
Lasiodiplodia fici sp. nov., Causing Leaf Spot on Ficus altissima in China

 , , ,
, , ,
Abstract
:1. Introduction
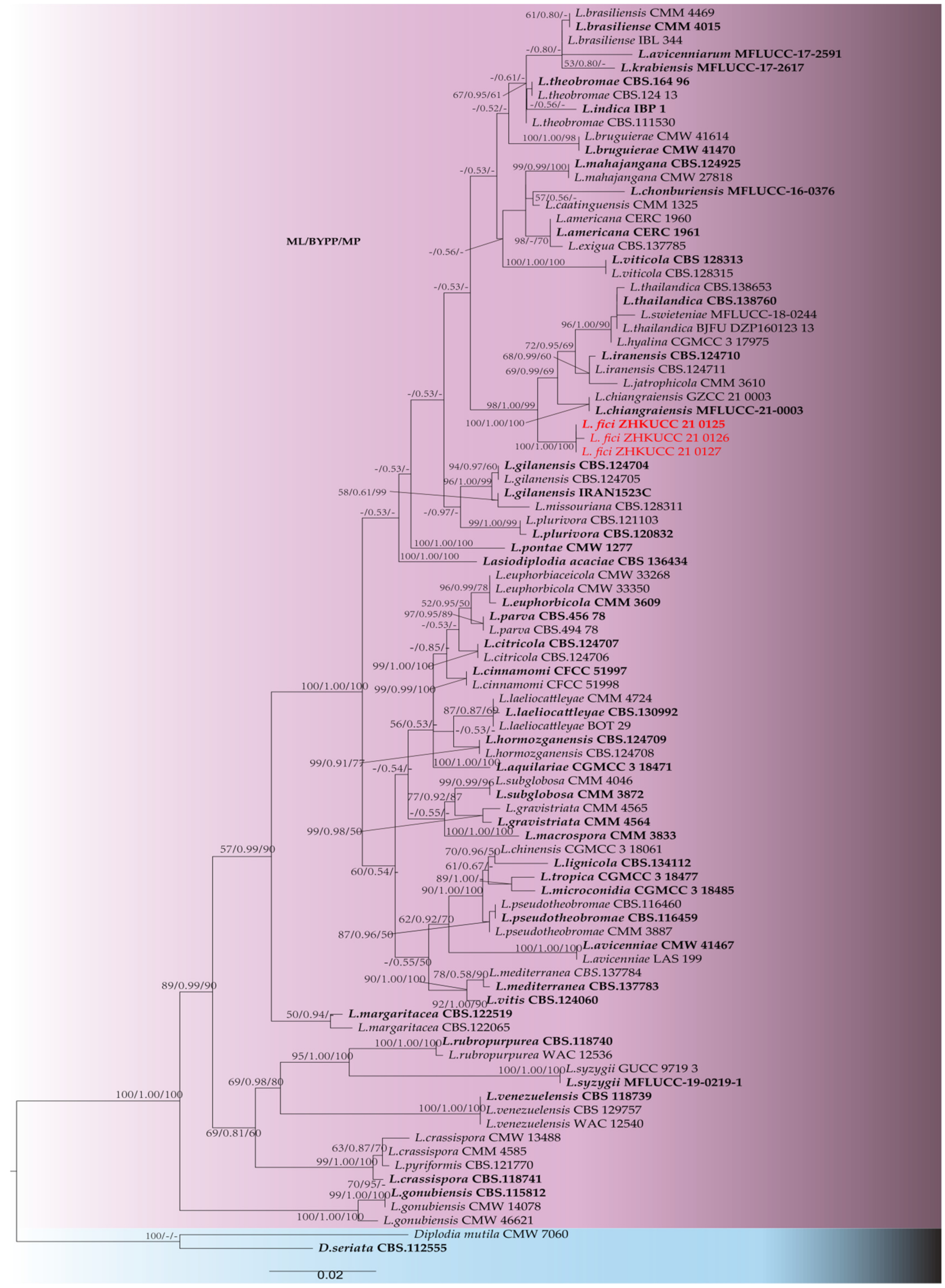
2. Results
2.1. Sampling and Isolation
2.2. Morphological and Molecular Characterisation
2.3. Pathogenicity Assay
3. Discussion
4. Materials and Methods
4.1. Samples Collection and Isolation
4.2. DNA Extraction, Amplification and Sequencing
4.3. Phylogenetic Analysis
4.4. Pathogenicity Assay
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brussaard, L. Biodiversity and ecosystem functioning in soil. Ambio 1997, 26, 563–570. [Google Scholar]
- Hyde, K.; Jeewon, R.; Chen, Y.J.; Bhunjun, B.C.; Calabon, M.S.; Jiang, H.B.; Lin, C.G.; Norphanphoun, C.; Sysouphanthong, P.; Pem, D.; et al. The numbers of fungi: Is the descriptive curve flattening? Fungal Divers. 2020, 103, 219–271. [Google Scholar] [CrossRef]
- Chethana, K.W.T.; Li, X.; Zhang, W.; Hyde, K.D.; Yan, J.Y. Trail of decryption of molecular research on Botryosphaeriaceae in woody plants. Phytopathol. Mediterr. 2016, 55, 147–171. [Google Scholar]
- Manawasinghe, I.S.; Phillips, A.J.L.; Hyde, K.D.; Chethana, K.W.T.; Zhang, W.; Zhao, W.S.; Yan, J.Y.; Li, X.H. Mycosphere Essays 14: Assessing the aggressiveness of plant pathogenic Botryosphaeriaceae. Mycosphere 2016, 7, 883–892. [Google Scholar] [CrossRef]
- Yan, J.Y.; Zhao, W.S.; Chen, Z.; Xing, Q.K.; Zhang, W.; Chethana, K.W.T.; Xue, M.F.; Xu, J.P.; Phillips, A.J.L.; Wang, Y.; et al. Comparative genome and transcriptome analyses reveal adaptations to opportunistic infections in woody plant degrading pathogens of Botryosphaeriaceae. DNA Res. 2018, 25, 87–102. [Google Scholar] [CrossRef] [Green Version]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2006, 55, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Dissanayake, A.J.; Manawasinghe, I.S.; Rathnayaka, A.R.; Liu, J.K.; Promputtha, I.; Hyde, K.D. https://botryosphaeriales.org/, an online platform for up-to-date classification and account of taxa of Botryosphaeriales. Database 2021, 2021, baab061. [Google Scholar] [CrossRef]
- Theissen, F.; Sydow, H. Vorentwürfe zu den Pseudosphaeriales. Ann. Mycol. 1918, 16, 1–34. [Google Scholar]
- Schoch, C.L.; Shoemaker, R.A.; Seifert, K.A.; Hambleton, S.; Spatafora, J.W.; Crous, P.W. A multigene phylogeny of the Dothideomycetes using four nuclear loci. Mycologia 2006, 98, 1041–1052. [Google Scholar] [CrossRef]
