
Tetraplex Fluorescent Microbead-Based Immunoassay for the Serodiagnosis of Newcastle Disease Virus and Avian Influenza Viruses in Poultry Sera

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experimentation Permits
2.2. Virus Propagation
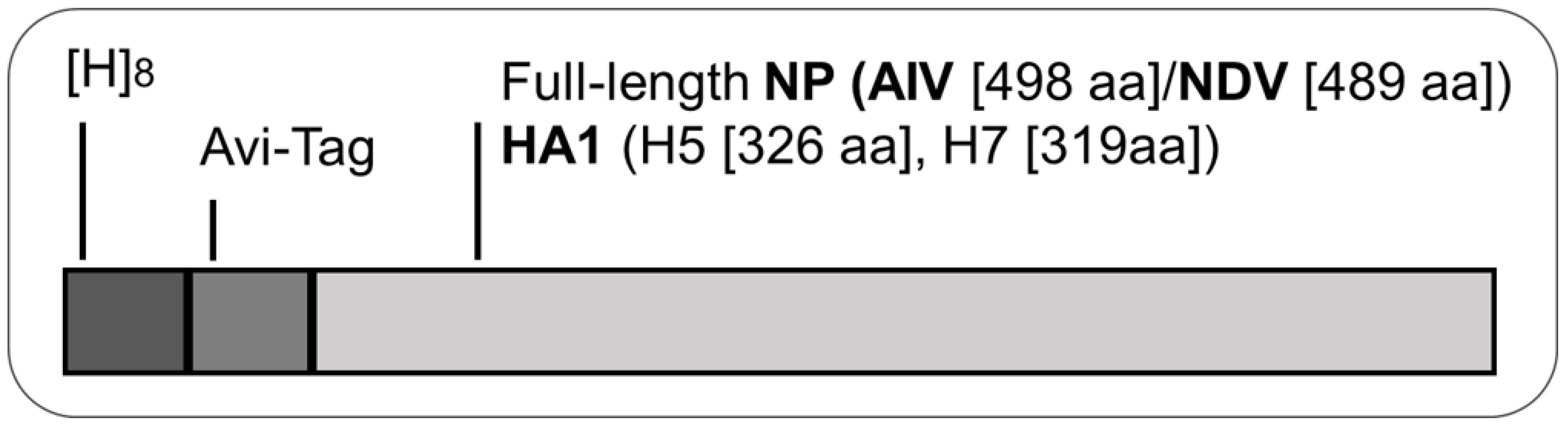
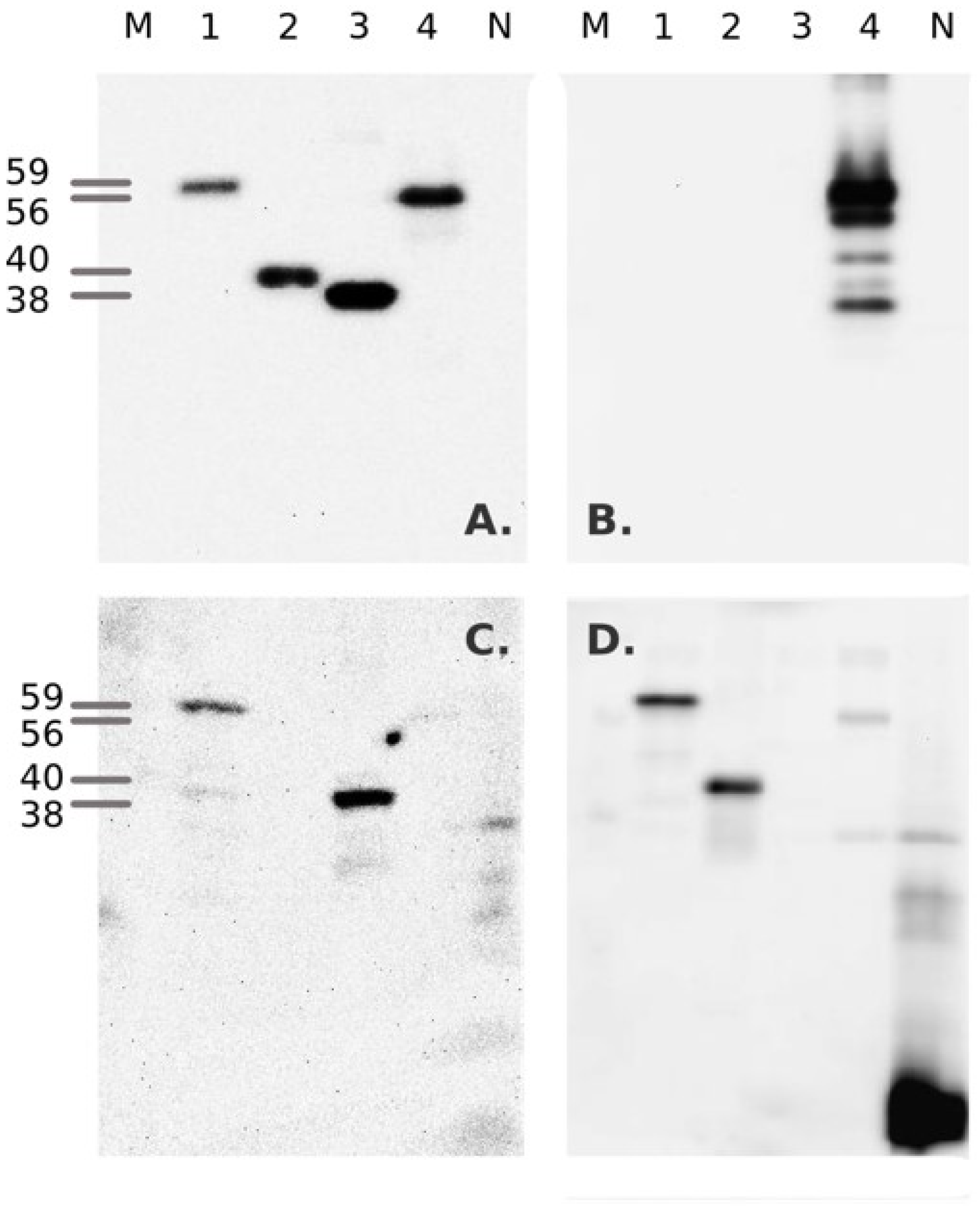
2.3. Cloning and Bacterial Expression of Recombinant Proteins
2.4. Purification and Reconstitution of Recombinant Proteins from Bacterial Inclusion Bodies
2.5. Production of Positive Control Sera
2.6. Production of Experimental Infection or Vaccination Sera in Chickens or Turkeys
2.7. Origin of Field Sera Not Used for Reference Purposes
2.8. Monoclonal Antibodies (mAbs)
2.9. Western Blotting
2.10. Coupling of Recombinant Antigens to Streptavidin Precoated Luminex Beads
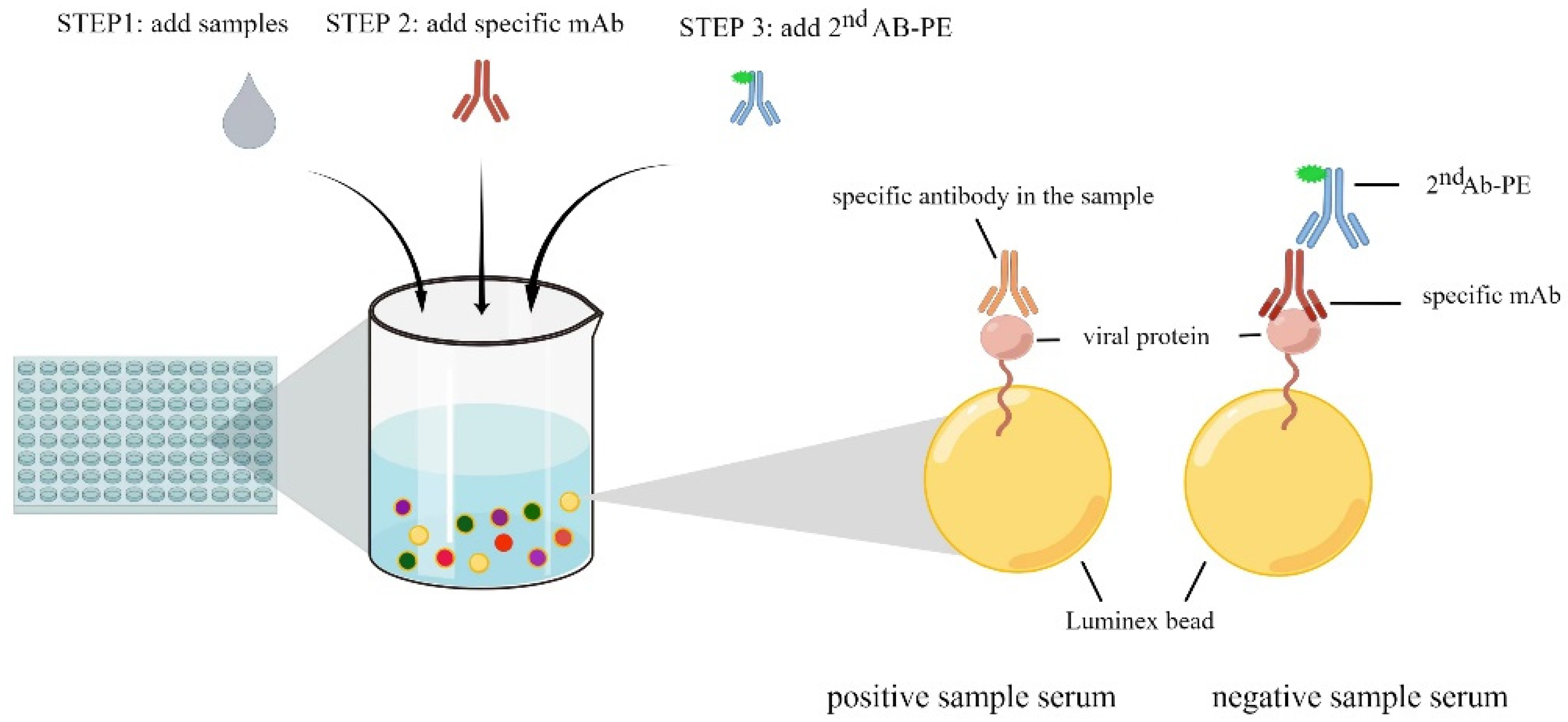
2.11. Multiplex Bead-Based Immunoassay
2.12. Commercial ELISAs for Detection of AIV and NDV Antibodies
2.13. Hemagglutination Inhibition Assay (HI)
2.14. Statistical Analysis
3. Results
3.1. Bacterial Expression of Recombinant Proteins
3.2. Development of the 4Plex iFMIA Assay
3.3. Four Plex iFMIA Cutoff Determination and Performances Compared with ELISAs and HI
3.4. Performance Characteristics of the 4Plex iFMIA Assay with Field Sera
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dimitrov, K.M.; Abolnik, C.; Afonso, C.L.; Albina, E.; Bahl, J.; Berg, M.; Briand, F.X.; Brown, I.H.; Choi, K.S.; Chvala, I.; et al. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2019, 74, 103917. [Google Scholar] [CrossRef] [PubMed]
- Spackman, E. A brief introduction to avian influenza virus. Methods Mol. Biol. 2014, 1161, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Sims, L.D. Intervention strategies to reduce the risk of zoonotic infection with avian influenza viruses: Scientific basis, challenges and knowledge gaps. Influenza Other Respir. Viruses 2013, 7 (Suppl. S2), 15–25. [Google Scholar] [CrossRef] [PubMed]
