scott, J.W. Ty-6, a major begomovirus resistance gene located on chromosome 10. TGC Rep. 2014, 64, 14–18. [Google Scholar]
- Spassova, M.I.; Prins, T.W.; Folkertsma, R.T.; Klein-Lankhorst, R.M.; Hille, J.; Goldbach, R.W.; Prins, M. The tomato gene Sw5 is a member of the coiled coil, nucleotide binding, leucine-rich repeat class of plant resistance genes and confers resistance to TSWV in tobacco. Mol. Breed. 2001, 7, 151–161. [Google Scholar] [CrossRef]
- Qi, S.; Shen, Y.; Wang, X.; Zhang, S.; Li, Y.; Islam, M.; Liang, Y. A new NLR gene for resistance to tomato spotted wilt virus in tomato (Solanum lycopersicum). Theor. Appl. Genet. 2022, 135, 1493–1509. [Google Scholar] [CrossRef]
- Lv, J.; Deng, M.; Jiang, S.; Zhu, H.; Li, Z.; Wang, Z.; Zhao, K. Mapping and Function Characterization of the Tomato Spotted wilt Virus Resistance Gene SlCHS3 in Solanum Lycopersicum. Mol. Breed. 2022, 42, 55. [Google Scholar] [CrossRef]
- Pelham, J. Resistance in tomato to tobacco mosaic virus. Euphytica 1966, 15, 258–267. [Google Scholar] [CrossRef]
- Pelham, J. Strain-genotype interaction of tobacco mosaic virus in tomato. Ann. Appl. Biol. 1972, 71, 219–288. [Google Scholar] [CrossRef]
- Sharma, N.; Sahu, P.P.; Prasad, A.; Muthamilarasan, M.; Waseem, M.; Khan, Y.; Prasad, M. The Sw5a gene confers resistance to ToLCNDV and triggers an HR response after direct AC4 effector recognition. Proc. Natl. Acad. Sci. USA 2021, 118, e2101833118. [Google Scholar] [CrossRef] [PubMed]
- Bendahmane, A.; Kanyuka, K.; Baulcombe, D.C. The Rx gene from potato controls separate virus resistance and cell death responses. Plant Cell 1999, 11, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Bendahmane, A.; Querci, M.; Kanyuka, K.; Baulcombe, D.C. Agrobacterium transient expression system as a tool for the isolation of disease resistance genes: Application to the Rx2 locus in potato. Plant J. 2000, 21, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, C.; Bellin, D.; Henselewski, H.; Lehmann, W.; Schwarzfischer, J.; Valkonen, J.P.T. Marker-assisted combination of major genes for pathogen resistance in potato. Theor. Appl. Genet. 2006, 112, 1458–1464. [Google Scholar] [CrossRef]
- Saka, N. A rice (Oryza sativa L.) breeding for field resistance to blast disease (Pyricularia oryzae) in Mountainous Region Agricultural Research Institute, Aichi Agricultural Research Center of Japan. Plant Prod. Sci. 2006, 9, 3–9. [Google Scholar] [CrossRef]
- Fulladolsa, A.C.; Jansky, S.H.; Smith, D.R.; Abramczak, C.M.; Charkowski, A.O. Development and Evaluation of Four Molecular Markers Tightly Linked to the Potato Virus Y Resistance Gene Rychc in Diploid Potato Populations; American Phytopathological Society: St. Paul, MN, USA, 2017. [Google Scholar]
- Kondrák, M.; Kopp, A.; Uri, C.; Sós-Hegedűs, A.; Csákvári, E.; Schiller, M.; Bánfalvi, Z. Mapping and DNA sequence characterisation of the Rysto locus conferring extreme virus resistance to potato cultivar ‘White Lady’. PLoS ONE 2020, 15, e0224534. [Google Scholar] [CrossRef]
- Murphy, J.F.; Blauth, J.R.; Livingstone, K.D.; Lackney, V.K.; Jahn, M.K. Genetic mapping of the pvr1 locus in Capsicum spp. and evidence that distinct potyvirus resistance loci control responses that differ at the whole plant and cellular levels. Mol. Plant-Microbe Interact. 1998, 11, 943–951. [Google Scholar] [CrossRef]
