
Beyond CC398: Characterisation of Other Tetracycline and Methicillin-Resistant Staphylococcus aureus Genetic Lineages Circulating in Spanish Hospitals

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
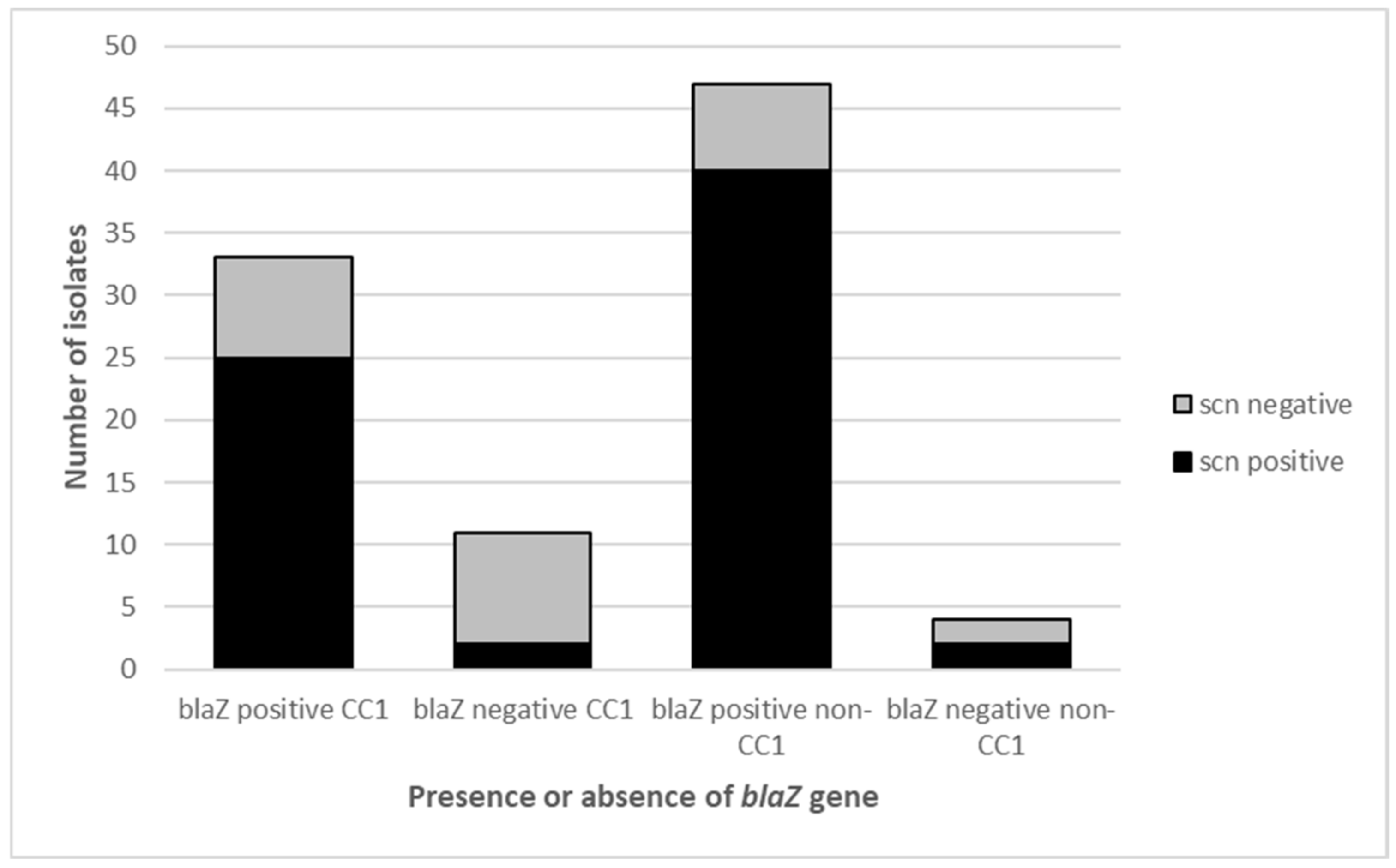
2.1. Molecular Characterisation of MRSA Isolates
2.2. Sample Origin
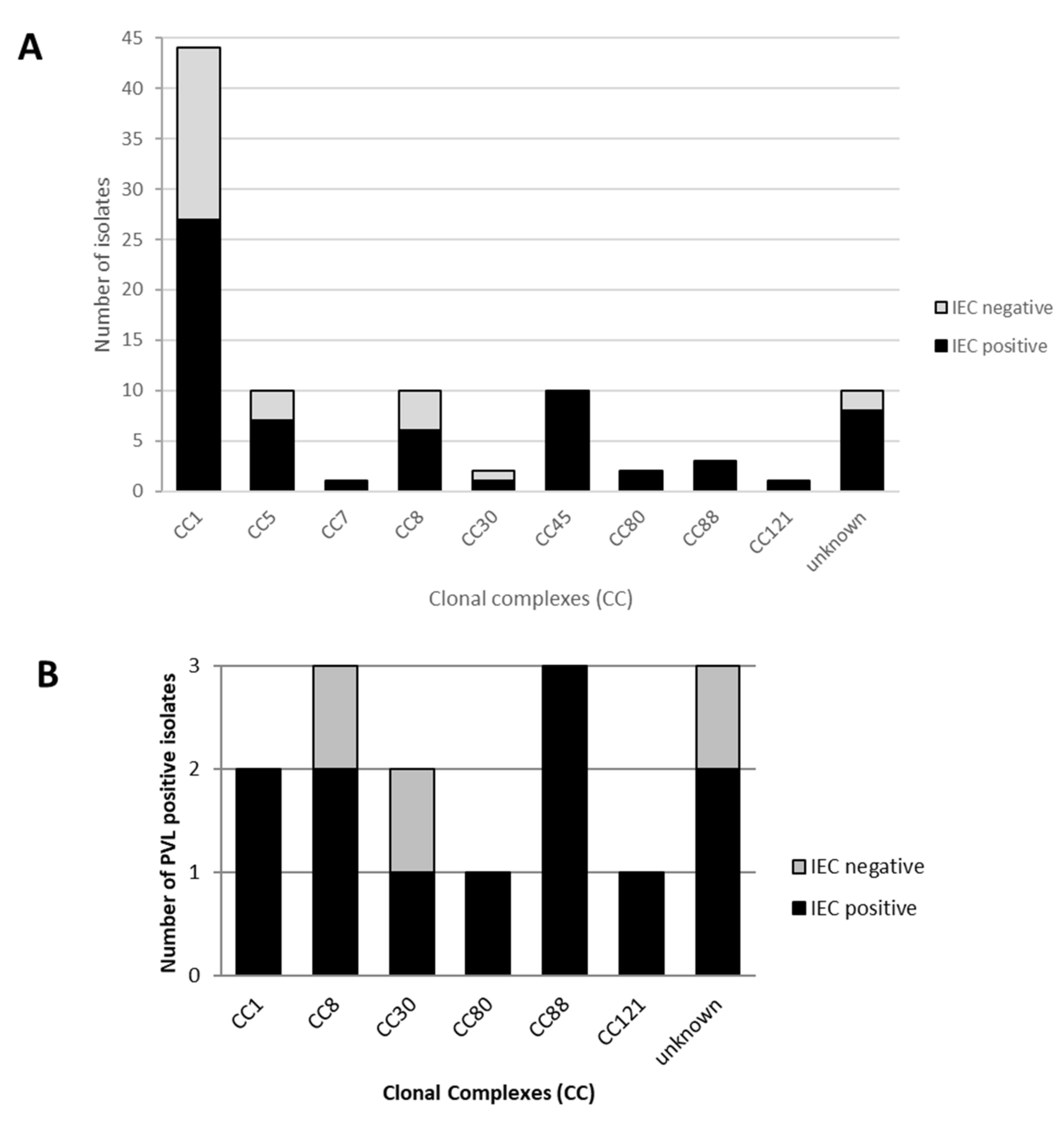
2.3. Presence of the IEC and PVL Toxin
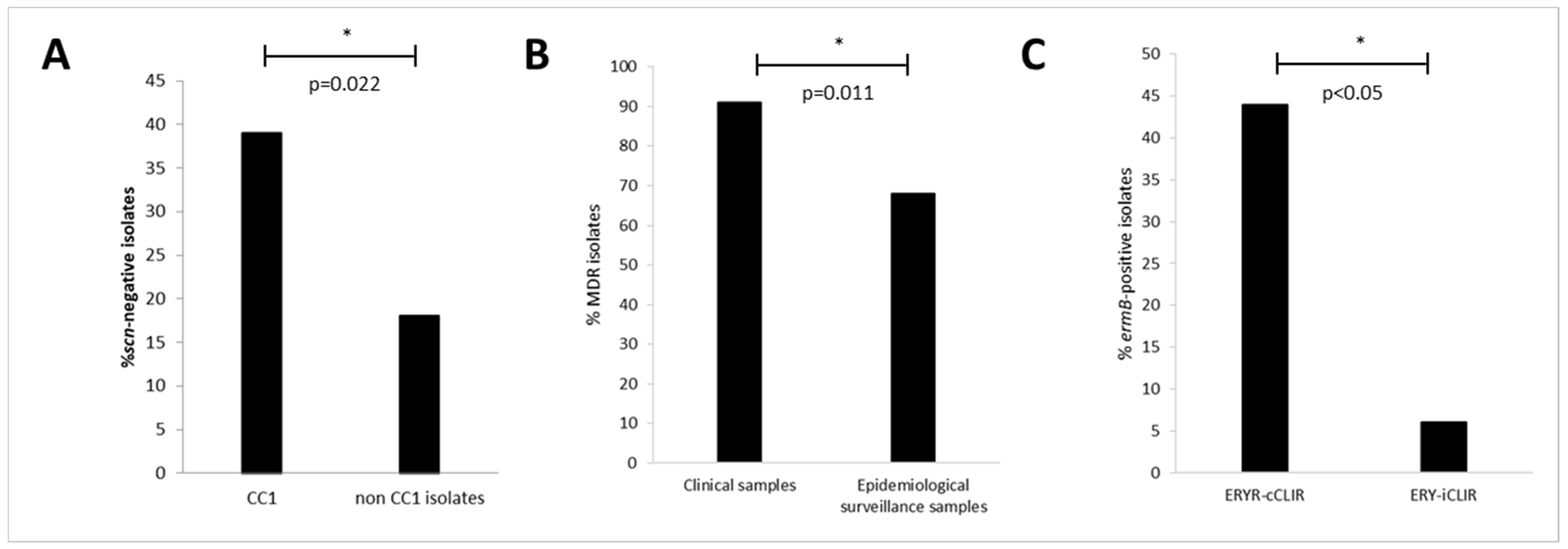
2.4. Multidrug Resistance (MDR) Phenotypes
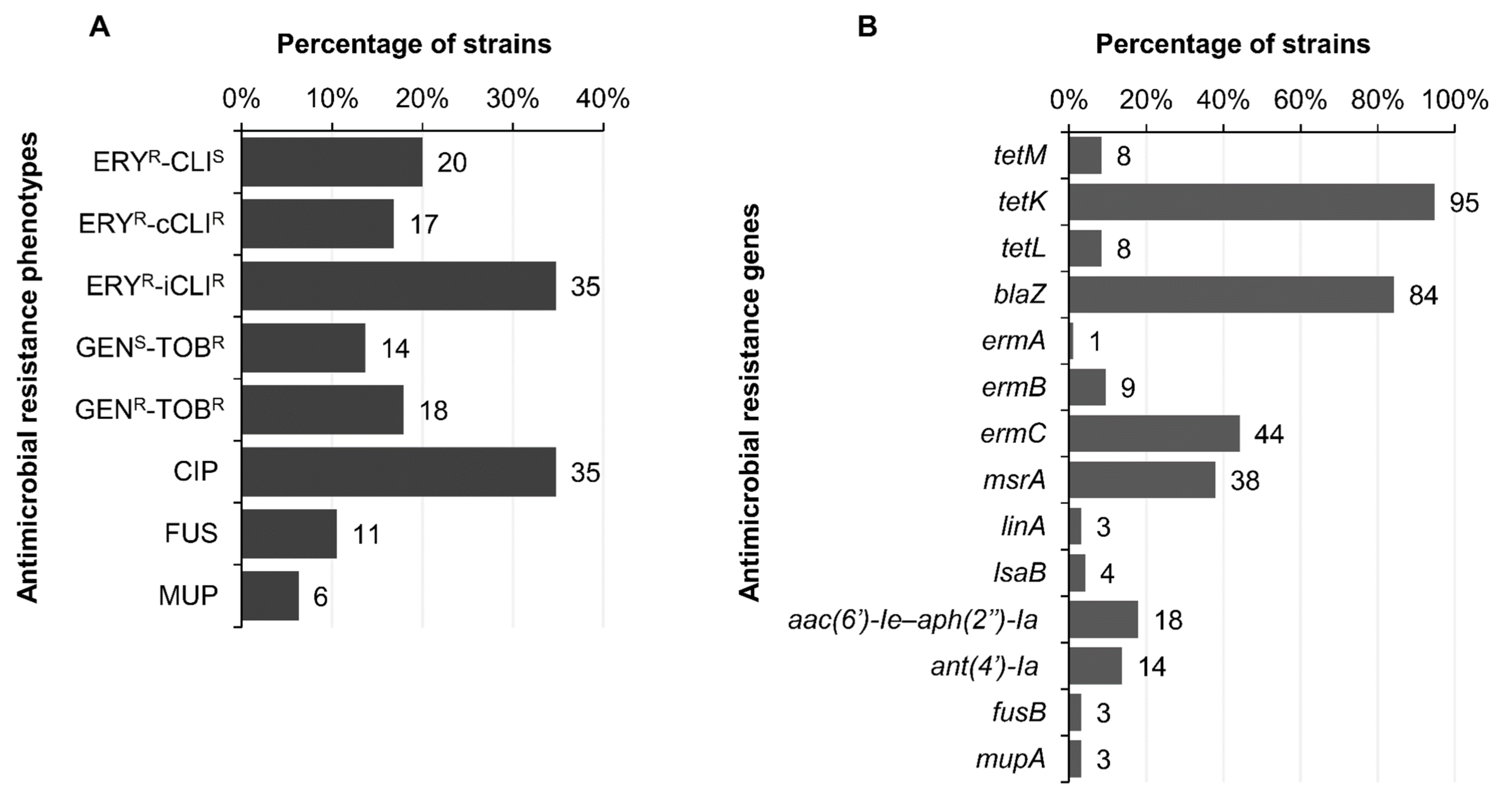
2.5. Antimicrobial Resistance Phenotypes and Genotypes
2.6. Analysis of a Possible Relationship between MRSA CC1 and Pig Farming Density of Adjoining Region
3. Discussion
4. Materials and Methods
4.1. Selection of Isolates
4.2. Molecular Typing
4.3. Detection of the Immune Evasion Cluster System (IEC) and the Panton–Valentine Leucocidin (PVL) Genes
4.4. Antimicrobial Susceptibility Testing
4.5. Detection of Antimicrobial Resistance Genes
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lakhundi, S.; Zhang, K. Methicillin-resistant Staphylococcus aureus: Molecular characterization, evolution, and epidemiology. Clin. Microbiol. Rev. 2018, 31, e00020-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, C.; Porres-Osante, N.; Benito, D.; Zarazaga, M.; Torres, C.; Rojo-Bezares, B.; Sáenz, Y.; Crettaz, J.; Olarte, I. Changes in genetic lineages, resistance, and virulence in clinical methicillin-resistant Staphylococcus aureus in a Spanish hospital. J. Infect. Chemother. 2013, 19, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Elstrøm, P.; Grøntvedt, C.A.; Gabrielsen, C.; Stegger, M.; Angen, Ø.; Åmdal, S.; Enger, H.; Urdahl, A.M.; Jore, S.; Steinbakk, M.; et al. Livestock-associated MRSA CC1 in Norway; introduction to pig farms, zoonotic transmission, and eradication. Front. Microbiol. 2019, 10, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Wu, F. Livestock-associated methicillin-resistant Staphylococcus aureus (LA-MRSA) colonisation and infection among livestock workers and veterinarians: A systematic review and meta-analysis. Occup. Environ. Med. 2021, 78, 530–540. [Google Scholar] [CrossRef]
- Larsen, J.; Clasen, J.; Hansen, J.E.; Paulander, W.; Petersen, A.; Larsen, A.R.; Frees, D. Copresence of tet (K) and tet (M) in livestock-associated methicillin-resistant Staphylococcus aureus Clonal Complex 398 is associated with increased fitness during exposure to sublethal concentrations of tetracycline. Antimicrob. Agents Chemother. 2016, 60, 4401–4403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, M.J.A.; Stephan, R.; Johler, S. Draft genome sequence of Staphylococcus aureus S681, a tetracycline-sensitive livestock-associated methicillin-resistant Clonal Complex 398 Strain. Genome Announc. 2017, 5, e00805-17. [Google Scholar] [CrossRef] [Green Version]
