
Molecular Insights into Innate Immune Response in Captive Koala Peripheral Blood Mononuclear Cells Co-Infected with Multiple Koala Retrovirus Subtypes

 , ,
, ,
Abstract
:1. Introduction
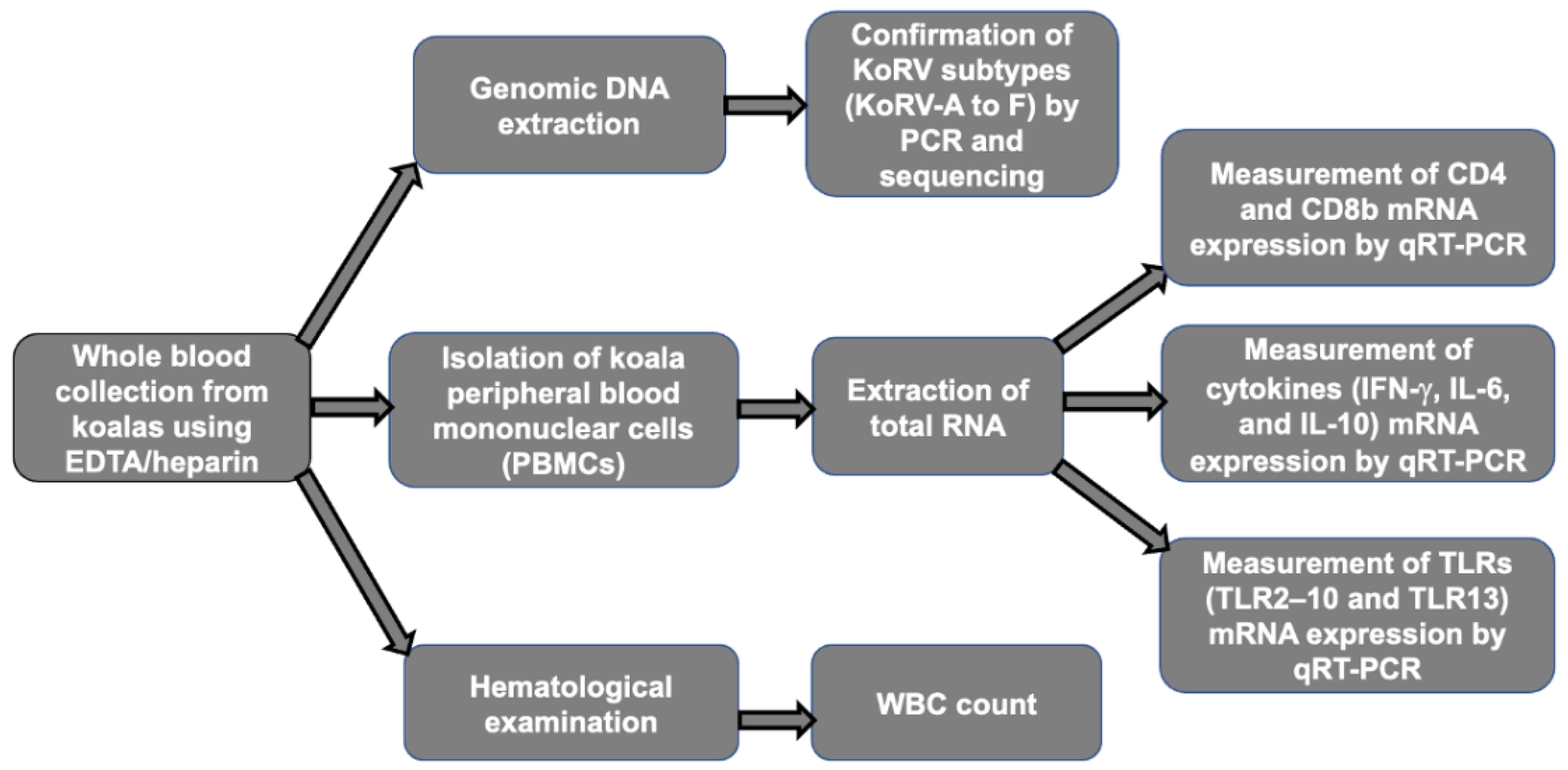
2. Materials and Methods
2.1. Animals
2.2. Hematological Examination and Clinical Sign Observation
2.3. Extraction of Genomic DNA
2.4. Extraction of RNA from Koala Peripheral Blood Mononuclear Cells (PBMCs)
2.5. Gene Expression Analysis by Quantitative Reverse Transcription-PCR (qRT-PCR)
2.6. Statistical Analysis
3. Results
3.1. Infection Status of Koalas for KoRV Subtypes
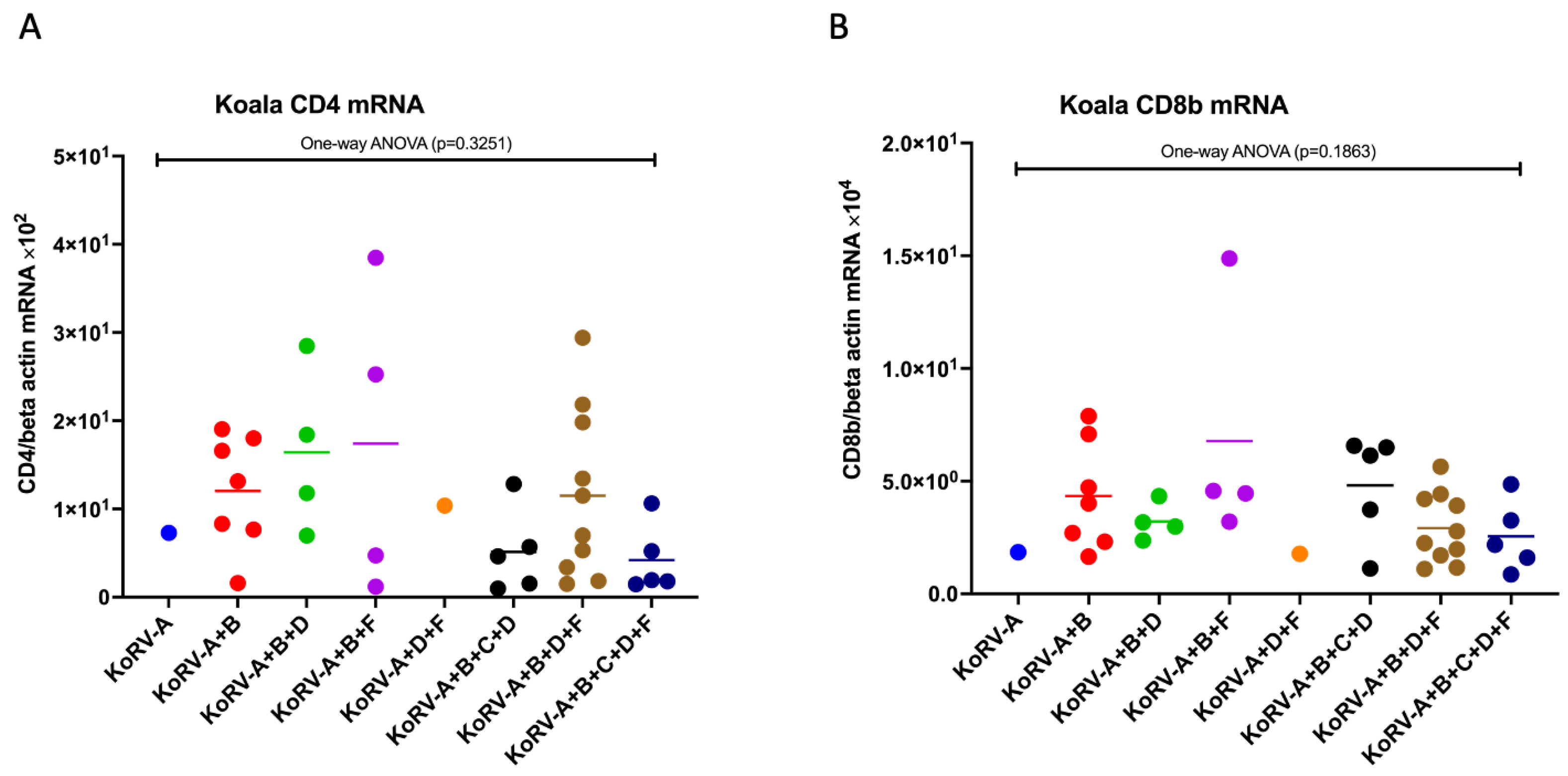