- Yeam, I.; Kang, B.C.; Lindeman, W.; Frantz, J.D.; Faber, N.; Jahn, M.M. Allele-specific CAPS markers based on point mutations in resistance alleles at the pvr1 locus encoding eIF4E in Capsicum. Theor. Appl. Genet. 2005, 112, 178–186. [Google Scholar] [CrossRef]
- Kang, B.C.; Yeam, I.; Frantz, J.D.; Murphy, J.F.; Jahn, M.M. The pvr1 locus in Capsicum encodes a translation initiation factor eIF4E that interacts with Tobacco etch virus VPg. Plant J. 2005, 42, 392–405. [Google Scholar] [CrossRef] [PubMed]
- Green, S.K.; Kim, J.S. Sources of Resistance to Viruses of Pepper (Capsicum spp.): A Catalog; (No. 633.8498/G798); AVRDC: Taipei, Taiwan, 1998. [Google Scholar]
- Parrella, G.; Ruffel, S.; Moretti, A.; Morel, C.; Palloix, A.; Caranta, C. Recessive resistance genes against potyviruses are localized in colinear genomic regions of the tomato (Lycopersicon spp.) and pepper (Capsicum spp.) genomes. Theor. Appl. Genet. 2002, 105, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Caranta, C.; Thabuis, A.; Palloix, A. Development of a CAPS marker for the Pvr4 locus: A tool for pyramiding potyvirus resistance genes in pepper. Genome 1999, 42, 1111–1116. [Google Scholar] [CrossRef] [PubMed]
- Grube, R.C.; Blauth, J.R.; Andrés, A.; Caranta, C.; Jahn, M.K. Identification and comparative mapping of a dominant potyvirus resistance gene cluster in Capsicum. Theor. Appl. Genet. 2000, 101, 852–859. [Google Scholar] [CrossRef]
- Caranta, C.; Palloix, A. Both common and specific genetic factors are involved in polygenic resistance of pepper to several potyviruses. Theor. Appl. Genet. 1996, 92, 15–20. [Google Scholar] [CrossRef]
- Koeda, S.; Onouchi, M.; Mori, N.; Pohan, N.S.; Nagano, A.J.; Kesumawati, E. A recessive gene pepy-1 encoding Pelota confers resistance to begomovirus isolates of PepYLCIV and PepYLCAV in Capsicum annuum. Theor. Appl. Genet. 2021, 134, 2947–2964. [Google Scholar] [CrossRef]
- Pérez-de-Castro, A.; Esteras, C.; Alfaro-Fernández, A.; Daròs, J.A.; Monforte, A.J.; Picó, B.; Gómez-Guillamón, M.L. Fine mapping of wmv1551, a resistance gene to Watermelon mosaic virus in melon. Mol. Breed. 2019, 39, 1–15. [Google Scholar] [CrossRef]
- Gilbert, R.Z.; Kyle, M.M.; Munger, H.M.; Gray, S.M. Inheritance of resistance to Watermelon mosaic virus in Cucumis melo L. HortScience 1994, 29, 107–110. [Google Scholar] [CrossRef]
- Anagnostou, K.; Jahn, M.; Perl-Treves, R. Inheritance and linkage analysis of resistance to zucchini yellow mosaic virus, watermelon mosaic virus, papaya ringspot virus and powdery mildew in melon. Euphytica 2000, 116, 265–270. [Google Scholar] [CrossRef]
- Brotman, Y.; Normantovich, M.; Goldenberg, Z.; Zvirin, Z.; Kovalski, I.; Stovbun, N.; Perl-Treves, R. Dual resistance of melon to Fusarium oxysporum races 0 and 2 and to Papaya ring-spot virus is controlled by a pair of head-to-head-oriented NB-LRR genes of unusual architecture. Mol. Plant 2013, 6, 235–238. [Google Scholar] [CrossRef]
- Pitrat, M.; Lecoq, H. Two alleles for watermelon mosaic virus 1 resistance in melon. Cucurbit Genet. Coop. 1983, 6, 52–53. [Google Scholar]
- Pitrat, M.; Lecoq, H. Inheritance of zucchini yellow mosaic virus resistance in Cucumis melo L. Euphytica 1984, 33, 57–61. [Google Scholar] [CrossRef]
- Danin-Poleg, Y.; Paris, H.S.; Cohen, S.; Rabinowitch, H.D.; Karchi, Z. Oligogenic inheritance of resistance to zucchini yellow mosaic virus in melons. Euphytica 1997, 93, 331–337. [Google Scholar] [CrossRef]