- Benito, D.; Lozano, C.; Rezusta, A.; Ferrer, I.; Vasquez, M.A.; Ceballos, S.; Zarazaga, M.; Revillo, M.J.; Torres, C. Characterization of tetracycline and methicillin resistant Staphylococcus aureus strains in a Spanish hospital: Is livestock-contact a risk factor in infections caused by MRSA CC398? Int. J. Med. Microbiol. 2014, 304, 1226–1232. [Google Scholar] [CrossRef]
- Ruiz-Ripa, L.; Gómez, P.; Alonso, C.A.; Camacho, M.C.; De La Puente, J.; Fernández-Fernández, R.; Ramiro, Y.; Quevedo, M.A.; Blanco, J.M.; Zarazaga, M.; et al. Detection of MRSA of lineages CC130-mecC and CC398-mecA and Staphylococcus delphini-lnu (A) in magpies and cinereous vultures in Spain. Microb. Ecol. 2019, 78, 409–415. [Google Scholar] [CrossRef]
- Butaye, P.; Argudín, M.A.; Smith, T.C. Livestock-associated MRSA and its current evolution. Curr. Clin. Microbiol. Rep. 2016, 3, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Lozano, C.; Fernández-Fernández, R.; Ruiz-Ripa, L.; Gómez, P.; Zarazaga, M.; Torres, C. Human mecC-carrying MRSA: Clinical implications and risk factors. Microorganisms 2020, 8, 1615. [Google Scholar] [CrossRef]
- Aspiroz, C.; Lozano, C.; Vindel, A.; Lasarte, J.J.; Zarazaga, M.; Torres, C. Skin Lesion caused by ST398 and ST1 MRSA, Spain. Emerg. Infect. Dis. 2010, 16, 157–159. [Google Scholar] [CrossRef] [PubMed]
- Fetsch, A.; Etter, D.; Johler, S. Livestock-associated meticillin-resistant Staphylococcus aureus—Current situation and impact from a One Health perspective. Curr. Clin. Microbiol. Rep. 2021, 8, 103–113. [Google Scholar] [CrossRef]
- Hansen, J.E.; Ronco, T.; Stegger, M.; Sieber, R.N.; Fertner, M.E.; Martin, H.L.; Farre, M.; Toft, N.; Larsen, A.R.; Pedersen, K. LA-MRSA CC398 in dairy cattle and veal calf farms indicates spillover from pig production. Front. Microbiol. 2019, 10, 2733. [Google Scholar] [CrossRef] [Green Version]
- Ceballos, S.; Aspiroz, C.; Ruiz-Ripa, L.; Reynaga, E.; Azcona-Gutiérrez, J.M.; Rezusta, A.; Seral, C.; Antoñanzas, F.; Torres, L.; López, A.R.; et al. Epidemiology of MRSA CC398 in hospitals located in Spanish regions with different pig-farming densities: A multicentre study. J. Antimicrob. Chemother. 2019, 74, 2157–2161. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.S.; De Lencastre, H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-resistant Staphylococcus aureus. Nat. Rev. Dis. Prim. 2018, 4, 18033. [Google Scholar] [CrossRef]