3.2. Expression Patterns of CD4 and CD8b mRNA in Koala PBMCs
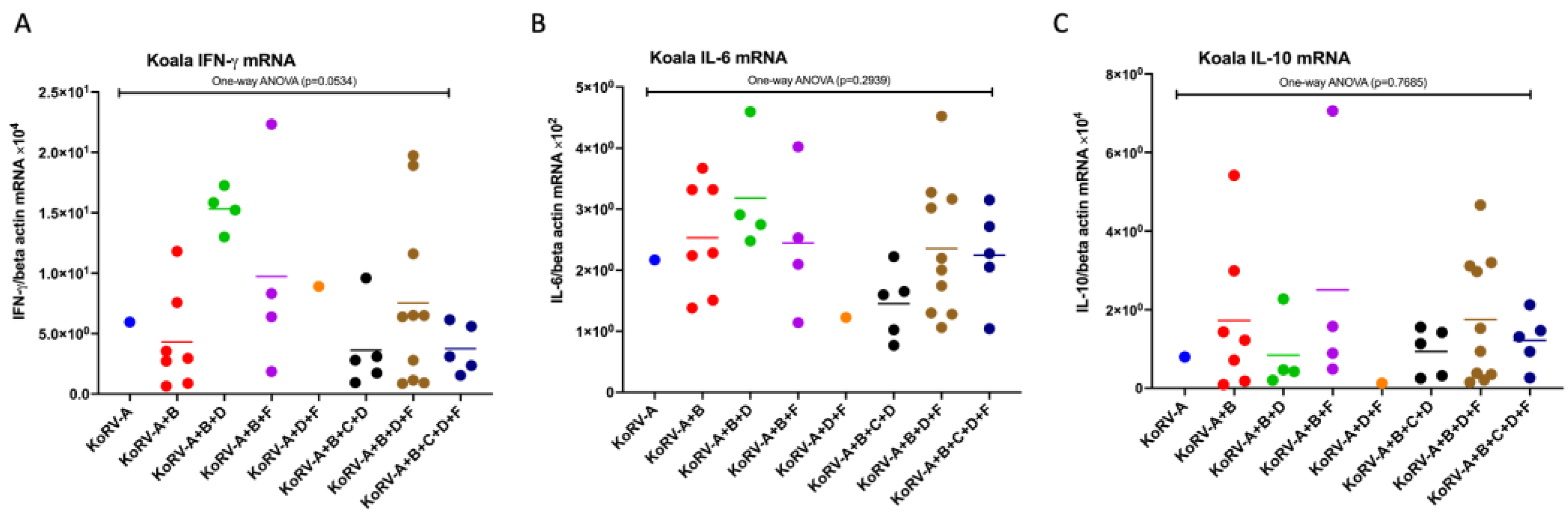
3.3. Expression Patterns of Cytokine mRNAs in Koala PBMCs
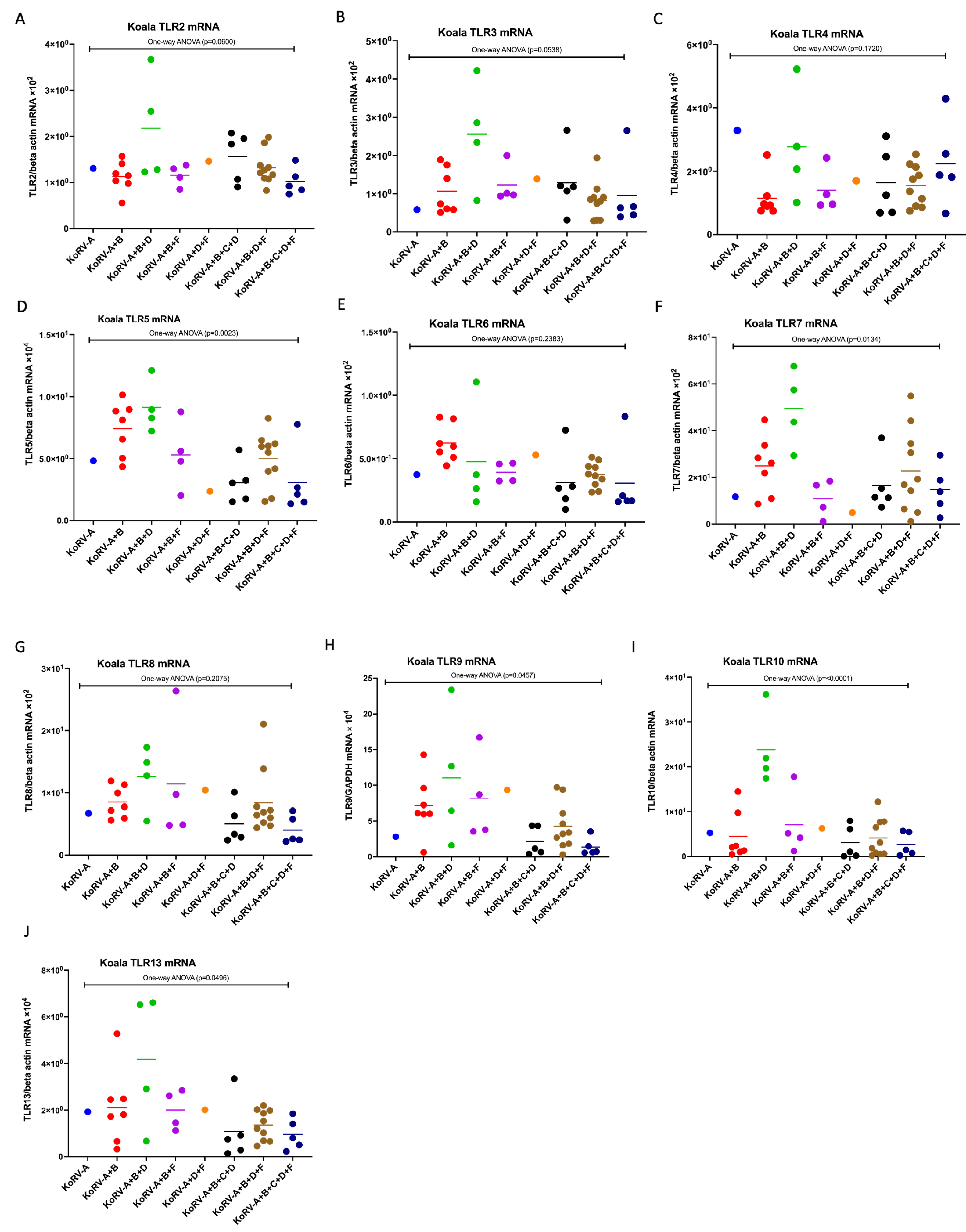
3.4. Expression Patterns of TLR mRNAs in Koala PBMCs
3.5. Difference in the Expression Patterns of CD4, CD8b, Cytokines, and TLR mRNAs between the Groups in Koala PBMCs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanger, J.J.; Bromham, L.D.; McKee, J.J.; O’Brien, T.M.; Robinson, W.F. The nucleotide sequence of koala (Phascolarctos cinereus) retrovirus: A novel type C endogenous virus related to Gibbon ape leukemia virus. J. Virol. 