
The N-terminal Subunit of the Porcine Deltacoronavirus Spike Recombinant Protein (S1) Does Not Serologically Cross-react with Other Porcine Coronaviruses

 , , , , , and
, , , , , and
Abstract
:1. Introduction
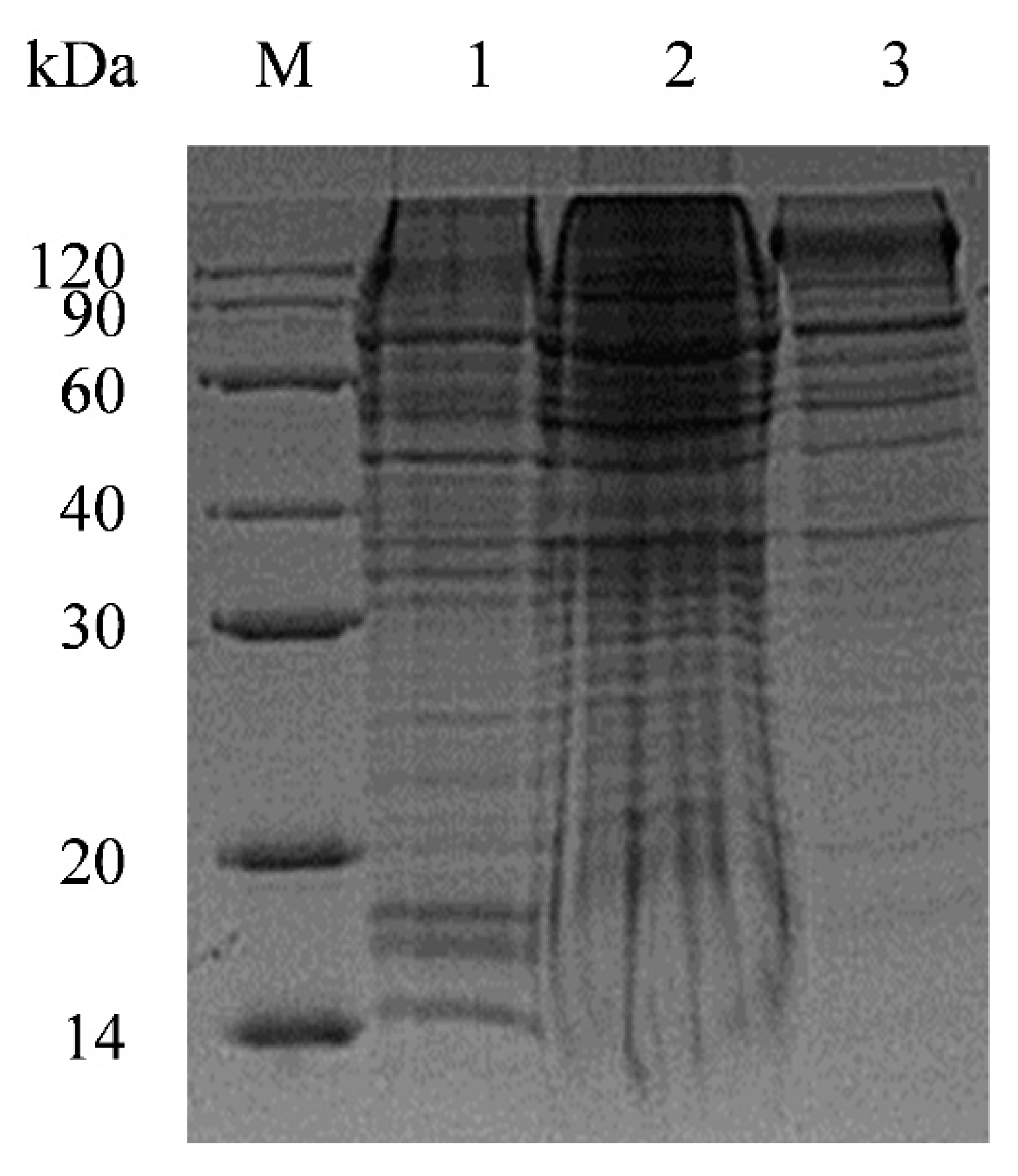
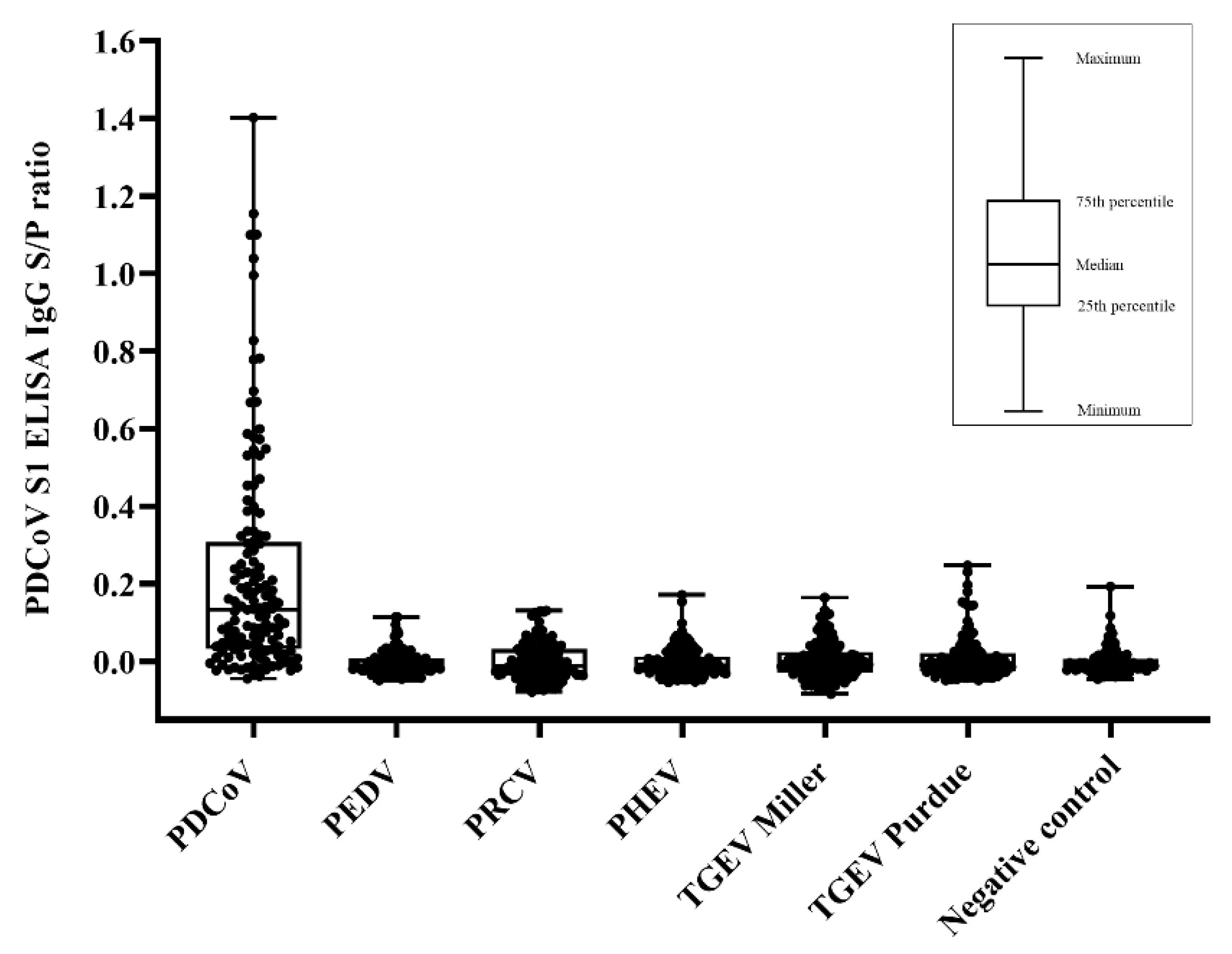
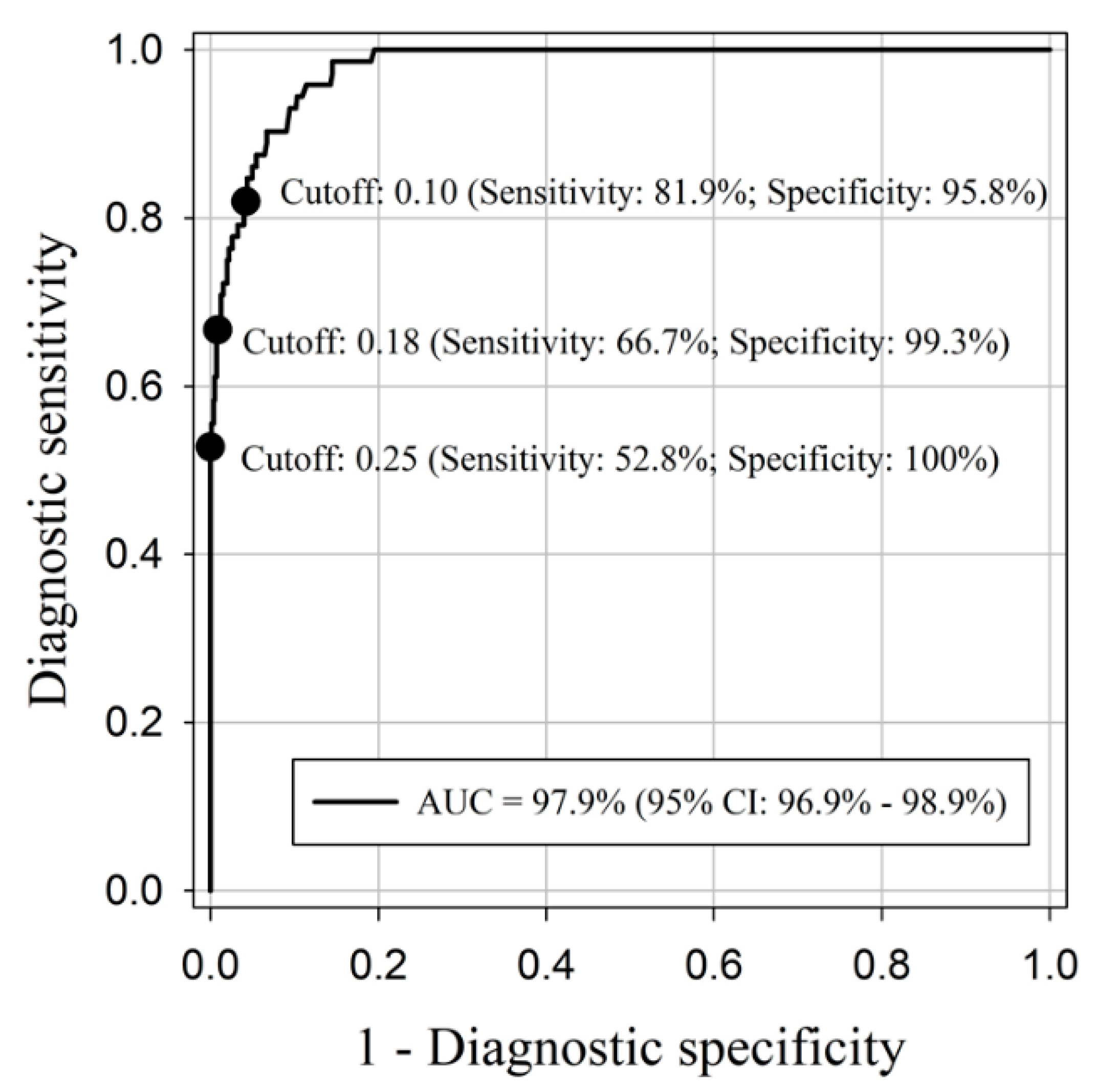
2. Results
3. Discussion
4. Materials and Methods
4.1. Experimental Samples of Known Porcine Coronavirus Infection Status
4.2. Generation of PDCoV S1 Recombinant Protein
4.3. PDCoV Indirect ELISA
4.4. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taxonomy History – Taxonomy—ICTV. Available online: https://talk.ictvonline.org/taxonomy/p/taxonomy-history?taxnode_id=202101870 (accessed on 2 June 2022).