- Dhillon, N.P.S.; Ranjana, R.; Singh, K.; Eduardo, I.; Monforte, A.J.; Pitrat, M.; Singh, P.P. Diversity among landraces of Indian snapmelon (Cucumis melo var. momordica). Genet. Resour. Crop Evol. 2007, 54, 1267–1283. [Google Scholar] [CrossRef]
- Karchi, Z. Inheritance of resistance to cucumber mosaic virus in melons. Phytopathology 1975, 65, 479–481. [Google Scholar] [CrossRef]
- Pascual, L.; Yan, J.; Pujol, M.; Monforte, A.J.; Picó, B.; Martín-Hernández, A.M. CmVPS41 Is a general gatekeeper for resistance to Cucumber mosaic virus phloem entry in melon. Front. Plant Sci. 2019, 10, 1219. [Google Scholar] [CrossRef] [PubMed]
- Essafi, A.; Díaz-Pendón, J.A.; Moriones, E.; Monforte, A.J.; Garcia-Mas, J.; Martín-Hernández, A.M. Dissection of the oligogenic resistance to Cucumber mosaic virus in the melon accession PI 161375. Theor. Appl. Genet. 2009, 118, 275–284. [Google Scholar] [CrossRef]
- Guiu-Aragonés, C.; Monforte, A.J.; Saladié, M.; Corrêa, R.X.; Garcia-Mas, J.; Martín-Hernández, A.M. The complex resistance to cucumber mosaic cucumovirus (CMV) in the melon accession PI161375 is governed by one gene and at least two quantitative trait loci. Mol. Breed. 2014, 34, 351–362. [Google Scholar] [CrossRef]
- Daryono, B.S.; Somowiyarjo, S.; Natsuaki, K.T. New source of resistance to Cucumber mosaic virus in melon. SABRAO J. Breed. Genet. 2003, 35, 19–26. [Google Scholar]
- Pitrat, M.; Wipf-Scheibel, C.; Besombes, D.; Desbiez, C.; Lecoq, H. Resistance of melon to Cucumber Vein Yellowing Virus (CVYV). In Cucurbitaceae 2012, Proceedings of the Xth EUCARPIA Meeting on Genetics and Breeding of Cucurbitaceae, Antalya, Turkey, 15–18 October 2012; HAL: Antalya, Turkey, 2012; pp. 157–164. [Google Scholar]
- Romay, G.; Pitrat, M.; Lecoq, H.; Wipf-Scheibel, C.; Millot, P.; Girardot, G.; Desbiez, C. Resistance against melon chlorotic mosaic virus and tomato leaf curl New Delhi virus in melon. Plant Dis. 2019, 103, 2913–2919. [Google Scholar] [CrossRef]
- Sugiyama, M.; Ohara, T.; Sakata, Y. Inheritance of resistance to Cucumber green mottle mosaic virus in Cucumis melo L. ‘Chang Bougi’. J. Jpn. Soc. Hortic. Sci. 2007, 76, 316–318. [Google Scholar] [CrossRef]
- Provvidenti, R. Sources of resistance to viruses in two accessions of Cucumis sativus. Rep. Cucurbit Genet. Coop. USA 1985, 8, 12. [Google Scholar]
- Ramírez-Madera, A.O.; Havey, M.J. Different haplotypes encode the same protein for independent sources of Zucchini Yellow Mosaic Virus resistance in cucumber. HortScience 2017, 52, 1040–1042. [Google Scholar] [CrossRef]
- Cohen, S.; Gertman, E.; Kedar, N. Inheritance of resistance to melon mosaic virus in cucumbers. Phytopathology 1971, 61, 253–255. [Google Scholar] [CrossRef]
- Wai, T.; Grumet, R. Inheritance of resistance to watermelon mosaic virus in the cucumber line TMG-1: Tissue-specific expression and relationship to zucchini yellow mosaic virus resistance. Theor. Appl. Genet. 1995, 91, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Tian, G.; Yang, Y.; Zhang, S.; Miao, H.; Lu, H.; Wang, Y.; Gu, X. Genetic analysis and gene mapping of papaya ring spot virus resistance in cucumber. Mol. Breed. 2015, 35, 1–9. [Google Scholar] [CrossRef]