2000, 74, 4264–4272. [Google Scholar] [CrossRef] [PubMed]
- Tarlinton, R.; Meers, J.; Hanger, J.; Young, P. Real-time reverse transcriptase PCR for the endogenous koala retrovirus reveals an association between plasma viral load and neoplastic disease in koalas. J. Gen. Virol. 2005, 86, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Tarlinton, R.E.; Meers, J.; Young, P.R. Retroviral invasion of the koala genome. Nature 2006, 442, 79–81. [Google Scholar] [CrossRef] [PubMed]
- Quigley, B.L.; Timms, P. Helping koalas battle disease—Recent advances in Chlamydia and koala retrovirus (KoRV) disease understanding and treatment in koalas. FEMS Microbiol. Rev. 2020, 44, 583–605. [Google Scholar] [CrossRef] [PubMed]
- Melzer, A.; Carrick, F.; Menkhorst, P.; Lunney, D.; John, B.S. Overview, critical assessment, and conservation implications of koala distribution and abundance. Conserv. Biol. 2000, 14, 619–628. [Google Scholar] [CrossRef]
- Woinarski, J.; Burbidge, A.A. Phascolarctos Cinereus (Amended Version of 2016 Assessment). The IUCN Red List of Threatened Species 2020: E.T16892A166496779. 2020. Available online: https://www.iucnredlist.org/species/16892/166496779 (accessed on 30 September 2021).