- Woo, P.C.Y.; Lau, S.K.P.; Lam, C.S.F.; Lau, C.C.Y.; Tsang, A.K.L.; Lau, J.H.N.; Bai, R.; Teng, J.L.L.; Tsang, C.C.C.; Wang, M.; et al. Discovery of Seven Novel Mammalian and Avian Coronaviruses in the Genus Deltacoronavirus Supports Bat Coronaviruses as the Gene Source of Alphacoronavirus and Betacoronavirus and Avian Coronaviruses as the Gene Source of Gammacoronavirus and Deltacoronavirus. J. Virol. 2012, 86, 3995–4008. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Huang, Y.; Lau, S.K.P.; Yuen, K.Y. Coronavirus Genomics and Bioinformatics Analysis. Viruses 2010, 2, 1805–1820. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhu, Y.; Wu, M.; Ku, X.; Yao, L.; He, Q. Full-Length Genome Characterization of Chinese Porcine Deltacoronavirus Strain CH/SXD1/2015. Genome Announc. 2015, 3, 1284–1299. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.; Fang, L.; Liu, X.; Hong, Y.; Wang, Y.; Dong, N.; Ma, P.; Bi, J.; Wang, D.; Xiao, S. Identification and Subcellular Localization of Porcine Deltacoronavirus Accessory Protein NS6. Virology 2016, 499, 170. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.; Fang, L.; Hong, Y.; Liu, X.; Dong, N.; Ma, P.; Bi, J.; Wang, D.; Xiao, S. Discovery of a Novel Accessory Protein NS7a Encoded by Porcine Deltacoronavirus. J. Gen. Virol. 2017, 98, 173–178. [Google Scholar] [CrossRef]
- Wang, L.; Byrum, B.; Zhang, Y. Detection and Genetic Characterization of Deltacoronavirus in Pigs, Ohio, USA, 2014. Emerg. Infect. Dis. 2014, 20, 1227. [Google Scholar] [CrossRef]
- Hu, H.; Jung, K.; Vlasova, A.N.; Chepngeno, J.; Lu, Z.; Wang, Q.; Saif, L.J. Isolation and Characterization of Porcine Deltacoronavirus from Pigs with Diarrhea in the United States. J. Clin. Microbiol. 2015, 53, 1537. [Google Scholar] [CrossRef]
- Wang, L.; Byrum, B.; Zhang, Y. Porcine Coronavirus HKU15 Detected in 9 US States, 2014. Emerg. Infect. Dis. 2014, 20, 1594. [Google Scholar] [CrossRef]
- Ajayi, T.; Dara, R.; Misener, M.; Pasma, T.; Moser, L.; Poljak, Z. Herd-level Prevalence and Incidence of Porcine Epidemic Diarrhoea Virus (PEDV) and Porcine Deltacoronavirus (PDCoV) in Swine Herds in Ontario, Canada. Transbound. Emerg. Dis. 2018, 65, 1197. [Google Scholar] [CrossRef]
- Lee, S.; Lee, C. Complete Genome Characterization of Korean Porcine Deltacoronavirus Strain KOR/KNU14-04/2014. Genome Announc. 2014, 2, e01191-14. [Google Scholar] [CrossRef]
- Song, D.; Zhou, X.; Peng, Q.; Chen, Y.; Zhang, F.; Huang, T.; Zhang, T.; Li, A.; Huang, D.; Wu, Q.; et al. Newly Emerged Porcine Deltacoronavirus Associated with Diarrhoea in Swine in China: Identification, Prevalence and Full-Length Genome Sequence Analysis. Transbound. Emerg. Dis. 2015, 62, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Saeng-chuto, K.; Lorsirigool, A.; Temeeyasen, G.; Vui, D.T.; Stott, C.J.; Madapong, A.; Tripipat, T.; Wegner, M.; Intrakamhaeng, M.; Chongcharoen, W.; et al. Different Lineage of Porcine Deltacoronavirus in Thailand, Vietnam and Lao PDR in 2015. Transbound. Emerg. Dis. 2017, 64, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Shibahara, T.; Imai, N.; Yamamoto, T.; Ohashi, S. Genetic Characterization and Pathogenicity of Japanese Porcine Deltacoronavirus. Infect. Genet. Evol. 2018, 61, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rivera, C.; Ramírez-Mendoza, H.; Mendoza-Elvira, S.; Segura-Velázquez, R.; Sánchez-Betancourt, J.I. First Report and Phylogenetic Analysis of Porcine Deltacoronavirus in Mexico. Transbound. Emerg. Dis. 2019, 66, 1436. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Huaman, J.; Gómez-Quispe, O.E. Evaluation of a Porcine Deltacoronavirus Eradication Program in a Full-Cycle Pig Farm in Peru. J. Adv. Vet. Anim. Res. 2021, 8, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Doyle, L.; Hutchings, L. A Transmissible Gastroenteritis in Pigs. J. Am. Vet. Med. Assoc. 1946, 108, 257–259. [Google Scholar]