- Vignaroli, C.; Varaldo, P.E.; Camporese, A. Methicillin-resistant Staphylococcus aureus USA400 clone, Italy. Emerg. Infect. Dis. 2009, 15, 995–996. [Google Scholar] [CrossRef]
- Petersen, A.; Stegger, M.; Heltberg, O.; Christensen, J.; Zeuthen, A.; Knudsen, L.; Urth, T.; Sorum, M.; Schouls, L.; Larsen, J.; et al. Epidemiology of methicillin-resistant Staphylococcus aureus carrying the novel mecC gene in Denmark corroborates a zoonotic reservoir with transmission to humans. Clin. Microbiol. Infect. 2013, 19, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Cooke, F.; Gkrania-Klotsas, E.; Howard, J.; Stone, M.; Kearns, A.; Ganner, M.; Carmichael, A.; Brown, N. Clinical, molecular and epidemiological description of a cluster of community-associated methicillin-resistant Staphylococcus aureus isolates from injecting drug users with bacteraemia. Clin. Microbiol. Infect. 2010, 16, 921–926. [Google Scholar] [CrossRef] [Green Version]
- Otter, J.A.; Havill, N.L.; Boyce, J.M.; French, G.L. Comparison of community-associated meticillin-resistant Staphylococcus aureus from teaching hospitals in London and the USA, 2004–2006: Where is USA300 in the UK? Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 835–839. [Google Scholar] [CrossRef]
- Alba, P.; Feltrin, F.; Cordaro, G.; Porrero, M.C.; Kraushaar, B.; Argudín, M.A.; Nykäsenoja, S.; Monaco, M.; Stegger, M.; Aarestrup, F.; et al. Livestock-associated methicillin resistant and methicillin susceptible Staphylococcus aureus Sequence Type (CC)1 in European farmed animals: High genetic relatedness of isolates from Italian cattle herds and humans. PLoS ONE 2015, 10, e0137143. [Google Scholar] [CrossRef] [Green Version]
- Franco, A.; Hasman, H.; Iurescia, M.; Lorenzetti, R.; Stegger, M.; Pantosti, A.; Feltrin, F.; Ianzano, A.; Porrero, M.C.; Liapi, M.; et al. Molecular characterization of spa type t127, sequence type 1 methicillin-resistant Staphylococcus aureus from pigs. J. Antimicrob. Chemother. 2011, 66, 1231–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Normanno, G.; Dambrosio, A.; Lorusso, V.; Samoilis, G.; Di Taranto, P.; Parisi, A. Methicillin-resistant Staphylococcus aureus (MRSA) in slaughtered pigs and abattoir workers in Italy. Food Microbiol. 2015, 51, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Lozano, C.; Rezusta, A.; Gómez, P.; Gómez-Sanz, E.; Báez, N.; Martin-Saco, G.; Zarazaga, M.; Torres, C. High prevalence of spa types associated with the clonal lineage CC398 among tetracycline-resistant methicillin-resistant Staphylococcus aureus isolates in a Spanish hospital. J. Antimicrob. Chemother. 2012, 67, 330–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monecke, S.; Coombs, G.; Shore, A.C.; Coleman, D.C.; Akpaka, P.; Borg, M.; Chow, H.; Ip, M.; Jatzwauk, L.; Jonas, D.; et al. A field guide to pandemic, epidemic and sporadic clones of methicillin-resistant Staphylococcus aureus. PLoS ONE 2011, 6, e17936. [Google Scholar] [CrossRef] [PubMed]