- Kayesh, M.E.H.; Hashem, M.A.; Tsukiyama-Kohara, K. Koala retrovirus epidemiology, transmission mode, pathogenesis, and host immune response in koalas (Phascolarctos cinereus): A review. Arch. Virol. 2020, 165, 2409–2417. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Stadler, C.K.; Gorman, K.; Jensen, N.; Kim, D.; Zheng, H.; Tang, S.; Switzer, W.M.; Pye, G.W.; Eiden, M.V. An exogenous retrovirus isolated from koalas with malignant neoplasias in a US zoo. Proc. Natl. Acad. Sci. USA 2013, 110, 11547–11552. [Google Scholar] [CrossRef]
- Hashem, M.A.; Maetani, F.; Kayesh, M.E.H.; Eiei, T.; Mochizuki, K.; Ito, A.; Sakurai, H.; Asai, T.; Tsukiyama-Kohara, K. Transmission of Koala Retrovirus from Parent Koalas to a Joey in a Japanese Zoo. J. Virol. 2020, 94, e00019-20. [Google Scholar] [CrossRef]
- Kayesh, M.E.H.; Hashem, M.A.; Tsukiyama-Kohara, K. Toll-Like Receptor and Cytokine Responses to Infection with Endogenous and Exogenous Koala Retrovirus, and Vaccination as a Control Strategy. Curr. Issues Mol. Biol. 2021, 43, 52–64. [Google Scholar] [CrossRef]
- Blyton, M.D.J.; Pyne, M.; Young, P.; Chappell, K. Koala retrovirus load and non-A subtypes are associated with secondary disease among wild northern koalas. PLoS Pathog. 2022, 18, e1010513. [Google Scholar] [CrossRef]
- Joyce, B.A.; Blyton, M.D.J.; Johnston, S.D.; Young, P.R.; Chappell, K.J. Koala retrovirus genetic diversity and transmission dynamics within captive koala populations. Proc. Natl. Acad. Sci. USA 2021, 118, e2024021118. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, M.; King, A.; Salinas, R.; Chen, Z.; Tsangaras, K.; Greenwood, A.D.; Johnson, R.N.; Belov, K.; Wilkins, M.R.; Timms, P. Long-read genome sequence assembly provides insight into ongoing retroviral invasion of the koala germline. Sci. Rep. 2017, 7, 15838. [Google Scholar] [CrossRef] [PubMed]
- Lober, U.; Hobbs, M.; Dayaram, A.; Tsangaras, K.; Jones, K.; Alquezar-Planas, D.E.; Ishida, Y.; Meers, J.; Mayer, J.; Quedenau, C.; et al. Degradation and remobilization of endogenous retroviruses by recombination during the earliest stages of a germ-line invasion. Proc. Natl. Acad. Sci. USA 2018, 115, 8609–8614. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.N.; O’Meally, D.; Chen, Z.; Etherington, G.J.; Ho, S.Y.W.; Nash, W.J.; Grueber, C.E.; Cheng, Y.; Whittington, C.M.; Dennison, S.; et al. Adaptation and conservation insights from the koala genome. Nat. Genet. 2018, 50, 1102–1111. [Google Scholar] [CrossRef] [PubMed]
- Chiu, E.S.; VandeWoude, S. Endogenous Retroviruses Drive Resistance and Promotion of Exogenous Retroviral Homologs. Annu. Rev. Anim. Biosci. 2021, 9, 225–248. [Google Scholar] [CrossRef]
- Stoye, J.P. Koala retrovirus: A genome invasion in real time. Genome Biol. 2006, 7, 241. [Google Scholar] [CrossRef]