- Wood, E. An Apparently New Syndrome of Porcine Epidemic Diarrhoea. Vet. Rec. 1977, 100, 243–244. [Google Scholar] [CrossRef]
- Akimkin, V.; Beer, M.; Blome, S.; Hanke, D.; Höper, D.; Jenckel, M.; Pohlmann, A. New Chimeric Porcine Coronavirus in Swine Feces, Germany, 2012. Emerg. Infect. Dis. 2016, 22, 1314. [Google Scholar] [CrossRef]
- Zhou, P.; Fan, H.; Lan, T.; Yang, X.-L.; Shi, W.F.; Zhang, W.; Zhu, Y.; Zhang, Y.W.; Xie, Q.M.; Mani, S.; et al. Fatal Swine Acute Diarrhoea Syndrome Caused by an HKU2-Related Coronavirus of Bat Origin. Nature 2018, 556, 255–259. [Google Scholar] [CrossRef]
- Greig, A.S.; Mitchell, D.; Corner, A.H.; Bannister, G.L.; Meads, E.B.; Julian, R.J. A Hemagglutinating Virus Producing Encephalomyelitis in Baby Pigs. Can. J. Comp. Med. Vet. Sci. 1962, 26, 49–56. [Google Scholar]
- Pensaert, M.; Callebaut, P.; Vergote, J. Isolation of a Porcine Respiratory, Non-Enteric Coronavirus Related to Transmissible Gastroenteritis. Vet. Q. 1986, 8, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Gauger, P.; Stafne, M.; Thomas, J.; Arruda, P.; Burrough, E.; Madson, D.; Brodie, J.; Magstadt, D.; Derscheid, R.; et al. Pathogenicity and Pathogenesis of a United States Porcine Deltacoronavirus Cell Culture Isolate in 5-Day-Old Neonatal Piglets. Virology 2015, 482, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Hu, H.; Eyerly, B.; Lu, Z.; Chepngeno, J.; Saif, L.J. Pathogenicity of 2 Porcine Deltacoronavirus Strains in Gnotobiotic Pigs. Emerg. Infect. Dis. 2015, 21, 650. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, Y.; Liang, X.; Lou, F.; Oglesbee, M.; Krakowka, S.; Li, J. Origin, Evolution, and Virulence of Porcine Deltacoronaviruses in the United States. mBio 2015, 6, e00064-15. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Jung, K.; Vlasova, A.N.; Saif, L.J. Experimental Infection of Gnotobiotic Pigs with the Cell-Culture-Adapted Porcine Deltacoronavirus Strain OH-FD22. Arch. Virol. 2016, 161, 3421. [Google Scholar] [CrossRef]
- Li, G.; Chen, Q.; Harmon, K.M.; Yoon, K.J.; Schwartz, K.J.; Hoogland, M.J.; Gauger, P.C.; Main, R.G.; Zhang, J. Full-Length Genome Sequence of Porcine Deltacoronavirus Strain USA/IA/2014/8734. Genome Announc. 2014, 2, 278–292. [Google Scholar] [CrossRef]
- Janetanakit, T.; Lumyai, M.; Bunpapong, N.; Boonyapisitsopa, S.; Chaiyawong, S.; Nonthabenjawan, N.; Kesdaengsakonwut, S.; Amonsin, A. Porcine Deltacoronavirus, Thailand, 2015. Emerg. Infect. Dis. 2016, 22, 757. [Google Scholar] [CrossRef]
- Wang, L.; Hayes, J.; Sarver, C.; Byrum, B.; Zhang, Y. Porcine Deltacoronavirus: Histological Lesions and Genetic Characterization. Arch. Virol. 2016, 161, 171. [Google Scholar] [CrossRef]
- Zhang, J. Porcine Deltacoronavirus: Overview of Infection Dynamics, Diagnostic Methods, Prevalence and Genetic Evolution. Virus Res. 2016, 226, 71–84. [Google Scholar] [CrossRef]
- Su, M.; Li, C.; Guo, D.; Wei, S.; Wang, X.; Geng, Y.; Yao, S.; Gao, J.; Wang, E.; Zhao, X.; et al. A Recombinant Nucleocapsid Protein-Based Indirect Enzyme-Linked Immunosorbent to Detect Antibodies against Porcine Deltacoronavirus. J. Vet. Med. Sci. 2016, 78, 601. [Google Scholar] [CrossRef]
- Thachil, A.; Gerber, P.F.; Xiao, C.T.; Huang, Y.W.; Opriessnig, T. Development and Application of an ELISA for the Detection of Porcine Deltacoronavirus IgG Antibodies. PLoS ONE 2015, 10, e0124363. [Google Scholar] [CrossRef]