- Earls, M.R.; Shore, A.C.; Brennan, G.I.; Simbeck, A.; Schneider-Brachert, W.; Vremerǎ, T.; Dorneanu, O.S.; Slickers, P.; Ehricht, R.; Monecke, S.; et al. A novel multidrug-resistant PVL-negative CC1-MRSA-IV clone emerging in Ireland and Germany likely originated in South-Eastern Europe. Infect. Genet. Evol. 2019, 69, 117–126. [Google Scholar] [CrossRef]
- Coombs, G.W.; Monecke, S.; Pearson, J.C.; Tan, H.L.; Chew, Y.K.; Wilson, L.; Ehricht, R.; O’Brien, F.G.; Christiansen, K.J. Evolution and diversity of community-associated methicillin-resistant Staphylococcus aureus in a geographical region. BMC Microbiol. 2011, 11, 215. [Google Scholar] [CrossRef] [Green Version]
- Earls, M.R.; Kinnevey, P.M.; Brennan, G.I.; Lazaris, A.; Skally, M.; O’Connell, B.; Humphreys, H.; Shore, A.C.; Coleman, D.C. The recent emergence in hospitals of multidrug-resistant community-associated sequence type 1 and spa type t127 methicillin-resistant Staphylococcus aureus investigated by whole-genome sequencing: Implications for screening. PLoS ONE 2017, 12, e0175542. [Google Scholar] [CrossRef]
- Earls, M.R.; Steinig, E.J.; Monecke, S.; Castruita, J.A.S.; Simbeck, A.; Schneider-Brachert, W.; Vremerǎ, T.; Dorneanu, O.S.; Loncaric, I.; Bes, M.; et al. Exploring the evolution and epidemiology of European CC1- MRSA-IV: Tracking a multidrug-resistant community-associated meticillin-resistant Staphylococcus aureus clone. Microb. Genom. 2021, 7, 000601. [Google Scholar] [CrossRef]
- Gruteke, P.; Ho, P.L.; Haenen, A.; Lo, W.-U.; Lin, C.-H.; De Neeling, A.J. MRSA spa t1081, a highly transmissible isolate endemic to Hong Kong, China, in the Netherlands. Emerg. Infect. Dis. 2015, 21, 1074–1076. [Google Scholar] [CrossRef]
- Tsao, F.-Y.; Kou, H.-W.; Huang, Y.-C. Dissemination of methicillin-resistant Staphylococcus aureus sequence type 45 among nursing home residents and staff in Taiwan. Clin. Microbiol. Infect. 2015, 21, 451–458. [Google Scholar] [CrossRef] [Green Version]
- Van Gaalen, R.D.; Hopman, H.A.; Haenen, A.; Van Den Dool, C. Staff exchange within and between nursing homes in The Netherlands and potential implications for MRSA transmission. Epidemiol. Infect. 2017, 145, 739–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, A.; Lim, V.W.; Khan, A.; Pettigrew, K.; Lye, D.C.B.; Kanagasabai, K.; Phua, K.; Krishnan, P.; Ang, B.; Marimuthu, K.; et al. MRSA Transmission Dynamics Among Interconnected Acute, Intermediate-Term, and Long-Term Healthcare Facilities in Singapore. Clin. Infect. Dis. 2017, 64 (Suppl. S2), S76–S81. [Google Scholar] [CrossRef] [Green Version]