- Grumet, R.; Kabelka, E.; McQueen, S.; Wai, T.; Humphrey, R. Characterization of sources of resistance to the watermelon strain of Papaya ringspot virus in cucumber: Allelism and co-segregation with other potyvirus resistances. Theor. Appl. Genet. 2000, 101, 463–472. [Google Scholar] [CrossRef]
- Kabelka, E.; Grumet, R. Inheritance of resistance to the Moroccan watermelon mosaic virus in the cucumber line TMG-1 and cosegregation with zucchini yellow mosaic virus resistance. Euphytica 1997, 95, 237–242. [Google Scholar] [CrossRef]
- Wal, T.; Staub, J.E.; Kabelka, E.; Grumet, R. Linkage analysis of potyvirus resistance alleles in cucumber. J. Hered. 1997, 88, 454–458. [Google Scholar] [CrossRef]
- Shi, L.; Yang, Y.; Xie, Q.; Miao, H.; Bo, K.; Song, Z.; Gu, X. Inheritance and QTL mapping of cucumber mosaic virus resistance in cucumber (Cucumis sativus L.). PLoS ONE 2018, 13, e0200571. [Google Scholar] [CrossRef]
- Sugiyama, M.; Okuda, M.; Sakata, Y. Evaluation of resistance to melon yellow spot virus in a cucumber germplasm collection. Plant Breed. 2009, 128, 696–700. [Google Scholar] [CrossRef]
- Sugiyama, M.; Kawazu, Y.; Fukino, N.; Yoshioka, Y.; Shimomura, K.; Sakata, Y.; Okuda, M. Mapping of quantitative trait loci for Melon yellow spot virus resistance in cucumber (Cucumis sativus L.). Euphytica 2015, 205, 615–625. [Google Scholar] [CrossRef]
- Eid, S.; Abou-Jawdah, Y.; El-Mohtar, C.; Sobh, H.; Havey, M. Tolerance in Cucumber to Cucurbit yellow stunting disorder virus. Plant Dis. 2006, 90, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Pujol, M.; Alexiou, K.G.; Fontaine, A.S.; Mayor, P.; Miras, M.; Jahrmann, T.; Aranda, M.A. Mapping cucumber vein yellowing virus resistance in cucumber (Cucumis sativus L.) by using BSA-seq analysis. Front. Plant Sci. 2019, 10, 1583. [Google Scholar] [CrossRef]
- Brown, R.N.; Bolanos-Herrera, A.; Myers, J.R.; Miller Jahn, M. Inheritance of resistance to four cucurbit viruses in Cucurbita moschata. Euphytica 2003, 129, 253–258. [Google Scholar] [CrossRef]
- Paris, H.S.; Cohen, S.; Burger, Y.; Yoseph, R. Single-gene resistance to zucchini yellow mosaic virus in Cucurbita moschata. Euphytica 1988, 37, 27–29. [Google Scholar] [CrossRef]
- Paris, H.S.; Cohen, S. Oligogenic inheritance for resistance to zucchini yellow mosaic virus in Cucurbita pepo. Ann. Appl. Biol. 2000, 136, 209–214. [Google Scholar] [CrossRef]
- Wessel-Beaver, L. Cultivar and germplasm release. Release of ‘Soler’tropical pumpkin. J. Agric. Univ. Puerto Rico 2005, 89, 263–266. [Google Scholar]
- Provvidenti, R. New American Summer Squash Cultivars Possessing a High Level of Resistance to a Strata of Zucchini Yellow Mosaic Virus from China. Cucurbit Genet. Coop. Rep. 1997, 20, 57–58. [Google Scholar]
- Gilbert-Albertini, F.; Lecoq, H.; Pitrat, M.; Nicolet, J.L. Resistance of Cucurbita moschata to watermelon mosaic virus type 2 and its genetic relation to resistance to zucchini yellow mosaic virus. Euphytica 1993, 69, 231–237. [Google Scholar] [CrossRef]
- Miranda-Vélez, M.A.; Wessel-Beaver, L.; Rodrigues, J.C.V. Non-transmission of ZYMV and PRSV through resistant Cucurbita moschata genotypes ‘Nigerian Local’and ‘Menina’. Cucurbit Genet. Coop. Rep. 2019, 42, 37. [Google Scholar]
- Saha, D.; Rana, R.S.; Sureja, A.K.; Verma, M.; Arya, L.; Munshi, A.D. Cloning and characterization of NBS-LRR encoding resistance gene candidates from Tomato Leaf Curl New Delhi Virus resistant genotype of Luffa cylindrica Roem. Physiol. Mol. Plant Pathol. 2013, 81, 107–117. [Google Scholar] [CrossRef]