- Okda, F.; Lawson, S.; Liu, X.; Singrey, A.; Clement, T.; Hain, K.; Nelson, J.; Christopher-Hennings, J.; Nelson, E.A. Development of Monoclonal Antibodies and Serological Assays Including Indirect ELISA and Fluorescent Microsphere Immunoassays for Diagnosis of Porcine Deltacoronavirus. BMC Vet. Res. 2016, 12, 95. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.-X.; Fan, J.H.; Opriessnig, T.; Di, J.M.; Liu, B.-J.; Zuo, Y.Z. Development and Application of a Recombinant M Protein-Based Indirect ELISA for the Detection of Porcine Deltacoronavirus IgG Antibodies. J. Virol. Methods 2017, 249, 76. [Google Scholar] [CrossRef] [PubMed]
- Hsu, T.H.; Liu, H.P.; Chin, C.Y.; Wang, C.; Zhu, W.Z.; Wu, B.L.; Chang, Y.C. Detection, Sequence Analysis, and Antibody Prevalence of Porcine Deltacoronavirus in Taiwan. Arch. Virol. 2018, 163, 3113–3117. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Liu, Q.; Wang, X.; Zhang, J.; Zhang, X.; Shi, D.; Liu, J.; Shi, H.; Chen, J.; Feng, L. Development of an Indirect ELISA for Detecting Porcine Deltacoronavirus IgA Antibodies. Arch. Virol. 2020, 165, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Zhang, Y.; Hu, H. ELISA, Virus Neutralization, and Immunofluorescence Assays for Detection of Antibodies Against Porcine Deltacoronavirus. In Animal Coronaviruses; Humana: New York, NY, USA, 2022; pp. 197–207. [Google Scholar]
- Ma, Y.; Zhang, Y.; Liang, X.; Oglesbee, M.; Krakowka, S.; Niehaus, A.; Wang, G.; Jia, A.; Song, H.; Li, J. Two-Way Antigenic Cross-Reactivity between Porcine Epidemic Diarrhea Virus and Porcine Deltacoronavirus. Vet. Microbiol. 2016, 186, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Gimenez-Lirola, L.G.; Zhang, J.; Carrillo-Avila, J.A.; Chen, Q.; Magtoto, R.; Poonsuk, K.; Baum, D.H.; Piñeyro, P.; Zimmerman, J. Reactivity of Porcine Epidemic Diarrhea Virus Structural Proteins to Antibodies against Porcine Enteric Coronaviruses: Diagnostic Implications. J. Clin. Microbiol. 2017, 55, 1426. [Google Scholar] [CrossRef]
- Magtoto, R.; Poonsuk, K.; Baum, D.; Zhang, J.; Chen, Q.; Ji, J.; Piñeyro, P.; Zimmerman, J.; Giménez-Lirola, L.G. Evaluation of the Serologic Cross-Reactivity between Transmissible Gastroenteritis Coronavirus and Porcine Respiratory Coronavirus Using Commercial Blocking Enzyme-Linked Immunosorbent Assay Kits. mSphere 2019, 4, e00017-19. [Google Scholar] [CrossRef] [PubMed]
- Vitosh-Sillman, S.; Loy, J.D.; Brodersen, B.; Kelling, C.; Doster, A.; Topliff, C.; Nelson, E.; Bai, J.; Schirtzinger, E.; Poulsen, E.; et al. Experimental Infection of Conventional Nursing Pigs and Their Dams with Porcine Deltacoronavirus. J. Vet. Diagn. Investig. 2016, 28, 486–497. [Google Scholar] [CrossRef] [PubMed]
- Bosch, B.J.; van der Zee, R.; de Haan, C.A.M.; Rottier, P.J.M. The Coronavirus Spike Protein Is a Class I Virus Fusion Protein: Structural and Functional Characterization of the Fusion Core Complex. J. Virol. 2003, 77, 8801–8811. [Google Scholar] [CrossRef]
- Li, W.; Hulswit, R.J.G.; Kenney, S.P.; Widjaja, I.; Jung, K.; Alhamo, M.A.; van Dieren, B.; van Kuppeveld, F.J.M.; Saif, L.J.; Bosch, B.J. Broad Receptor Engagement of an Emerging Global Coronavirus May Potentiate Its Diverse Cross-Species Transmissibility. Proc. Natl. Acad. Sci. USA 2018, 115, E5135–E5143. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, S.; Wang, X.; Luo, Z.; Shi, Y.; Wang, D.; Peng, G.; Chen, H.; Fang, L.; Xiao, S. Contribution of Porcine Aminopeptidase N to Porcine Deltacoronavirus Infection. Emerg. Microbes Infect. 2018, 7, 65. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, A.N.; Kenney, S.P.; Jung, K.; Wang, Q.; Saif, L.J. Deltacoronavirus Evolution and Transmission: Current Scenario and Evolutionary Perspectives. Front. Vet. Sci. 2021, 7, 1257. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Fu, J.; Hu, J.; Li, C.; Zhao, Y.; Qu, H.; Wen, X.; Cao, S.; Wen, Y.; Wu, R.; et al. Identification of the Immunodominant Neutralizing Regions in the Spike Glycoprotein of Porcine Deltacoronavirus. Virus Res. 2020, 276, 197834. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, B.; Liang, Q.-Z.; Shi, F.-S.; Ji, C.-M.; Yang, X.-L.; Yang, Y.-L.; Qin, P.; Chen, R.; Huang, Y.-W. Roles of Two Major Domains of the Porcine Deltacoronavirus S1 Subunit in Receptor Binding and Neutralization. J. Virol. 2021, 95, e01118-21. [Google Scholar] [CrossRef]
- Boley, P.A.; Alhamo, M.A.; Lossie, G.; Yadav, K.K.; Vasquez-Lee, M.; Saif, L.J.; Kenney, S.P. Porcine Deltacoronavirus Infection and Transmission in Poultry, United States. Emerg. Infect. Dis. 2020, 26, 255. [Google Scholar] [CrossRef]
- Jung, K.; Hu, H.; Saif, L.J. Calves Are Susceptible to Infection with the Newly Emerged Porcine Deltacoronavirus, but Not with the Swine Enteric Alphacoronavirus, Porcine Epidemic Diarrhea Virus. Arch. Virol. 2017, 162, 2357. [Google Scholar] [CrossRef]
- Lednicky, J.A.; Tagliamonte, M.S.; White, S.K.; Elbadry, M.A.; Alam, M.M.; Stephenson, C.J.; Bonny, T.S.; Loeb, J.C.; Telisma, T.; Chavannes, S.; et al. Independent Infections of Porcine Deltacoronavirus among Haitian Children. Nature 2021, 600, 133–137. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, I.J.; Pyo, H.M.; Tark, D.S.; Song, J.Y.; Hyun, B.H. Multiplex Real-Time RT-PCR for the Simultaneous Detection and Quantification of Transmissible Gastroenteritis Virus and Porcine Epidemic Diarrhea Virus. J. Virol. Methods 2007, 146, 172–177. [Google Scholar] [CrossRef]
- Madson, D.M.; Magstadt, D.R.; Arruda, P.H.E.; Hoang, H.; Sun, D.; Bower, L.P.; Bhandari, M.; Burrough, E.R.; Gauger, P.C.; Pillatzki, A.E.; et al. Pathogenesis of Porcine Epidemic Diarrhea Virus Isolate (US/Iowa/18984/2013) in 3-Week-Old Weaned Pigs. Vet. Microbiol. 2014, 174, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Mora-Díaz, J.C.; Temeeyasen, G.; Magtoto, R.; Rauh, R.; Nelson, W.; Carrillo-Ávila, J.A.; Zimmerman, J.; Piñeyro, P.; Giménez-Lirola, L. Infection and Immune Response to Porcine Hemagglutinating Encephalomyelitis Virus in Grower Pigs. Vet. Microbiol. 2021, 253, 108958. [Google Scholar] [CrossRef]
- Mora-Díaz, J.C.; Magtoto, R.; Houston, E.; Baum, D.; Carrillo-Ávila, J.A.; Temeeyasen, G.; Zimmerman, J.; Piñeyro, P.; Giménez-Lirola, L. Detecting and Monitoring Porcine Hemagglutinating Encephalomyelitis Virus, an Underresearched Betacoronavirus. mSphere 2020, 5, e00199-20. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Li, G.; Stasko, J.; Thomas, J.T.; Stensland, W.R.; Pillatzki, A.E.; Gauger, P.C.; Schwartz, K.J.; Madson, D.; Yoon, K.J.; et al. Isolation and Characterization of Porcine Epidemic Diarrhea Viruses Associated with the 2013 Disease Outbreak among Swine in the United States. J. Clin. Microbiol. 2014, 52, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.C.; Müller, M. PROC: An Open-Source Package for R and S+ to Analyze and Compare ROC Curves. BMC Bioinformatics 2011, 12, 77. [Google Scholar] [CrossRef] [PubMed]
- DeLong, E.R.; DeLong, D.M.; Clarke-Pearson, D.L. Comparing the Areas under Two or More Correlated Receiver Operating Characteristic Curves: A Nonparametric Approach. Biometrics 1988, 44, 837–845. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pig ID | Cutoff | Day Post-Inoculation | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 7 | 10 | 14 | 17 | 21 | 28 | 35 | 42 | ||