- Senne, D.A.; King, D.J.; Kapczynski, D.R. Control of Newcastle disease by vaccination. Dev. Biol. 2004, 119, 165–170. [Google Scholar]
- Steglich, C.; Grund, C.; Ramp, K.; Breithaupt, A.; Hoper, D.; Keil, G.; Veits, J.; Ziller, M.; Granzow, H.; Mettenleiter, T.C.; et al. Chimeric newcastle disease virus protects chickens against avian influenza in the presence of maternally derived NDV immunity. PLoS ONE 2013, 8, e72530. [Google Scholar] [CrossRef]
- Zhao, N.; Grund, C.; Beer, M.; Harder, T.C. Engineered recombinant protein products of the avian paramyxovirus type-1 nucleocapsid and phosphoprotein genes for serological diagnosis. Virol. J. 2018, 15, 8. [Google Scholar] [CrossRef]
- Zhao, N.; Lange, E.; Kubald, S.; Grund, C.; Beer, M.; Harder, T.C. Distinction of subtype-specific antibodies against European porcine influenza viruses by indirect ELISA based on recombinant hemagglutinin protein fragment-1. Virol. J. 2013, 10, 246. [Google Scholar] [CrossRef]
- Wood, J.M.; Major, D.; Heath, A.; Newman, R.W.; Hoschler, K.; Stephenson, I.; Clark, T.; Katz, J.M.; Zambon, M.C. Reproducibility of serology assays for pandemic influenza H1N1: Collaborative study to evaluate a candidate WHO International Standard. Vaccine 2012, 30, 210–217. [Google Scholar] [CrossRef]
- Grund, S.; Adams, O.; Wahlisch, S.; Schweiger, B. Comparison of hemagglutination inhibition assay, an ELISA-based micro-neutralization assay and colorimetric microneutralization assay to detect antibody responses to vaccination against influenza A H1N1 2009 virus. J. Virol. Methods 2011, 171, 369–373. [Google Scholar] [CrossRef]