- Cabrera, R.; Fernández-Barat, L.; Motos, A.; López-Aladid, R.; Vázquez, N.; Panigada, M.; Álvarez-Lerma, F.; López, Y.; Muñoz, L.; Castro, P.; et al. Molecular characterization of methicillin-resistant Staphylococcus aureus clinical isolates from the endotracheal tubes of patients with nosocomial pneumonia. Antimicrob. Resist. Infect. Control 2020, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Kwapisz, E.; Garbacz, K.; Kosecka-Strojek, M.; Schubert, J.; Bania, J.; Międzobrodzki, J. Presence of egc-positive major clones ST 45, 30 and 22 among methicillin-resistant and methicillin-susceptible oral Staphylococcus aureus strains. Sci. Rep. 2020, 10, 18889. [Google Scholar] [CrossRef]
- Ugolotti, E.; Di Marco, E.; Bandettini, R.; Biassoni, R. Genomic characterization of a paediatric MRSA outbreak by next-generation sequencing. J. Hosp. Infect. 2018, 98, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Rao, Q.; Shang, W.; Hu, X.; Rao, X. Staphylococcus aureus ST121: A globally disseminated hypervirulent clone. J. Med. Microbiol. 2015, 64, 1462–1473. [Google Scholar] [CrossRef]
- Founou, R.C.; Founou, L.L.; Allam, M.; Ismail, A.; Essack, S.Y. Genomic characterisation of Staphylococcus aureus ST121 isolated from hospitalised patients in South Africa. S. Afr. Med. J. 2018, 108, 1007. [Google Scholar] [CrossRef] [Green Version]
- Gharsa, H.; Slama, K.B.; Lozano, C.; Gómez-Sanz, E.; Klibi, N.; Ben Sallem, R.; Gómez, P.; Zarazaga, M.; Boudabous, A.; Torres, C. Prevalence, antibiotic resistance, virulence traits and genetic lineages of Staphylococcus aureus in healthy sheep in Tunisia. Vet. Microbiol. 2012, 156, 367–373. [Google Scholar] [CrossRef]
- Attili, A.R.; Bellato, A.; Robino, P.; Galosi, L.; Papeschi, C.; Rossi, G.; Fileni, E.; Linardi, M.; Cuteri, V.; Chiesa, F.; et al. Analysis of the antibiotic resistance profiles in methicillin-sensitive S. aureus pathotypes isolated on a commercial rabbit farm in Italy. Antibiotics 2020, 9, 673. [Google Scholar] [CrossRef]
- Mama, O.M.; Aspiroz, C.; Lozano, C.; Ruiz-Ripa, L.; Azcona, J.M.; Seral, C.; Cercenado, E.; López-Cerero, L.; Palacian, P.; Belles-Belles, A.; et al. Penicillin susceptibility among invasive MSSA infections: A multicentre study in 16 Spanish hospitals. J. Antimicrob. Chemother. 2021, 76, 2519–2527. [Google Scholar] [CrossRef]