- Slippers, B.; Crous, P.W.; Jami, F.; Groenewald, J.Z.; Wingfield, M.J. Diversity in the Botryosphaeriales: Looking back, looking forward. Fungal Biol. 2017, 121, 307–321. [Google Scholar] [CrossRef]
- Yang, T.; Groenewald, J.Z.; Cheewangkoon, R.; Jami, F.; Abdollahzadeh, J.; Lombard, L.; Crous, P.W. Families, genera, and species of Botryosphaeriales. Fungal Biol. 2017, 121, 322–346. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Groenewald, J.Z.; Lombard, L.; Schumacher, R.K.; Phillips, A.J.L.; Crous, P.W. Evaluating species in Botryosphaeriales. Persoonia 2021, 46, 63–115. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdoğdu, M.; Selçuk, F.; Rajeshkumar, K.C.; Aptroot, A.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2021, 13, 53–453. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.; Tedersoo, L.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; et al. Outline of Fungi and funguslike taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Clendenin, I. Lasiodiplodia E. & E., n. gen. Bot Gaz 1896, 21, 92. [Google Scholar]
- Jayawardena, R.S.; Hyde, K.D.; Jeewon, R.; Ghobad-Nejhad, M.; Wanasinghe, D.N.; Liu, N.; Phillips, A.J.; Oliveira-Filho, J.R.C.; da Silva, G.A.; Gibertoni, T.B.; et al. One stop shop II: Taxonomic update with molecular phylogeny for important phytopathogenic genera: 26–50. Fungal Divers. 2019, 94, 41–129. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Battany, M.; Bettiga, L.J.; Gispert, C.; McGourty, G.; Roncoroni, J.; Smith, R.J.; Verdegaal, P.; Gubler, W.D. Botryosphaeriaceae species spore-trapping studies in California vineyards. Plant Dis. 2010, 94, 717–724. [Google Scholar] [CrossRef] [Green Version]
- Urbez-Torres, J.R. The status of Botryosphaeriaceae species infecting grapevines. Phytopatho. Mediterr. 2011, 50, S5–S45. [Google Scholar]
- Whitelaw-Weckert, M.A.; Rahman, L.; Appleby, L.M.; Hall, A.; Clark, A.C.; Waite, H.; Hardie, W.J. Co-infection by B otryosphaeriaceae and I lyonectria spp. fungi during propagation causes decline of young grafted grapevines. Plant Pathol. 2013, 62, 1226–1237. [Google Scholar] [CrossRef]