| 1 | 0.25 | Neg a | Neg | Pos | Neg | Pos | Pos | Pos | Pos |
| 0.18 | Neg | Neg | Pos | Pos | Pos | Pos | Pos | Pos | |
| 0.10 | Neg | Pos b | Pos | Pos | Pos | Pos | Pos | Pos | |
| 2 | 0.25 | Neg | Neg | Pos | Pos | Pos | Pos | Pos | Pos |
| 0.18 | Neg | Pos | Pos | Pos | Pos | Pos | Pos | Pos | |
| 0.10 | Neg | Pos | Pos | Pos | Pos | Pos | Pos | Pos | |
| 3 | 0.25 | Neg | Pos | Neg | Pos | Pos | Pos | Neg | Pos |
| 0.18 | Neg | Pos | Neg | Pos | Pos | Pos | Neg | Pos | |
| 0.10 | Pos | Pos | Pos | Pos | Pos | Pos | Neg | Pos | |
| 4 | 0.25 | Neg | Neg | Pos | Pos | Pos | Pos | Neg | Pos |
| 0.18 | Neg | Neg | Pos | Pos | Pos | Pos | Pos | Pos | |
| 0.10 | Pos | Neg | Pos | Pos | Pos | Pos | Pos | Pos | |
| 5 | 0.25 | Neg | Neg | Neg | Pos | Neg | Neg | Pos | Neg |
| 0.18 | Neg | Neg | Neg | Pos | Neg | Neg | Pos | Pos | |
| 0.10 | Neg | Neg | Neg | Pos | Neg | Neg | Pos | Pos | |
| 6 | 0.25 | Neg | Neg | Neg | Neg | Pos | Pos | Neg | Pos |
| 0.18 | Neg | Neg | Neg | Pos | Pos | Pos | Neg | Pos | |
| 0.10 | Neg | Neg | Pos | Pos | Pos | Pos | Neg | Pos | |
| 7 | 0.25 | Neg | Neg | Pos | Pos | Pos | Pos | Neg | Pos |
| 0.18 | Neg | Neg | Pos | Pos | Pos | Pos | Neg | Pos | |
| 0.10 | Neg | Neg | Pos | Pos | Pos | Pos | Neg | Pos | |
| 8 | 0.25 | Neg | Neg | Neg | Neg | Pos | Neg | Pos | Neg |
| 0.18 | Neg | Neg | Pos | Neg | Pos | Neg | Pos | Neg | |
| 0.10 | Neg | Neg | Pos | Neg | Pos | Pos | Pos | Pos | |
| 9 | 0.25 | Neg | Neg | Neg | Neg | Neg | Neg | Neg | Neg |
| 0.18 | Neg | Neg | Neg | Pos | Neg | Neg | Neg | Neg | |
| 0.10 | Neg | Neg | Neg | Pos | Neg | Neg | Pos | Pos | |
| 10 | 0.25 | Neg | Neg | Neg | Pos | Neg | Neg | Pos | Neg |
| 0.18 | Neg | Neg | Pos | Pos | Neg | Pos | Pos | Neg | |
| 0.10 | Neg | Pos | Pos | Pos | Pos | Pos | Pos | Pos | |
| 11 | 0.25 | Neg | Neg | Neg | Neg | Neg | Pos | Pos | Pos |
| 0.18 | Neg | Neg | Neg | Neg | Neg | Pos | Pos | Pos | |
| 0.10 | Neg | Neg | Neg | Pos | Pos | Pos | Pos | Pos | |
| 12 | 0.25 | Neg | Neg | Neg | Neg | Neg | Neg | Pos | Neg |
| 0.18 | Neg | Neg | Neg | Neg | Neg | Pos | Pos | Pos | |
| 0.10 | Pos | Neg | Neg | Pos | Neg | Pos | Pos | Pos | |
| Total | 0.25 | 0/12 (0%) | 1/12 (8.3%) | 4/12 (33.3%) | 6/12 (50%) | 7/12 (58.3%) | 7/12 (58.3%) | 7/12 (58.3%) | 7/12 (58.3%) |
| 0.18 | 0/12 (0%) | 2/12 (16.7%) | 6/12 (50%) | 9/12 (75%) | 7/12 (58.3%) | 9/12 (75%) | 8/12 (66.7%) | 9/12 (75%) | |
| 0.10 | 3/12 (25%) | 4/12 (33.3%) | 8/12 (66.7%) | 11/12 (91.7%) | 9/12 (75%) | 10/12 (83.3%) | 9/12 (75%) | 12/12 (100%) | |
| Virus Strain | Cell Line | Virus Titer (TCID50/mL) | Virus Inoculum (mL) | Inoculation Route | Reference | |
|---|---|---|---|---|---|---|
| PDCoV (12) | USA/IL/2014 | Swine testicle (ATCC® CRL-1746) | 1.5 × 106 | 30 | Orogastric | [23] |
| TGEV Miller (12) | ATCC a VR-1740 | 4.0 × 106 | 35 | Orogastric | [39,55] | |
| TGEV Purdue (12) | ATCC VR-763 | 2.4 × 108 | 30 | Orogastric | ||
| PRCV (12) | ATCC VR-2384 | 4.0 × 105 | 15 | Nasal | ||
| PEDV (12) | USA/IN/2013/19338E | Vero (ATCC® CCL-81) | 1.5 × 106 | 15 | Orogastric | |
| PHEV (12) | NVSL PHEV 67N | Swine kidney primary cells (NVSL-USDA b) | 1:128 c | 5 | Oronasal | [53] |
| Negative control (12) | Culture medium | - | - | 20 | Oronasal | - |
| Sequences | |
|---|---|