- Shrestha, S.; Michael, V.N.E.; Fu, Y.; Meru, G. Genetic Loci Associated with Resistance to Zucchini Yellow Mosaic Virus in Squash. Plants 2021, 10, 1935. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Pathak, M.; Singla, D.; Sharma, A.; Chhuneja, P.; Sarao, N.K. High-density GBS-based genetic linkage map construction and QTL identification associated with yellow mosaic disease resistance in bitter gourd (Momordica charantia L.). Front. Plant Sci. 2021, 12, 671620. [Google Scholar] [CrossRef] [PubMed]
- Provvidenti, R.; Alconero, R. Sources of resistance to pathotypes of pea seed-borne mosaic virus in the US plant introductions of Pisum sativum. PNL 1988, 20, 30–31. [Google Scholar]
- Smýkal, P.; Šafářová, D.; Navrátil, M.; Dostalová, R. Marker assisted pea breeding: eIF4E allele specific markers to pea seed-borne mosaic virus (PSbMV) resistance. Mol. Breed. 2010, 26, 425–438. [Google Scholar] [CrossRef]
- Ali, M.A. Genetics of resistance to the common bean mosaic virus (bean virus 1) in the bean (Phaseolus vulgaris L.). Phytopathology 1950, 40, 69–79. [Google Scholar]
- Drijfhout, E. Genetic Interaction between Phaseolus vulgaris and Bean Common Mosaic Virus with Implications for Strain Identification and Breeding for Resistance; Wageningen University and Research: Wageningen, The Netherlands, 1978. [Google Scholar]
Figure 1.
Possible ‘weak points’ in the virus infection cycle targeted by host defences: (1) Entry into plant cell through vector/pinocytosis/pollen/seed/vegetative propagation/epidermal hair/fungal parasite. (2) Uncoating of viral capsid. (3a) Early translation. (3b) Late translation. (4) Genome replication. (5) Encapsidation. (6) Cell-to-cell movement through plasmodesmata. (Figure created in Biorender.com, accessed on 8 March 2023).
Figure 1.
Possible ‘weak points’ in the virus infection cycle targeted by host defences: (1) Entry into plant cell through vector/pinocytosis/pollen/seed/vegetative propagation/epidermal hair/fungal parasite. (2) Uncoating of viral capsid. (3a) Early translation. (3b) Late translation. (4) Genome replication. (5) Encapsidation. (6) Cell-to-cell movement through plasmodesmata. (Figure created in Biorender.com, accessed on 8 March 2023).

Figure 2.
Natural antiviral defence mechanisms in plants: (1) Innate antiviral immunity; (2) Translation repression; (3) Small RNA-mediated resistance: (3a) MicroRNA mediated resistance. (3b) Small interfering RNA-mediated resistance. (4) Autophagy (Figure created in Biorender.com, accessed on 8 March 2023). CP/PAMP: Coat protein of virus/Pathogen-associated molecular patterns; PRR: Pattern recognition receptors; SERK: Somatic embryogenesis receptor kinases; PTI: PAMP triggered immunity; ETI: Effector-triggered immunity; S/RL: Sarcin/ricin loop; RIPs: Ribosome-inactivating proteins; ![Pathogens 12 00619 i001]() : depurination of S/R loop; miRNA gene/amiRNA: Micro RNA gene/artificial micro RNA; Pri-miRNA: Primary micro RNA; Pre-miRNA: Precursor micro RNA; miRNA: micro RNA; vsiRNA: Virus-derived small interfering RNA; DCL1: DICER LIKE 1; DCLs: DICER LIKE proteins; HST: HASTY; HEN1: HUA ENHANCER 1; RISK: RNA-induced silencing complex; AGO: Argonautes; EXO: Exonuclease; AR: Autophagy receptors; VF: Viral factors; ATGs: Autophagy-related genes.