- Yan, J.Y.; Xie, Y.; Zhang, W.; Wang, Y.; Liu, J.K.; Hyde, K.D.; Seem, R.C.; Zhang, G.Z.; Wang, Z.Y.; Yao, S.W.; et al. Species of Botryosphaeriaceae involved in grapevine dieback in China. Fungal Divers. 2013, 61, 221–236. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Deidda, A.; Scanu, B.; Franceschini, A.; Serra, S.; Berraf-Tebbal, A.; Zouaoui Boutiti, M.; Ben Jamâa, M.L.; Phillips, A.J. Diversity of Botryosphaeriaceae species associated with grapevine and other woody hosts in Italy, Algeria and Tunisia, with descriptions of Lasiodiplodia exigua and Lasiodiplodia mediterranea sp. nov. Fungal Divers. 2015, 71, 201–214. [Google Scholar] [CrossRef]
- Coutinho, T.A.; Brady, C.L.; Van der Vaart, M.; Venter, S.N.; Telechea, N.; Rolfo, M.; Perez, C.; Wingfield, M.J. A new shoot and stem disease of Eucalyptus species caused by Erwinia psidii. Australas. Plant Path. 2011, 40, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Farr, D.F.; Rossman, A.Y.; Fungal Databases, U.S. National Fungus Collections, ARS, USDA. Available online: https://nt.ars-grin.gov/fungaldatabases (accessed on 16 May 2022).
- Moustafa, A.A. A threatened introduced species (Ficus benghalensis L.) in Ismailia, Egypt. In Modern Fruit Industry; IntechOpen: London, UK, 2020. [Google Scholar]
- Edlin, H.; Nimmo, M. The Illustrated Encyclopedia of Trees. In Salamander; Timbers and Forests of the World: London, UK, 1978; pp. 220–221. [Google Scholar]
- Henley, D. An Overview of Ficus for Interior Landscapes. Greenhouse 1991, 9, 62–64. [Google Scholar]
- Henley, R.W.; Poole, R.T. Evaluation of selected ornamental figs for interior use. In Proceedings of the Annual Meeting of the Florida State Horticulture Society, Tampa, FL, USA, 31 October–2 November 1989. [Google Scholar]
- Villard, C.; Larbat, R.; Munakata, R.; Hehn, A. Defence mechanisms of Ficus: Pyramiding strategies to cope with pests and pathogens. Planta 2019, 249, 617–633. [Google Scholar] [CrossRef]
- Alizadeh, A.; Javan-Nikkhah, M.; Nourmohammadi Nazarian, R.; Liu, F.; Zare, R.; Fotouhifar, K.B.; Stukenbrock, E.H.; Damm, U. New species of Colletotrichum from wild Poaceae and Cyperaceae plants in Iran. Mycologia 2022, 114, 89–113. [Google Scholar] [CrossRef]
- Abdollahzadeh, J.; Javadi, A.; Goltapeh, E.M.; Zare, R.; Phillips, A.J.L. Phylogeny and morphology of four new species of Lasiodiplodia from Iran. Persoonia 2010, 25, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Dissanayake, A.J.; Chethana, K.T.; Hyde, K.D.; Liu, J.K. Morpho-phylogenetic evidence reveals Lasiodiplodia chiangraiensis sp. nov. (Botryosphaeriaceae) associated with woody hosts in northern Thailand. Phytotaxa 2021, 508, 142–154. [Google Scholar] [CrossRef]