- Jenson, T.A. Agar gel immunodiffusion assay to detect antibodies to Type A influenza virus. Methods Mol. Biol. 2014, 1161, 141–150. [Google Scholar] [CrossRef]
- Chappell, L.; Killian, M.L.; Spackman, E. Detection of influenza A antibodies in avian serum samples by ELISA. Methods Mol. Biol. 2014, 1161, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Charlton, B.; Crossley, B.; Hietala, S. Conventional and future diagnostics for avian influenza. Comp. Immunol. Microbiol. Infect. Dis. 2009, 32, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.J.; Torchetti, M.K. Newcastle disease virus detection and differentiation from avian influenza. Methods Mol. Biol. 2014, 1161, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Katz, J.M.; Hancock, K.; Xu, X. Serologic assays for influenza surveillance, diagnosis and vaccine evaluation. Expert Rev. Anti-Infect. Ther. 2011, 9, 669–683. [Google Scholar] [CrossRef] [PubMed]
- Häuslaigner, R.; Sonnenburg, J.; Kothlow, S.; Kaspers, B.; Staubach, C.; Grund, C. Evaluation of an indirect enzyme-linked immunosorbent assay to study the specific humoral immune response of Muscovy ducks (Cairina moschata) and domestic geese (Anser anser var. domestica) after vaccination against Newcastle disease virus. Avian Pathol. 2009, 38, 89–95. [Google Scholar] [CrossRef]
- Chen, R.; Yu, X.L.; Gao, X.B.; Xue, C.Y.; Song, C.X.; Li, Y.; Cao, Y.C. Bead-based suspension array for simultaneous differential detection of five major swine viruses. Appl. Microbiol. Biotechnol. 2015, 99, 919–928. [Google Scholar] [CrossRef]
- Dunbar, S.A.; Ritchie, V.B.; Hoffmeyer, M.R.; Rana, G.S.; Zhang, H. Luminex® multiplex bead suspension arrays for the detection and serotyping of Salmonella spp. Methods Mol. Biol. 2015, 1225, 1–27. [Google Scholar] [CrossRef]