: depurination of S/R loop; miRNA gene/amiRNA: Micro RNA gene/artificial micro RNA; Pri-miRNA: Primary micro RNA; Pre-miRNA: Precursor micro RNA; miRNA: micro RNA; vsiRNA: Virus-derived small interfering RNA; DCL1: DICER LIKE 1; DCLs: DICER LIKE proteins; HST: HASTY; HEN1: HUA ENHANCER 1; RISK: RNA-induced silencing complex; AGO: Argonautes; EXO: Exonuclease; AR: Autophagy receptors; VF: Viral factors; ATGs: Autophagy-related genes.
 : depurination of S/R loop; miRNA gene/amiRNA: Micro RNA gene/artificial micro RNA; Pri-miRNA: Primary micro RNA; Pre-miRNA: Precursor micro RNA; miRNA: micro RNA; vsiRNA: Virus-derived small interfering RNA; DCL1: DICER LIKE 1; DCLs: DICER LIKE proteins; HST: HASTY; HEN1: HUA ENHANCER 1; RISK: RNA-induced silencing complex; AGO: Argonautes; EXO: Exonuclease; AR: Autophagy receptors; VF: Viral factors; ATGs: Autophagy-related genes.
: depurination of S/R loop; miRNA gene/amiRNA: Micro RNA gene/artificial micro RNA; Pri-miRNA: Primary micro RNA; Pre-miRNA: Precursor micro RNA; miRNA: micro RNA; vsiRNA: Virus-derived small interfering RNA; DCL1: DICER LIKE 1; DCLs: DICER LIKE proteins; HST: HASTY; HEN1: HUA ENHANCER 1; RISK: RNA-induced silencing complex; AGO: Argonautes; EXO: Exonuclease; AR: Autophagy receptors; VF: Viral factors; ATGs: Autophagy-related genes.
Figure 2.
Natural antiviral defence mechanisms in plants: (1) Innate antiviral immunity; (2) Translation repression; (3) Small RNA-mediated resistance: (3a) MicroRNA mediated resistance. (3b) Small interfering RNA-mediated resistance. (4) Autophagy (Figure created in Biorender.com, accessed on 8 March 2023). CP/PAMP: Coat protein of virus/Pathogen-associated molecular patterns; PRR: Pattern recognition receptors; SERK: Somatic embryogenesis receptor kinases; PTI: PAMP triggered immunity; ETI: Effector-triggered immunity; S/RL: Sarcin/ricin loop; RIPs: Ribosome-inactivating proteins; ![Pathogens 12 00619 i001]() : depurination of S/R loop; miRNA gene/amiRNA: Micro RNA gene/artificial micro RNA; Pri-miRNA: Primary micro RNA; Pre-miRNA: Precursor micro RNA; miRNA: micro RNA; vsiRNA: Virus-derived small interfering RNA; DCL1: DICER LIKE 1; DCLs: DICER LIKE proteins; HST: HASTY; HEN1: HUA ENHANCER 1; RISK: RNA-induced silencing complex; AGO: Argonautes; EXO: Exonuclease; AR: Autophagy receptors; VF: Viral factors; ATGs: Autophagy-related genes.
: depurination of S/R loop; miRNA gene/amiRNA: Micro RNA gene/artificial micro RNA; Pri-miRNA: Primary micro RNA; Pre-miRNA: Precursor micro RNA; miRNA: micro RNA; vsiRNA: Virus-derived small interfering RNA; DCL1: DICER LIKE 1; DCLs: DICER LIKE proteins; HST: HASTY; HEN1: HUA ENHANCER 1; RISK: RNA-induced silencing complex; AGO: Argonautes; EXO: Exonuclease; AR: Autophagy receptors; VF: Viral factors; ATGs: Autophagy-related genes.