- Trakunyingcharoen, T.; Lombard, L.; Groenewald, J.Z.; Cheewangkoon, R.; To-Anun, C.; Crous, P.W. Caulicolous Botryosphaeriales from Thailand. Persoonia 2015, 34, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Senanayake, I.C.; Jeewon, R.; Hyde, K.D.; Bhat, J.D.; Cheewangkoon, R. Taxonomy and phylogeny of Leptosillia cordylinea sp. nov. from China. Phytotaxa 2020, 435, 213–226. [Google Scholar] [CrossRef]
- Burgess, T.I.; Tan, Y.P.; Garnas, J.; Edwards, J.; Scarlett, K.A.; Shuttleworth, L.A.; Daniel, R.; Dann, E.K.; Parkinson, L.E.; Dinh, Q.; et al. Current status of the Botryosphaeriaceae in Australia. Australas. Plant Pathol. 2019, 48, 35–44. [Google Scholar] [CrossRef]
- Rehab, M.A.; Rashed, M.F.; Ammar, M.I.; El-Morsy, S.A. Dieback and sooty canker of Ficus trees in Egypt and its control. Pak. J. 2014, 17, 364–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wei, H.; Du, G.; Zhu, L.; Song, Q.; Hu, Y.; Wang, E.; Wang, M.; Fan, X. First Report of Lasiodiplodia theobromae Causing Stem Canker on Common Fig (Ficus carica) in Zhejiang Province of China. Plant Dis. 2018, 102, 2656–2657. [Google Scholar] [CrossRef]
- Urtiaga, R. Indice de Enfermedadesy Desordenes en Plantas de Venezuela—Addendum; Published by the Author; 2004. Available online: https://www.researchgate.net/profile/Radames-Urtiaga/publication/315148773_INDICE_DE_ENFERMEDADES_Y_DESORDENES_EN_PLANTAS_DE_VENEZUELA/links/58cbef1fa6fdccdf531c6f8f/INDICE-DE-ENFERMEDADES-Y-DESORDENES-EN-PLANTAS-DE-VENEZUELA.pdf (accessed on 20 March 2022).
- Seo, Y.; Back, C.G.; Park, M.J.; Park, J.H. First Report of Lasiodiplodia theobromae Causing Canker and Dieback of Common Fig in Korea. Plant Dis. 2019, 103, 1023. [Google Scholar] [CrossRef]
- Çeliker, N.M.; Michailides, T.J. First report of Lasiodiplodia theobromae causing canker and shoot blight of fig in Turkey. New Dis. Rep. 2012, 25, 12. [Google Scholar] [CrossRef]
- Kobayashi, T. Index of Fungi Inhabiting Woody Plants in Japan; Zenkoku-Noson-Kyoiku Kyokai Publishing: Tokyo, Japan, 2007. [Google Scholar]
- Mohali, S.R.; Castro-Medina, F.; Úrbez-Torres, J.R.; Gubler, W.D. First report of Lasiodiplodia theobromae and L. venezuelensis associated with blue stain on Ficus insipida wood from the Natural Forest of Venezuela. Forest Pathol. 2017, 47, e12355. [Google Scholar] [CrossRef]
- French, A.M. California Plant Disease Host Index; California Department of Food and Agriculture, Division of Plant Industry: Sacramento, CA, USA, 1989.