- Bohm, K.; Filomena, A.; Schneiderhan-Marra, N.; Krause, G.; Sievers, C. Validation of HAV biomarker 2A for differential diagnostic of hepatitis A infected and vaccinated individuals using multiplex serology. Vaccine 2017, 35, 5883–5889. [Google Scholar] [CrossRef]
- Li, L.; Wagner, B.; Freer, H.; Schilling, M.; Bannantine, J.P.; Campo, J.J.; Katani, R.; Grohn, Y.T.; Radzio-Basu, J.; Kapur, V. Early detection of Mycobacterium avium subsp. paratuberculosis infection in cattle with multiplex-bead based immunoassays. PLoS ONE 2017, 12, e0189783. [Google Scholar] [CrossRef]
- Kamminga, S.; van der Meijden, E.; Wunderink, H.F.; Touze, A.; Zaaijer, H.L.; Feltkamp, M.C.W. Development and Evaluation of a Broad Bead-Based Multiplex Immunoassay to Measure IgG Seroreactivity against Human Polyomaviruses. J. Clin. Microbiol. 2018, 56, e01566-17. [Google Scholar] [CrossRef]
- Mechaly, A.; Vitner, E.; Levy, H.; Weiss, S.; Bar-David, E.; Gur, D.; Koren, M.; Cohen, H.; Cohen, O.; Mamroud, E.; et al. Simultaneous Immunodetection of Anthrax, Plague, and Tularemia from Blood Cultures by Use of Multiplexed Suspension Arrays. J. Clin. Microbiol. 2018, 56, e01479-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez-Munguia, I.; van Wamel, W.J.B.; Rodriguez-Ortega, M.J.; Obando, I. Detection of Natural Antibodies and Serological Diagnosis of Pneumococcal Pneumonia Using a Bead-Based High-Throughput Assay. Methods Mol. Biol. 2017, 1643, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Starick, E.; Lange, E.; Fereidouni, S.; Bunzenthal, C.; Hoveler, R.; Kuczka, A.; grosse Beilage, E.; Hamann, H.P.; Klingelhofer, I.; Steinhauer, D.; et al. Reassorted pandemic (H1N1) 2009 influenza A virus discovered from pigs in Germany. J. Gen. Virol. 2011, 92, 1184–1188. [Google Scholar] [CrossRef] [PubMed]