- Tarlinton, R.; Meers, J.; Young, P. Biology and evolution of the endogenous koala retrovirus. Cell Mol. Life Sci. 2008, 65, 3413–3421. [Google Scholar] [CrossRef]
- Denner, J.; Young, P.R. Koala retroviruses: Characterization and impact on the life of koalas. Retrovirology 2013, 10, 108. [Google Scholar] [CrossRef]
- Ellmeier, W.; Haust, L.; Tschismarov, R. Transcriptional control of CD4 and CD8 coreceptor expression during T cell development. Cell Mol. Life Sci. 2013, 70, 4537–4553. [Google Scholar] [CrossRef]
- Li, Q.J.; Dinner, A.R.; Qi, S.; Irvine, D.J.; Huppa, J.B.; Davis, M.M.; Chakraborty, A.K. CD4 enhances T cell sensitivity to antigen by coordinating Lck accumulation at the immunological synapse. Nat. Immunol. 2004, 5, 791–799. [Google Scholar] [CrossRef]
- Lyons, G.E.; Moore, T.; Brasic, N.; Li, M.; Roszkowski, J.J.; Nishimura, M.I. Influence of human CD8 on antigen recognition by T-cell receptor-transduced cells. Cancer Res. 2006, 66, 11455–11461. [Google Scholar] [CrossRef] [PubMed]
- Rustagi, A.; Gale, M., Jr. Innate antiviral immune signaling, viral evasion and modulation by HIV-1. J. Mol. Biol. 2014, 426, 1161–1177. [Google Scholar] [CrossRef] [PubMed]
- Zuniga, E.I.; Macal, M.; Lewis, G.M.; Harker, J.A. Innate and Adaptive Immune Regulation during Chronic Viral Infections. Annu. Rev. Virol. 2015, 2, 573–597. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like Receptors and the Control of Immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity 2011, 34, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef]
- Cui, J.; Frankham, G.J.; Johnson, R.N.; Polkinghorne, A.; Timms, P.; O’Meally, D.; Cheng, Y.; Belov, K. SNP marker discovery in koala TLR genes. PLoS ONE 2015, 10, e0121068. [Google Scholar] [CrossRef]
- Kayesh, M.E.H.; Hashem, M.A.; Maetani, F.; Eiei, T.; Mochizuki, K.; Ochiai, S.; Ito, A.; Ito, N.; Sakurai, H.; Asai, T.; et al. CD4, CD8b, and Cytokines Expression Profiles in Peripheral Blood Mononuclear Cells Infected with Different Subtypes of KoRV from Koalas (Phascolarctos cinereus) in a Japanese Zoo. Viruses 2020, 12, 1415. [Google Scholar] [CrossRef]
- Kayesh, M.E.H.; Hashem, M.A.; Tsukiyama-Kohara, K. Toll-Like Receptor Expression Profiles in Koala (Phascolarctos cinereus) Peripheral Blood Mononuclear Cells Infected with Multiple KoRV Subtypes. Animals 2021, 11, 983. [Google Scholar] [CrossRef]
- Hashem, M.A.; Kayesh, M.E.H.; Maetani, F.; Goto, A.; Nagata, N.; Kasori, A.; Imanishi, T.; Tsukiyama-Kohara, K. Subtype distribution and expression of the koala retrovirus in the Japanese zoo koala population. Infect Genet. Evol. 2022, 102, 105297. [Google Scholar] [CrossRef]