- Sakalidis, M.L.; Hardy, G.E.S.J.; Burgess, T.I. Endophytes as potential pathogens of the baobab species Adansonia gregorii: A focus on the Botryosphaeriaceae. Fungal Ecol. 2011, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Manawasinghe, I.S.; Zhang, W.; Li, X.; Zhao, W.; Chethana, K.T.; Xu, J.; Chen, Z.; Dissanayaka, A.J.; Mugnai, L.; Úrbez-Torres, J.R.; et al. Novel microsatellite markers reveal multiple origins of Botryosphaeria dothidea causing the Chinese grapevine trunk disease. Fungal Ecol. 2018, 33, 134–142. [Google Scholar] [CrossRef]
- Pavlic-Zupanc, D.; Wingfield, M.J.; Boissin, E.; Slippers, B. The distribution of genetic diversity in the Neofusicoccum parvum/N. ribis complex suggests structure correlated with level of disturbance. Fungal Ecol. 2015, 13, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Mohali, S.; Burgess, T.I.; Wingfield, M.J. Diversity and host association of the tropical tree endophyte Lasiodiplodia theobromae revealed using simple sequence repeat markers. Forest Pathol. 2005, 35, 385–396. [Google Scholar] [CrossRef]
- Yongkang, H. Mulberry cultivation and utilization in China. In Proceedings of the Electronic Conference on Mulberry for Animal Production (Morus 1-L), Rome, Italy; 2002. [Google Scholar]
- Dong, Z.Y.; Huang, Y.H.; Manawasinghe, I.S.; Wanasinghe, D.N.; Liu, J.W.; Shu, Y.-X.; Zhao, M.P.; Xiang, M.M.; Luo, M. Stagonosporopsis pogostemonis: A Novel Ascomycete Fungus Causing Leaf Spot and Stem Blight on Pogostemon cablin (Lamiaceae) in South China. Pathogens 2021, 10, 1093. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Carbone, I.; Anderson, J.B.; Kohn, L.M. Patterns of descent in clonal lineages and their multilocus fingerprints are resolved with combined gene genealogies. Evolution 1999, 53, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Xiao, X.E.; Wang, W.; Crous, P.W.; Wang, H.K.; Jiao, C.; Huang, F.; Pu, Z.X.; Zhu, Z.R.; Li, H.Y. Species of Botryosphaeriaceae associated with citrus branch diseases in China. Persoonia 2021, 47, 106–135. [Google Scholar] [CrossRef]
- Swofford, D.L. Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4.0b10. Mac. Vers. 2002, 56, 1776–1788. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Bio. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The faces of fungi database: Fungal names linked with morphology, molecular and human attributes. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Jayawardena, S.; Hyde, K.D.; de Farias, A.R.G.; Bhunjun, C.S.; Ferdinandez, H.S.; Manamgoda, D.S.; Udayanga, D.; Herath, I.S.; Thambugala, K.M.; Manawasinghe, I.S.; et al. What is a species in fungal plant pathogens? Fungal Divers. 2021, 109, 239–266. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Phillips, A.J.L.; Xu, J.; Balasuriya, A.; Hyde, K.D.; tępień, L.; Harischandra, D.L.; Karunarathna, A.; Yan, J.; Weerasinghe, J.; et al. Defining a species in plant pathology: Beyond the species level. Fungal Divers. 2021, 109, 267–282. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conidial Dimensions (μm) | L/W Ratio | Reference | |
|---|---|---|---|
| Lasiodiplodia fici | 17–28 × 9–14 | 2.0 | This study |
| Lasiodiplodia chiangraiensis | 22–27 × 13–15 | 1.9 | [30] |
| Lasiodiplodia iranensis | 17–23 × 11–14 | 1.6 | [31] |
| Lasiodiplodia thailandica | 22–25 × 13–15 | 1.7 | [32] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, G.; Manawasinghe, I.S.; Phillips, A.J.L.; You, C.; Jayawardena, R.S.; Luo, M.; Hyde, K.D. Lasiodiplodia fici sp. nov., Causing Leaf Spot on Ficus altissima in China. Pathogens 2022, 11, 840. https://doi.org/10.3390/pathogens11080840
Xia G, Manawasinghe IS, Phillips AJL, You C, Jayawardena RS, Luo M, Hyde KD. Lasiodiplodia fici sp. nov., Causing Leaf Spot on Ficus altissima in China. Pathogens. 2022; 11(8):840. https://doi.org/10.3390/pathogens11080840
Chicago/Turabian StyleXia, GuiYan, Ishara S. Manawasinghe, Alan J. L. Phillips, ChunPing You, Ruvishika S. Jayawardena, Mei Luo, and Kevin D. Hyde. 2022. "Lasiodiplodia fici sp. nov., Causing Leaf Spot on Ficus altissima in China" Pathogens 11, no. 8: 840. https://doi.org/10.3390/pathogens11080840