- Rigaut, G.; Shevchenko, A.; Rutz, B.; Wilm, M.; Mann, M.; Seraphin, B. A generic protein purification method for protein complex characterization and proteome exploration. Nat. Biotechnol. 1999, 17, 1030–1032. [Google Scholar] [CrossRef]
- Khurana, S.; Larkin, C.; Verma, S.; Joshi, M.B.; Fontana, J.; Steven, A.C.; King, L.R.; Manischewitz, J.; McCormick, W.; Gupta, R.K.; et al. Recombinant HA1 produced in E. coli forms functional oligomers and generates strain-specific SRID potency antibodies for pandemic influenza vaccines. Vaccine 2011, 29, 5657–5665. [Google Scholar] [CrossRef]
- Werner, O.; Romer-Oberdorfer, A.; Kollner, B.; Manvell, R.J.; Alexander, D.J. Characterization of avian paramyxovirus type 1 strains isolated in Germany during 1992 to 1996. Avian Pathol. 1999, 28, 79–88. [Google Scholar] [CrossRef]
- Postel, A.; Letzel, T.; Muller, F.; Ehricht, R.; Pourquier, P.; Dauber, M.; Grund, C.; Beer, M.; Harder, T.C. In vivo biotinylated recombinant influenza A virus hemagglutinin for use in subtype-specific serodiagnostic assays. Anal. Biochem. 2011, 411, 22–31. [Google Scholar] [CrossRef]
- R Foundation for Statistical Computing. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Fawcett, T. An introduction to ROC analysis. Pattern Recognit. Lett. 2006, 27, 861–874. [Google Scholar] [CrossRef]
- Phan, L.V.; Park, M.J.; Kye, S.J.; Kim, J.Y.; Lee, H.S.; Choi, K.S. Development and field application of a competitive enzyme-linked immunosorbent assay for detection of Newcastle disease virus antibodies in chickens and ducks. Poult. Sci. 2013, 92, 2034–2043. [Google Scholar] [CrossRef]