| DNA | ATGACATCCACTTTGCCTTTCTCTCCACAGGTGTCCACTCCCAGGTCCAAGTTTAAACGGATCTCTAGCGAATTCGCCGCCACCATGCAGAGAGCACTGCTGATTATGACTCTGCTGTGTCTGGTCAGAGCTAAGTTCGCTGATGATCTGCTGGACCTGCTGACATTCCCTGGAGCTCATAGATTCCTGCATAAGCCTACCAGGAACAGCAGCTCCCTGTATTCCAGGGCTAACAACAACTTCGATGTGGGAGTGCTGCCTGGATACCCTACCAAGAACGTCAACCTGTTTAGCCCTCTGACAAATTCCACCCTGCCCATCAACGGACTGCACAGAAGCTACCAGCCTCTGATGCTGAATTGCCTGACTAAGATTACCAACCACACCCTGAGCATGTACCTGCTGCCCTCCGAAATCCAGACCTACAGCTGCGGAGGCGCCATGGTCAAATACCAAACTCATGATGCAGTGAGGATCATCCTGGATCTGACTGCCACAGACCACATCTCCGTCGAAGTGGTCGGCCAGCACGGAGAGAACTACGTGTTTGTGTGTAGCGAGCAGTTTAACTACACCACCGCCCTGCACAATAGCACATTCTTCAGCCTGAACTCCGAACTGTACTGCTTCACCAACAACACATACCTGGGCATCCTGCCACCCGACCTGACCGACTTCACTGTCTACAGGACCGGCCAGTTCTACGCCAATGGCTATCTGCTGGGAACACTGCCTATTACCGTGAACTATGTGAGACTGTATAGAGGCCACCTGAGCGCCAACAGCGCCCACTTTGCTCTGGCCAATCTGACAGATACTCTGATCACACTGACCAACACAACTATCAGCCAGATTACATACTGCGACAAGAGCGTGGTGGACAGCATCGCCTGCCAGAGAAGCAGCCACGAGGTGGAGGACGGCTTCTACTCCGATCCCAAATCCGCCGTCAGGGCAAGACAAAGGACTATCGTCACTCTGCCCAAGCTGCCCGAGCTGGAGGTCGTGCAGCTGAACATTTCCGCCCACATGGACTTCGGAGAAGCCAGGCTGGATAGCGTGACCATCAATGGCAACACCAGCTATTGCGTGACAAAGCCTTACTTCAGACTGGAGACAAACTTCATGTGCACCGGCTGCACCATGAACCTGAGGACCGACACCTGCAGCTTCGATCTGTCCGCTGTCAACAACGGGATGTCCTTCTCCCAATTCTGTCTGAGCACCGAGTCCGGAGCATGCGAGATGAAGATCATTGTGACCTACGTCTGGAATTACCTGCTGAGGCAGAGGCTGTATGTCACTGCCGTGGAAGGCCAAACCCACACCGGAACCACCTCCGTGCATGCCACTGACACTAGCTCCGTCATCACTGATGTGTGCACTGATTACACCATCTACGGCGTGAGCGGCACCGGGATCATTAAGCCCAGCGATCTGCTGCTGCACAACGGCATCGCTTTCACCTCTCCCACCGGCGAGCTGTACGCCTTCAAGAATATCACTACCGGCAAGACCCTGCAAGTCCTGCCTTGCGAGACACCCAGCCAGCTGATTGTCATCAACAATACCGTCGTGGGAGCAATCACAAGCTCCAACTCCACCGAGAACAATAGGTTCACCACAACAATCGTGACACCAACCTTCTTCTACGAGAACCTGTACTTCCAGAGCGGCTCCGACAAGACCCACACCGTCGAGTGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGAGGAGATGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTATAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAATGA |
| Primers | SPDCV-F-F: 5′-TAAACGGATCTCTAGCGAATTCGCCGCCACCATGCAGAGAGC-3′ SPDCV-F-R1:5′-GTCTTGTCGGAGCCGCTCTGGAAGTACAGGTTCTCGTAGAAGAAGGTTGGTGTCAC-3′ SPEDV-F1: 5′-CTTCCAGAGCGGCTCCGACAAGACCCACACCGTCGAGTGCCCACCGTGCCCAG-3′ SPEDV-R: 5′-CGAGCGGCCGCTAGCAAGCTTTCATTTACCCGGAGACAGGGAG-3′ |
| Amino acid | MQRALLIMTLLCLVRAKFADDLLDLLTFPGAHRFLHKPTRNSSSLYSRANNNFDVGVLPGYPTKNVNLFSPLTNSTLPINGLHRSYQPLMLNCLTKITNHTLSMYLLPSEIQTYSCGGAMVKYQTHDAVRIILDLTATDHISVEVVGQHGENYVFVCSEQFNYTTALHNSTFFSLNSELYCFTNNTYLGILPPDLTDFTVYRTGQFYANGYLLGTLPITVNYVRLYRGHLSANSAHFALANLTDTLITLTNTTISQITYCDKSVVDSIACQRSSHEVEDGFYSDPKSAVRARQRTIVTLPKLPELEVVQLNISAHMDFGEARLDSVTINGNTSYCVTKPYFRLETNFMCTGCTMNLRTDTCSFDLSAVNNGMSFSQFCLSTESGACEMKIIVTYVWNYLLRQRLYVTAVEGQTHTGTTSVHATDTSSVITDVCTDYTIYGVSGTGIIKPSDLLLHNGIAFTSPTGELYAFKNITTGKTLQVLPCETPSQLIVINNTVVGAITSSNSTENNRFTTTIVTPTFFYENLYFQSGSDKTHTVECPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yen, L.; Magtoto, R.; Mora-Díaz, J.C.; Carrillo-Ávila, J.A.; Zhang, J.; Cheng, T.-Y.; Magtoto, P.; Nelli, R.K.; Baum, D.H.; Zimmerman, J.J.; et al. The N-terminal Subunit of the Porcine Deltacoronavirus Spike Recombinant Protein (S1) Does Not Serologically Cross-react with Other Porcine Coronaviruses. Pathogens 2022, 11, 910. https://doi.org/10.3390/pathogens11080910
Yen L, Magtoto R, Mora-Díaz JC, Carrillo-Ávila JA, Zhang J, Cheng T-Y, Magtoto P, Nelli RK, Baum DH, Zimmerman JJ, et al. The N-terminal Subunit of the Porcine Deltacoronavirus Spike Recombinant Protein (S1) Does Not Serologically Cross-react with Other Porcine Coronaviruses. Pathogens. 2022; 11(8):910. https://doi.org/10.3390/pathogens11080910
Chicago/Turabian StyleYen, Lu, Ronaldo Magtoto, Juan Carlos Mora-Díaz, Jose Antonio Carrillo-Ávila, Jianqiang Zhang, Ting-Yu Cheng, Precy Magtoto, Rahul K. Nelli, David H. Baum, Jeffrey J. Zimmerman, and et al. 2022. "The N-terminal Subunit of the Porcine Deltacoronavirus Spike Recombinant Protein (S1) Does Not Serologically Cross-react with Other Porcine Coronaviruses" Pathogens 11, no. 8: 910. https://doi.org/10.3390/pathogens11080910