- Miyazawa, T.; Shojima, T.; Yoshikawa, R.; Ohata, T. Isolation of koala retroviruses from koalas in Japan. J. Vet. Med. Sci. 2011, 73, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Kayesh, M.E.H.; Yamato, O.; Rahman, M.M.; Hashem, M.A.; Maetani, F.; Eiei, T.; Mochizuki, K.; Sakurai, H.; Tsukiyama-Kohara, K. Molecular dynamics of koala retrovirus infection in captive koalas in Japan. Arch. Virol. 2019, 164, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Maher, I.E.; Griffith, J.E.; Lau, Q.; Reeves, T.; Higgins, D.P. Expression profiles of the immune genes CD4, CD8beta, IFNgamma, IL-4, IL-6 and IL-10 in mitogen-stimulated koala lymphocytes (Phascolarctos cinereus) by qRT-PCR. PeerJ 2014, 2, e280. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Koala Gene | Forward (5′ to 3′) | Reverse (5′ to 3′) | Product Length (bp) | Reference |
|---|---|---|---|---|
| TLR2 | CCATTCCAAGTGAGGGGCAA | ACTCCAGTCAGCAAGGCAAG | 122 | [30] |
| TLR3 | GGAATGGCTTGGGTTGGAGT | AGCCACTGGAAAGAAAAATCATCT | 162 | [30] |
| TLR4 | TCCACAAGAGCCGGAAAGTC | GAGTTCCACCTGTTGCCGTA | 176 | [30] |
| TLR5 | CCTTAGCCTGGATGGCAACA | GGTAGGGTCAGGGGATAGCA | 109 | [30] |
| TLR6 | TTCAGTTTCCCGCCCAACTA | ATGTGGCCATCCACTTACCA | 157 | [30] |
| TLR7 | TTGCCTTGTAACGTCACCCA | GTGAGGGTCAGGTTGGTTGT | 119 | [30] |
| TLR8 | CCTCTTCGTTTACCACCCTCC | CTTCAAAGGCCCCGTCATCT | 178 | [30] |
| TLR9 | ATCTTCAGCCACTTCCGCTC | AGGCTCTCTCCAGCCCTAAA | 133 | [30] |
| TLR10 | GCCCTAAAGGTGGAGCATGT | TATATGTGGCATCCCCGCAC | 123 | [30] |
| TLR13 | AGCCTACTGGTGGCTATGGA | TGGCCAGGTACAGGGACTTA | 172 | [30] |
| CD4 | GCCAACCCAAGTGACTCTGT | TCTCCTGGACCACTCCATTC | 105 | [34] |
| CD8b | GCATTGGCTTCTAATTGCTAGTATC | CACTTTCTATCATGCAAAGTAACCC | 88 | [29] |
| IFN-γ | CTGCCTGGTTACCTTCTTGCT | AACCCAACATAACACAAAGCCA | 79 | This study |
| IL-6 | TGGATGAGCTGAACTGTACCC | GCTTGCCAAGGATTGTGAGT | 119 | [34] |
| IL-10 | ACCAGAGACAAGCTCGAAAC | TCTTCCAGCAAAGATTTGTCTATC | 50 | [29] |
| Beta actin | AGATCATTGCCCCACCT | TGGAAGGCCCAGATTC | 123 | [3] |
| Zoo | Koala | Age (During Sampling) | Sex | KoRV Subtypes | WBC (×102/μL Blood) | Health Status | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| KoRV-A | KoRV-B | KoRV-C | KoRV-D | KoRV-E | KoRV-F | ||||||
| Kanazawa Zoo (n = 4) | KU_KAZ_01 | 1 yr 3 mo | F | P | P | N | N | ND | P | 115 | Healthy |
| KU_KAZ_02 | 6 yr 8 mo | M | P | P | N | N | ND | P | 75.5 | Healthy | |
| KU_KAZ_3 | 8 yr 1 mo | F | P | P | P | P | ND | N | 66.5 | Healthy | |
| KU_KAZ_4 | 4 yr 3 mo | F | P | P | N | P | ND | P | 95.5 | Healthy | |
| Tama Zoological Park (n = 2) | KU_TZ_05 | 6 yr | M | P | P | P | P | ND | N | NA | Healthy |
| KU_TZ_06 | 5 yr | M | P | P | N | P | ND | P | NA | Healthy | |
| Kobe Oji Zoo (n = 8) | KU_KZ_07 | 11 yr | F | P | P | N | N | ND | P | 45 | Healthy |
| KU_KZ_08 | 7 yr 2 mo | F | P | P | N | P | ND | P | 54 | Healthy | |
| KU_KZ_09 | 5 yr 5 mo | M | P | P | N | P | ND | N | 68 | Healthy | |
| KU_KZ_10 | 4 yr 11 mo | F | P | N | N | P | ND | P | 68.5 | Healthy | |
| KU_KZ_11 | 4 yr | M | P | P | P | P | ND | P | 30 | Healthy | |
| KU_KZ_12 | 2 yr 8 mo | F | P | P | N | P | ND | P | 147 | Healthy | |