- Matsubara, K.; Iwata, S.; Nakayama, T. Antibodies against mumps virus component proteins. J. Infect. Chemother. Off. J. Jpn. Soc. Chemother. 2012, 18, 466–471. [Google Scholar] [CrossRef]
- Deregt, D.; Furukawa-Stoffer, T.L.; Tokaryk, K.L.; Pasick, J.; Hughes, K.M.; Hooper-McGrevy, K.; Baxi, S.; Baxi, M.K. A microsphere immunoassay for detection of antibodies to avian influenza virus. J. Virol. Methods 2006, 137, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Watson, D.S.; Reddy, S.M.; Brahmakshatriya, V.; Lupiani, B. A multiplexed immunoassay for detection of antibodies against avian influenza virus. J. Immunol. Methods 2009, 340, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Lupiani, B.; Mozisek, B.; Mason, P.W.; Lamichhane, C.; Reddy, S.M. Simultaneous detection of avian influenza virus NP and H5 antibodies in chicken sera using a fluorescence microsphere immunoassay. Avian Dis. 2010, 54, 668–672. [Google Scholar] [CrossRef] [PubMed]
- Pinette, M.M.; Rodriguez-Lecompte, J.C.; Pasick, J.; Ojkic, D.; Leith, M.; Suderman, M.; Berhane, Y. Development of a duplex Fluorescent Microsphere Immunoassay (FMIA) for the detection of antibody responses to influenza A and newcastle disease viruses. J. Immunol. Methods 2014, 405, 167–177. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
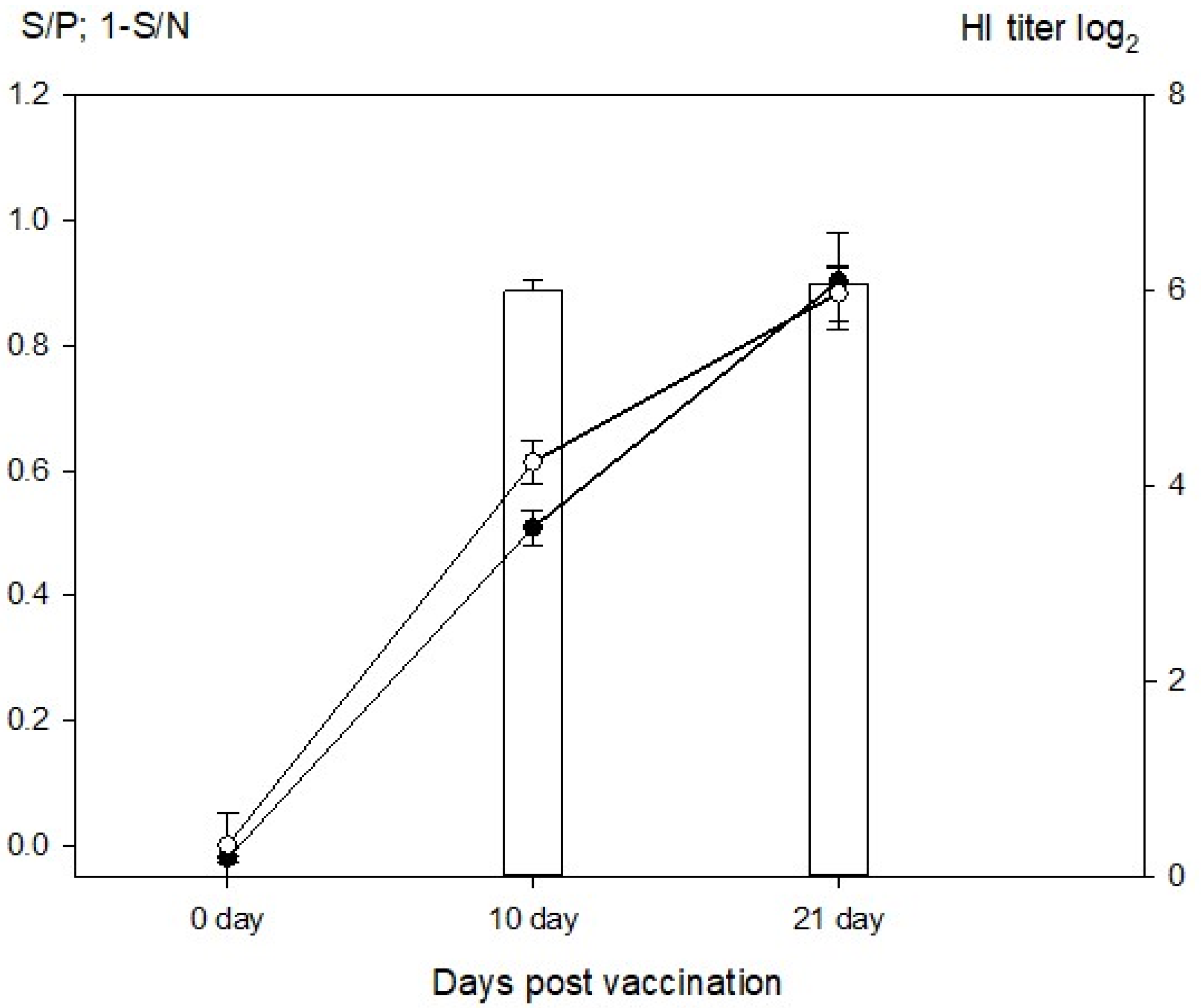{kind=link}
| Flock Immune Status | |||||||
|---|---|---|---|---|---|---|---|
| Panel | Serum Source | No. of Samples | Species | ND-NP | AI-NP | H5 | H7 |
| A | ND vaccination, EXP | 70 | chicken | + | − | − | − |
| B | AI H5/ND vaccination, EXP | 40 | chicken | + | + | + | − |
| C | AI H7 vaccination, EXP | 10 | chicken | − | + | − | + |
| D | AI H7 infection in ND vaccinated birds, EXP, Field | 11 | turkey | + | + | − | + |
| E | recND-H7 vaccination, EXP | 26 | chicken | + | − | − | + |