- Gómez-Sanz, E.; Torres, C.; Lozano, C.; Fernández-Pérez, R.; Aspiroz, C.; Ruiz-Larrea, F.; Zarazaga, M. Detection, molecular characterization, and clonal diversity of methicillin-resistant Staphylococcus aureus CC398 and CC97 in Spanish slaughter pigs of different age groups. Foodborne Pathog. Dis. 2010, 7, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Van Wamel, W.J.B.; Rooijakkers, S.H.M.; Ruyken, M.; van Kessel, K.P.M.; Van Strijp, J.A.G. The innate immune modulators staphylococcal complement inhibitor and chemotaxis inhibitory protein of Staphylococcus aureus are located on β-hemolysin-converting bacteriophages. J. Bacteriol. 2006, 188, 1310–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lina, G.; Piémont, Y.; Godail-Gamot, F.; Bes, M.; Peter, M.-O.; Gauduchon, V.; Vandenesch, F.; Etienne, J. Involvement of Panton-Valentine leukocidin-producing Staphylococcus aureus in primary skin infections and pneumonia. Clin. Infect. Dis. 1999, 29, 1128–1132. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hospital Number a | Number of Isolates | Rates (%) | Density (per km2) | scnc (Number of Non CC398 Isolates) | Clonal Complex (Number of Non-CC398 Isolates) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MRSA | TetR-MRSA | TetR-MRSA CC398 | TetR-MRSA Non-CC398 | TetR-MRSA CC1 | TetR-MRSA/ MRSA | TetR-MRSA Non-CC398/TetR-MRSA | TetR-MRSA CC1/TetR-MRSA | TetR-MRSA CC1/TetR-MRSA Non-CC398 | Pigs | Inhabitants | + | − | ||
| H1 | 87 | 33 | 27 | 6 | 6 | 38 | 18 | 18 | 100 | 247.46 | 717.36 | 6 | 0 | CC1 (6) |
| H2 | 135 | 20 | 19 | 1 | 0 | 15 | 5 | 0 | 0 | 217.68 | 14.05 | 1 | 0 | CC45 (1) |
| H3 | 328 | 24 | 15 | 9 | 1 | 7 | 38 | 4 | 11 | 217.68 | 14.05 | 8 | 1 | CC1 (1), CC45 (8) |
| H4 | 251 | 34 | 18 | 16 | 12 | 14 | 47 | 35 | 75 | 142.66 | 55.20 | 8 | 8 | CC1 (12), CC5 (1), CC8 (2), CC88 (1) |
| H5 | 175 | 20 | 9 | 11 | 5 | 11 | 55 | 25 | 45 | 142.66 | 55.20 | 10 | 1 | CC1 (5), CC5 (1), CC8 (1), CC45 (1), ST5427 (1), unknown (2) |
| H6 | 76 | 9 | 7 | 2 | 2 | 12 | 22 | 22 | 100 | 142.66 | 55.20 | 1 | 1 | CC1 (2) |
| H7 | 42 | 6 | 3 | 3 | 2 | 14 | 50 | 33 | 67 | 142.66 | 55.20 | 1 | 2 | CC1 (2), CC80 (1) |
| H8 | 36 | 4 | 4 | 0 | 0 | 11 | 0 | 0 | 0 | 69.97 | 9.15 | - | - | - |
| H9 | 36 | 2 | 2 | 0 | 0 | 6 | 0 | 0 | 0 | 50.89 | 61.90 | - | - | - |
| H10 | 206 | 14 | 7 | 7 | 3 | 7 | 50 | 21 | 43 | 50.89 | 61.90 | 5 | 2 | CC1 (3), CC8 (1), CC80 (1), CC121 (1), unknown (1) |
| H11 | 84 | 7 | 3 | 4 | 2 | 8 | 57 | 29 | 50 | 42.10 | 138.18 | 3 | 1 | CC1 (2), CC8 (2) |
| H12 | 220 | 6 | 3 | 3 | 1 | 3 | 50 | 17 | 33 | 27.66 | 25.54 | 3 | 0 | CC1 (1), CC8 (1), unknown (1) |
| H13 | 42 | 2 | 0 | 2 | 0 | 5 | 100 | 0 | 0 | 27.66 | 25.54 | 0 | 2 | CC5 (2) |
| H14 | 112 | 6 | 4 | 2 | 1 | 5 | 33 | 17 | 50 | 18.31 | 62.51 | 0 | 2 | CC1 (1), CC8 (1) |
| H15 | 334 | 5 | 5 | 0 | 0 | 1 | 0 | 0 | 0 | 40.99 | 107.53 | - | - | - |