: depurination of S/R loop; miRNA gene/amiRNA: Micro RNA gene/artificial micro RNA; Pri-miRNA: Primary micro RNA; Pre-miRNA: Precursor micro RNA; miRNA: micro RNA; vsiRNA: Virus-derived small interfering RNA; DCL1: DICER LIKE 1; DCLs: DICER LIKE proteins; HST: HASTY; HEN1: HUA ENHANCER 1; RISK: RNA-induced silencing complex; AGO: Argonautes; EXO: Exonuclease; AR: Autophagy receptors; VF: Viral factors; ATGs: Autophagy-related genes.
Figure 3.
CRISPR/Cas-based engineered resistance mechanisms: The Cas protein and sgRNA make up the CRISPR system. The virus-targeting Cas protein and its cognate transgenic or transit expression can successfully inhibit virus infection. (A) When a DNA virus enters a plant cell, its genome is converted into dsDNA intermediate that is targeted by the Cas9 protein from Streptococcus pyrogenes (SpCas9). (B) While it has been demonstrated that Cas13a effectively confers resistance to RNA viruses. Upon entry, Cas13a can cleave/bind the virus genome under the guidance of their associated sgRNA or crRNA, respectively. The cleave sites on viral mRNA are shown by arrowheads in green. (Figure created in Biorender.com, accessed on 8 March 2023).
Figure 3.
CRISPR/Cas-based engineered resistance mechanisms: The Cas protein and sgRNA make up the CRISPR system. The virus-targeting Cas protein and its cognate transgenic or transit expression can successfully inhibit virus infection. (A) When a DNA virus enters a plant cell, its genome is converted into dsDNA intermediate that is targeted by the Cas9 protein from Streptococcus pyrogenes (SpCas9). (B) While it has been demonstrated that Cas13a effectively confers resistance to RNA viruses. Upon entry, Cas13a can cleave/bind the virus genome under the guidance of their associated sgRNA or crRNA, respectively. The cleave sites on viral mRNA are shown by arrowheads in green. (Figure created in Biorender.com, accessed on 8 March 2023).

Table 1.
Reported cases of pathogen-derived resistance in crops (against plant viruses).
| Crop Species | Target Pathogen | Gene Expressed | Reference |
|---|---|---|---|
| Solanum lycopersicum | Cucumber mosaic virus | Coat protein | [44] |
| Capsicum annum | Cucumber mosaic virus | Coat protein | [45] |
| Solanum tuberosum | Potato leaf roll virus | Coat protein | [46] |
| Carica papaya | Papaya ringspot virus | Coat protein | [47] |
| Prunus domestica | Plum pox virus | Coat protein | [48] |
| Phaseolus vulgaris | Bean golden mosaic virus | +,− RNA of virus replication protein | [49] |
| Carica papaya | Papaya ringspot virus | Viral replicase gene | [50] |
| Prunus domestica | Plum pox virus | Coat protein, P1, HC-Pro | [51] |
Table 3.
Reported resistance genes against plant viruses in tomato (Solanum lycopersicum).
| R Gene | Name of the Virus | Reference |
|---|---|---|
| Ty-1 | Tomato yellow leaf curl virus | [72] |
| Ty-2 | Tomato yellow leaf curl virus | [73] |
| Ty-3 | Tomato yellow leaf curl virus | [74] |
| Ty-4 | Tomato yellow leaf curl virus | [75] |
| ty-5 | Tomato yellow leaf curl virus | [76] |
| Ty-6 | Tomato yellow leaf curl virus | [77] |
| Sw-5 | Tomato spotted wilt virus | [78] |
| Sl5R-1 | Tomato spotted wilt virus | [79] |
| SICSH3 | Tomato spotted wilt virus | [80] |
| Tm-1 | Tomato mosaic virus | [81] |
| Tm2 and Tm-22 | Tomato mosaic virus | [82] |
| SlSw5a | Tomato leaf curl New Delhi virus | [83] |
Table 4.
Reported resistance genes against plant viruses in potato (Solanum tuberosum).
| R Gene | Name of the Virus | Reference |
|---|---|---|
| Rx-1 | Potato virus X | [84] |
| Rx-2 | Potato virus X | [85] |
| Ryadg | Potato virus Y | [86] |
| Y-1 | Potato virus Y | [87] |
| Rychc | Potato virus Y | [88] |
| Ryfsto | Potato virus YNTN | [89] |
Table 5.