| F | AI H9N2 infection, EXP | 10 | chicken | − | + | − | − |
| G | AI H9N2 infection in ND vaccinated birds, EXP, Field | 10 | turkey | + | + | − | − |
| H | SPF | 50/30 | chicken/turkey | − | − | − | − |
| Target | Assay Format a | Sensitivity | 95% CI | Specificity | 95% CI | AUC b |
|---|---|---|---|---|---|---|
| NDV Full virion | iELISA (IDEXX) | 0.93 | 0.87–0.96 | 0.95 | 0.86–0.99 | |
| NDV-rNP | 4plex iFMIA | 0.93 | 0.87–0.96 | 0.91 | 0.81–0.97 | 0.937 |
| AIV Full virion | c-ELISA (iD.Vet) | 0.98 | 0.92–1 | 0.96 | 0.92–0.99 | |
| AIV-rNP | 4plex iFMIA | 0.97 | 0.91–1 | 0.96 | 0.91–0.99 | 0.987 |
| Subtype H5 virion | c-ELISA (iD.Vet) | 0.85 | 0.69–0.94 | 0.99 | 0.97–1 | |
| H5-rHA1 | 4plex iFMIA | 1 | 0.91–1 | 0.98 | 0.95–1 | 0.996 |
| Subtype H7 virion | c-ELISA (iD.Vet) | 0.9 | 0.76–0.97 | 0.99 | 0.97–1 | |
| H7-rHA1 | 4plex iFMIA | 0.95 | 0.83–1 | 0.98 | 0.94–1 | 0.986 |
| Serum Panel a | Detection Antigens | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| NDV NP | AIV NP | HA-H5 | HA-H7 | ||||||||
| HI | IDEXX | 4Plex iFMIA | iD.Vet | 4Plex iFMIA | HI | iD.Vet | 4Plex iFMIA | HI | iD.Vet | 4Plex iFMIA | |
| ND vaccination chicken | 10/10 | 9/10 | 9/10 | 0/10 | 0/10 | 0/10 | 0/10 | 0/10 | 0/10 | 0/10 | 0/10 |
| AI-H5 vaccination in ND vaccinated chicken | 39/40 | 39/40 | 33/40 | 32 (1)/40 b | 36/40 | 40/40 | 31 (2)/40 | 40/40 | 0/40 | 0/40 | 0/40 |
| AI H7 vaccination chicken | 0/10 | 0/10 | 0/10 | 10/10 | 10/10 | 0/10 | 0/10 | 0/10 | 8/10 | 5 (4)/10 | 6/10 |
| Recombinant ND-H7 vaccination chicken | 26/26 | 25/26 | 26/26 | 0/26 | 0/26 | 0/26 | 0/26 | 0/26 | 21/26 | 14 (5)/26 | 19/26 |
| Experimental AI-H7 infection in ND vaccinated turkeys | 11/11 | 6/11 | 3/11 | 11/11 | 11/11 | 0/11 | 0/11 | 0/11 | 10/11 | 7 (2)/11 | 9/11 |