| KU_KZ_13 | 2 yr 3 mo | F | P | P | N | N | ND | N | NA | Healthy | |
| KU_KZ_14 | 2 yr 3 mo | F | P | P | N | N | ND | N | 95.5 | Healthy | |
| Awaji Farm England Hill Zoo (n = 4) | KU_AZ_15 | 24 yr | F | P | N | N | N | ND | N | 50 | Healthy |
| KU_AZ_16 | 13 yr | F | P | P | N | N | ND | N | 58 | Healthy | |
| KU_AZ_17 | 12 yr | M | P | P | N | P | ND | N | 97 | Healthy | |
| KU_AZ_18 | 7 yr | M | P | P | P | ND | P | 42 | Healthy | ||
| Hirakawa Zoological Park (n = 4) | KU_HZ_19 | 1 yr | M | P | P | P | P | ND | N | 62 | Healthy |
| KU_HZ_20 | 1 yr | M | P | P | N | P | ND | N | 50 | Healthy | |
| KU_HZ_21 | 1 yr | F | P | P | N | P | ND | P | 82 | Healthy | |
| KU_HZ_22 | 1 yr | F | P | P | N | P | ND | N | 88 | Healthy | |
| Saitama Children’s Zoo (n = 5) | KU_SZ_23 | 3 yr | M | P | P | N | P | ND | P | NA | Healthy |
| KU_SZ_24 | 7 yr | F | P | P | P | P | ND | P | NA | Healthy | |
| KU_SZ_25 | 3 yr | F | P | P | P | P | ND | P | NA | Healthy | |
| KU_SZ_26 | 2 yr | F | P | P | P | P | ND | N | NA | Healthy | |
| KU_SZ_27 | 2 yr | F | P | P | P | P | ND | P | NA | Healthy | |
| Nagoya Higashiyama Zoo (n = 10) | KU_NZ_28 | 9 yr | M | P | P | P | P | ND | N | 76 | Healthy |
| KU_NZ_29 | 4 yr | M | P | P | N | P | ND | P | 85 | Healthy | |
| KU_NZ_30 | 11 yr | F | P | P | N | P | ND | P | 23 | Healthy | |
| KU_NZ_31 | 11 yr | F | P | P | P | P | ND | P | 51 | Healthy | |
| KU_NZ_32 | 7 yr | F | P | P | N | N | ND | N | 47 | Healthy | |
| KU_NZ_33 | 4 yr | F | P | P | N | P | ND | P | 57 | Healthy | |
| KU_NZ_34 | 10 mo | F | P | P | N | N | ND | N | 43 | Healthy | |
| KU_NZ_35 | 1 yr | F | P | P | N | N | ND | P | 69 | Healthy | |
| KU_NZ_36 | 11 mo | M | P | P | N | N | ND | N | 69 | Healthy | |
| KU_NZ_37 | 3 yr | F | P | P | N | N | ND | N | 66 | Healthy | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kayesh, M.E.H.; Hashem, M.A.; Maetani, F.; Goto, A.; Nagata, N.; Kasori, A.; Imanishi, T.; Tsukiyama-Kohara, K. Molecular Insights into Innate Immune Response in Captive Koala Peripheral Blood Mononuclear Cells Co-Infected with Multiple Koala Retrovirus Subtypes. Pathogens 2022, 11, 911. https://doi.org/10.3390/pathogens11080911
Kayesh MEH, Hashem MA, Maetani F, Goto A, Nagata N, Kasori A, Imanishi T, Tsukiyama-Kohara K. Molecular Insights into Innate Immune Response in Captive Koala Peripheral Blood Mononuclear Cells Co-Infected with Multiple Koala Retrovirus Subtypes. Pathogens. 2022; 11(8):911. https://doi.org/10.3390/pathogens11080911
Chicago/Turabian StyleKayesh, Mohammad Enamul Hoque, Md Abul Hashem, Fumie Maetani, Atsushi Goto, Noriko Nagata, Aki Kasori, Tetsuya Imanishi, and Kyoko Tsukiyama-Kohara. 2022. "Molecular Insights into Innate Immune Response in Captive Koala Peripheral Blood Mononuclear Cells Co-Infected with Multiple Koala Retrovirus Subtypes" Pathogens 11, no. 8: 911. https://doi.org/10.3390/pathogens11080911