| H16 | 130 | 7 | 3 | 4 | 0 | 5 | 57 | 0 | 0 | 30.56 | 360.18 | 3 | 1 | CC8 (1), CC88 (1), unknown (2) |
| H17 | 315 | 12 | 0 | 12 | 4 | 4 | 100 | 33 | 33 | 20.85 | 810.66 | 12 | 0 | CC1 (4), CC5 (2), CC7 (1), ST2625 (2), unknown (3) |
| H18 | 277 | 7 | 3 | 4 | 2 | 3 | 57 | 29 | 50 | 20.19 | 517.95 | 0 | 4 | CC1 (2), CC5 (1), CC30 (1) |
| H19 | 371 | 13 | 5 | 8 | 2 | 4 | 62 | 15 | 25 | 00.45 | 109.06 | 7 | 1 | CC1 (2), CC5 (3), CC8 (1), CC30 (1), CC88 (1) |
| H20 | 126 | 1 | 0 | 1 | 1 | 1 | 100 | 100 | 100 | 00.45 | 109.06 | 1 | 0 | CC1 (1) |
| Total | 3383 | 232 | 137 b | 95 | 45 | 6.8 | 40.9 | 13.4 | 47.4 | - | - | 69 | 26 | - |
| Clonal Complex (CC) (%) | spa-Type | No of Isolates | Sample Origin (No of Isolates) | scn+ g | PVL+ h |
|---|---|---|---|---|---|
| CC1 (46%) | t127 | 38 | SSTI b (17), ES c (11), RTI d (5), blood (3), UTI e (1), SSI f (1) | 23 | 1 |
| t174, t693 | 2 | SSTI | 2 | 0 | |
| t1381 | 2 | ES (1), RTI (1) | 1 | 0 | |
| t1784 | 1 | Blood | 1 | 1 | |
| t2207 | 1 | ES | 0 | 0 | |
| CC5 (11%) | t002 | 5 | SSTI (2), ES (2), UTI (1) | 4 | 0 |
| t067 | 3 | SSTI (2), UTI (1) | 1 | 0 | |
| t688 | 1 | RTI | 1 | 0 | |
| t1594 | 1 | ES | 1 | 0 | |
| CC7 (1%) | t091 | 1 | SSTI | 1 | 0 |
| CC8 (11%) | t008 | 1 | SSTI | 1 | 1 |
| t064, t1476 | 2 | RTI | 2 | 0 | |
| t148 | 6 | SSTI (5), ES (1) | 3 | 2 | |
| t2849 | 1 | RTI | 0 | 0 | |
| CC30 (2%) | t665 | 2 | SSTI (1), SSI (1) | 1 | 2 |
| CC45 (11%) | t1081 | 10 | SSTI (6), ES (2), SSI (1), RTI (1) | 10 | 0 |
| CC80 (2%) | t044 | 1 | SSTI | 1 | 1 |
| t088 | 1 | SSTI | 1 | 0 | |
| CC88 (3%) | t690 | 2 | RTI | 2 | 2 |
| t4103 | 1 | UTI | 1 | 1 | |
| CC121 (1%) | t17234 a | 1 | SSTI | 1 | 1 |
| ST2625 (2%) | t17235 a | 2 | SSTI | 2 | 0 |
| ST5427 a (1%) | t17236 a | 1 | SSTI | 1 | 0 |
| Unknown (11%) | t437 | 1 | RTI | 1 | 1 |
| t992 | 1 | SSI | 1 | 0 | |
| t1354 | 1 | SSTI | 1 | 1 | |
| t1954, t3324 | 2 | SSTI | 2 | 0 | |
| t4725 | 2 | RTI | 0 | 1 | |
| t10419 | 3 | SSTI | 3 | 0 |
| spa-Type (No of Isolates) | Antimicrobial Resistance (Non Beta-Lactams) | No of blaZ Positive Isolates | No of scn Negative Isolates | No of PVL Positive Isolates | Origin (No of Isolates) c | |
|---|---|---|---|---|---|---|
| Phenotype a (No of Isolates) | Genotype b | |||||
| t127 (38) | TET (5) | tetK | 4 | 1 | 0 | ES (5) |
| ERY-TET (6) | tetL1, tetK, msrA | 6 | 1 | 0 | RTI (1), SSTI (5) | |
| CLI-ERY-TET (17) | tetK, ermB2, ermC15, msrA4, lsaB1 | 11 | 8 | 1 | ES (3), RTI (3), SSTI (7), SSI (1), blood (3) | |
| CIP-TET (1) | tetK | 1 | 0 | 0 | ES (1) | |
| ERY-CIP-TET (1) | tetK, msrA | 1 | 1 | 0 | RTI (1) | |
| CLI-ERY-CIP-TET (5) | tetK, ermB3, ermC, msrA1, lsaB1 | 4 | 2 | 0 | ES (1), SSTI (3), UTI (1) | |