| AI H9N2 infection, EXP | 0/10 | 0/10 | 0/10 | 10/10 | 10/10 | 0/10 | 0/10 | 0/10 | 0/10 | 0/10 | 0/10 |
| AI H9N2 infection in ND vaccinated birds, Field | 9/10 | 8/10 | 9/10 | 10/10 | 10/10 | 0/10 | 0/10 | 0/10 | 0/10 | 0/10 | 0/10 |
| Target | Assay | No. of Positives | Sensitivity a | 95% CI | Specificity a | 95% CI |
|---|---|---|---|---|---|---|
| NDV | HI | 129 | ||||
| iELISA (IDEXX) | 179 | 0.83 | 0.76–0.89 | 0.82 | 0.74–0.89 | |
| 4plex iFMIA | 177 | 0.96 | 0.91–0.99 | 0.56 | 0.47–0.65 | |
| AIV | c-ELISA (iD.Vet) | 88 | n.a. b | n.a. | ||
| 4plex iFMIA | 91 | 0.87 | 0.79–0.93 | 0.95 | 0.91–0.98 | |
| Subtype-H5 | HI | 0 | ||||
| c-ELISA (iD.Vet) | 0 | No HI-positives c | 1 | 0.98–1 | ||
| 4plex iFMIA | 2 | No HI-positives | 0.99 | 0.97–1 | ||
| Subtype-H7 | HI | 40 | ||||
| c-ELISA (iD.Vet) | 40 | 1 | 0.91–1 | 1 | 0.97–1 | |
| 4plex iFMIA | 42 | 1 | 0.91–1 | 0.99 | 0.97–1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, N.; Grund, C.; Beer, M.; Wang, G.; Harder, T.C. Tetraplex Fluorescent Microbead-Based Immunoassay for the Serodiagnosis of Newcastle Disease Virus and Avian Influenza Viruses in Poultry Sera. Pathogens 2022, 11, 1059. https://doi.org/10.3390/pathogens11091059
Zhao N, Grund C, Beer M, Wang G, Harder TC. Tetraplex Fluorescent Microbead-Based Immunoassay for the Serodiagnosis of Newcastle Disease Virus and Avian Influenza Viruses in Poultry Sera. Pathogens. 2022; 11(9):1059. https://doi.org/10.3390/pathogens11091059
Chicago/Turabian StyleZhao, Na, Christian Grund, Martin Beer, Gang Wang, and Timm C. Harder. 2022. "Tetraplex Fluorescent Microbead-Based Immunoassay for the Serodiagnosis of Newcastle Disease Virus and Avian Influenza Viruses in Poultry Sera" Pathogens 11, no. 9: 1059. https://doi.org/10.3390/pathogens11091059