Reported resistance genes against plant viruses in chilli (Capsicum annuum).
| R gene | Name of the Virus | References |
|---|---|---|
| pvr1 | Potato Virus Y, Pepper mottle virus | [90,91] |
| pvr2 | Potato Virus Y | [91,92] |
| pvr3 | Pepper mottle virus | [93,94] |
| Pvr4 | Pepper mottle virus | [95,96] |
| pvr5 | Pepper vein mottle virus | [94] |
| pvr6 | Pepper vein mottle virus | [31,97] |
| Pvr7 | Pepper mottle virus | [96] |
| pepy-1 | Pepper yellow leaf curl Indonesia virus and Pepper yellow leaf curl Aceh virus | [98] |
Table 6.
Reported resistance genes and quantitative trait loci (QTLs) identified against plant viruses in cucurbits.
Table 6.
Reported resistance genes and quantitative trait loci (QTLs) identified against plant viruses in cucurbits.
| Host Plant | Group | R Gene | Name of the Virus | References |
|---|---|---|---|---|
| Cucumis melo | Acidulus | Wmv1551 Wmr | Watermelon mosaic virus | [99,100] |
| Prv2 Prv1 | Papaya ringspot virus | [101,102,103] | ||
| Acidulus | Zym-1 to Zym-3 | Zuchhini yellow mosaic virus | [104,105,106] | |
| Conomon, Conomon | cmv1 cmv1, cmqw3.1, cmqw10.1 Creb-2 | Cucumber mosaic virus | [107,108,109,110,111] | |
| Cvy-1 cvy-2 Cvy-3 | Cucumber vein yellowing virus | [112] | ||
| bgm-1, Bgm-2, Tolcndv | Tomato leaf curl New Delhi virus | [113] | ||
| Makuwa | cgmmv-1, cgmmv-2 | Cucumber green mottle mosaic virus | [114] | |
| Cucumis sativus | zym-1 | Zuchhini yellow mosaic virus | [115,116] | |
| Wmv wmv-2, wmv-3 | Watermelon mosaic virus | [117,118] | ||
| prsv Prsv-2 prsv-1 | Papaya ringspot virus | [119,120,121,122] | ||
| cmv6.1 | Cucumber mosaic virus | [123] | ||
| Swf-1, Swf-2, Swf-3 and Swf-4 | Melon yellow spot virus | [124,125] | ||
| cysdv5.1 | Cucurbit yellow stunting disorder virus | [126] | ||
| Cscys-1 | Cucumber vein yellowing virus | [127] | ||
| Cucurbita moschata | Zym-0, zym-4, Zym-1, Zym-2, Zym-3 zym-6 | Zucchini yellow mosaic virus | [128,129,130,131] | |
| Wmv | Watermelon mosaic virus | [128,132,133] | ||
| prv | Papaya ringspot virus | [128,134] | ||
| cmv | Cucumber mosaic virus | [128,132] | ||
| Luffa cylindrica | RGCLc28 | Tomato leaf curl New Delhi virus | [135] | |
| Cucurbita pepo | QtlZYMV-02 QtlZYMV-04 QtlZYMV-08 QtlZYMV-20 | Zucchini yellow mosaic virus | [136] | |
| Momordica charantia | qYMD.pau_3.1 q.YMD.pau_4.1 qYMD.pau_5.1 | Virus complex | [137] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Majumdar, A.; Sharma, A.; Belludi, R. Natural and Engineered Resistance Mechanisms in Plants against Phytoviruses. Pathogens 2023, 12, 619. https://doi.org/10.3390/pathogens12040619
AMA Style
Majumdar A, Sharma A, Belludi R. Natural and Engineered Resistance Mechanisms in Plants against Phytoviruses. Pathogens. 2023; 12(4):619. https://doi.org/10.3390/pathogens12040619
Chicago/Turabian StyleMajumdar, Anik, Abhishek Sharma, and Rakesh Belludi. 2023. "Natural and Engineered Resistance Mechanisms in Plants against Phytoviruses" Pathogens 12, no. 4: 619. https://doi.org/10.3390/pathogens12040619
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.