| TOB-CLI-ERY-TET (1) | tetK, ermC, ant(4′)-Ia | 1 | 0 | 0 | SSTI (1) | |
| TOB-CLI-ERY-MUP-TET (2) | tetK, ant(4′)-Ia, ermC, mupA1 | 1 | 2 | 0 | ES (1), SSTI (1) | |
| t1381 (2) | TET (1) | tetK | 0 | 1 | 0 | RTI (1) |
| CLI-ERY-CIP-TET (1) | tetK, ermC | 1 | 0 | 0 | RTI (1) | |
| t174 (1) | CIP-TET (1) | tetK | 1 | 0 | 0 | SSTI (1) |
| t693 (1) | TET (1) | tetK | 0 | 0 | 0 | SSTI (1) |
| t1784 (1) | ERY-TOB-TET (1) | tetK, ant(4′)-Ia, msrA | 1 | 0 | 1 | Blood (1) |
| t2207 (1) | CLI-ERY-MUP-CIP-TET (1) | tetK, ermC | 1 | 1 | 0 | RTI (1) |
| Phenotype | ermA | ermB | ermC | ermT | msrA | linA | linB | lsaB | vgaA | No of Isolates |
|---|---|---|---|---|---|---|---|---|---|---|
| ERYR-CLIS | − | − | − | − | + | − | − | − | − | 19 |
| ERYR-cCLIR | − | − | + | − | + | − | − | − | − | 5 |
| − | − | + | − | − | − | − | − | − | 3 | |
| − | + | + | − | − | − | − | − | − | 2 | |
| − | + | − | − | + | − | − | − | − | 2 | |
| + | + | − | − | + | + | − | + | − | 1 | |
| − | + | − | − | + | − | − | + | − | 1 | |
| − | + | + | − | − | − | − | + | − | 1 | |
| − | − | + | − | + | + | − | − | − | 1 | |
| ERYR-iCLIR | − | − | + | − | − | − | − | − | − | 25 |
| − | − | + | − | + | − | − | − | − | 4 | |
| − | − | + | − | + | + | − | − | − | 1 | |
| − | + | − | − | + | − | − | − | − | 1 | |
| − | + | − | − | + | − | − | + | − | 1 | |
| − | − | − | − | − | − | − | − | − | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceballos, S.; Lozano, C.; Aspiroz, C.; Ruiz-Ripa, L.; Eguizábal, P.; Campaña-Burguet, A.; Cercenado, E.; López-Calleja, A.I.; Castillo, J.; Azcona-Gutiérrez, J.M.; et al. Beyond CC398: Characterisation of Other Tetracycline and Methicillin-Resistant Staphylococcus aureus Genetic Lineages Circulating in Spanish Hospitals. Pathogens 2022, 11, 307. https://doi.org/10.3390/pathogens11030307
Ceballos S, Lozano C, Aspiroz C, Ruiz-Ripa L, Eguizábal P, Campaña-Burguet A, Cercenado E, López-Calleja AI, Castillo J, Azcona-Gutiérrez JM, et al. Beyond CC398: Characterisation of Other Tetracycline and Methicillin-Resistant Staphylococcus aureus Genetic Lineages Circulating in Spanish Hospitals. Pathogens. 2022; 11(3):307. https://doi.org/10.3390/pathogens11030307
Chicago/Turabian StyleCeballos, Sara, Carmen Lozano, Carmen Aspiroz, Laura Ruiz-Ripa, Paula Eguizábal, Allelen Campaña-Burguet, Emilia Cercenado, Ana Isabel López-Calleja, Javier Castillo, Jose Manuel Azcona-Gutiérrez, and et al. 2022. "Beyond CC398: Characterisation of Other Tetracycline and Methicillin-Resistant Staphylococcus aureus Genetic Lineages Circulating in Spanish Hospitals" Pathogens 11, no. 3: 307. https://doi.org/10.3390/pathogens11030307