
The C-Terminal Domain of Staphylococcus aureus Zinc Transport Protein AdcA Binds Plasminogen and Factor H In Vitro


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents, Purified Proteins, and Antibodies
2.2. Cloning, Expression, and Purification of Recombinant AdcA (Full-Length Protein) and AdcA Fragments
2.3. Antiserum against Recombinant AdcA
2.4. Binding of AdcA to Plasminogen
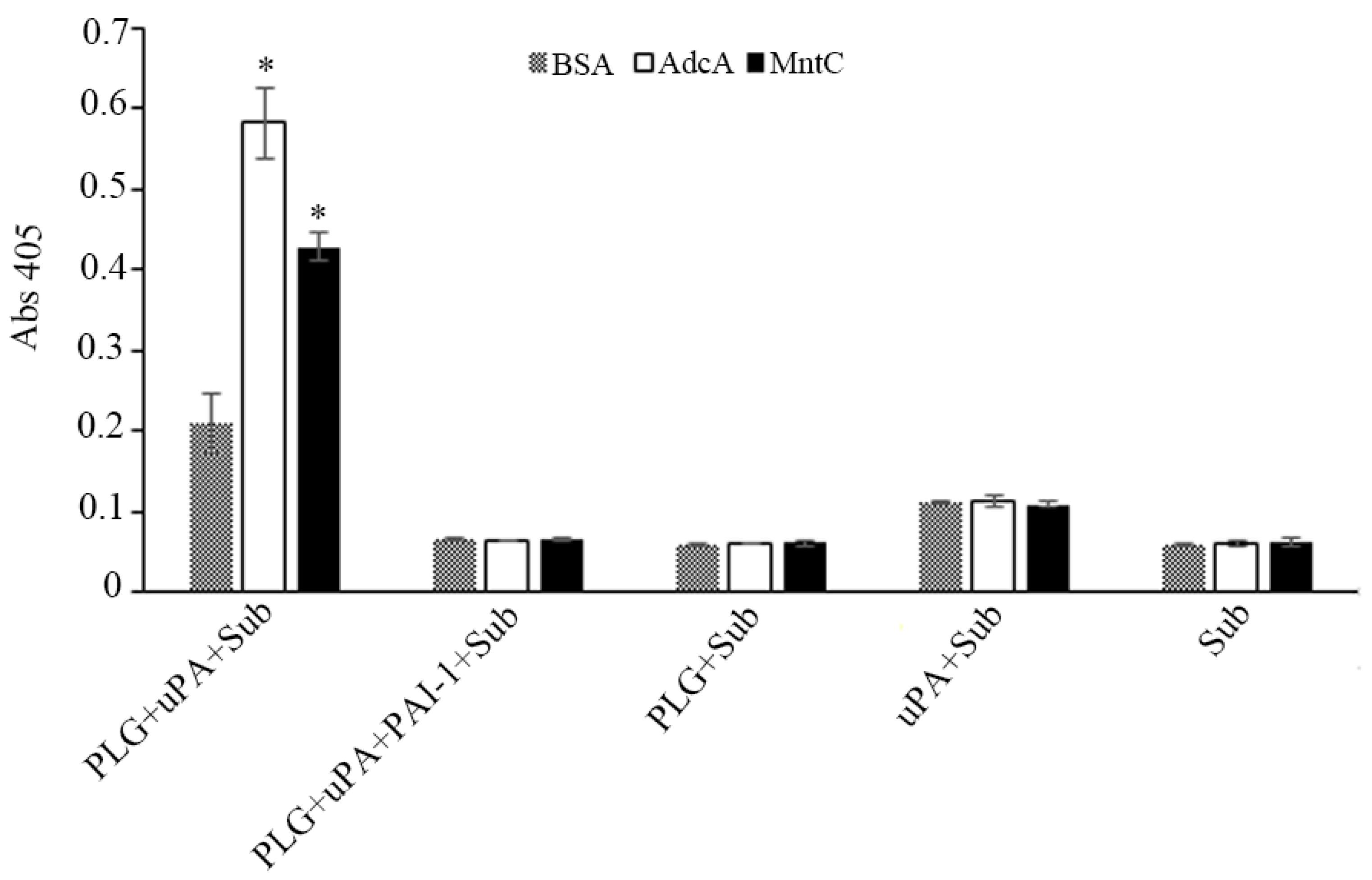
2.5. Effects of Ionic Strength and the Role of Lysines in AdcA–Plasminogen Interactions
2.6. Plasminogen Activation
2.7. Degradation of Fibrinogen and Vitronectin by Plasmin Bound to AdcA
2.8. Far Western Blot
2.9. Competition Assays
3. Results
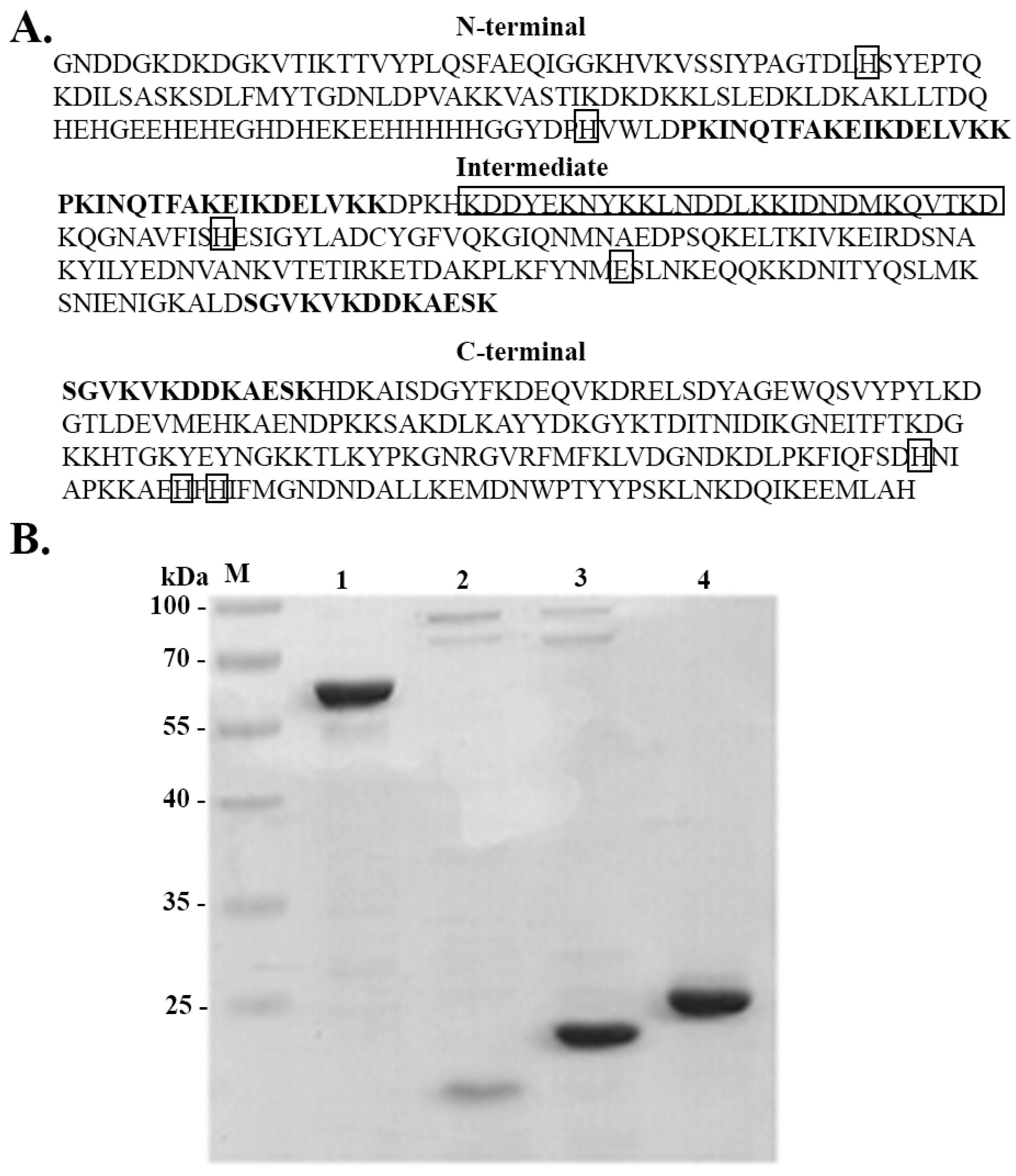
3.1. AdcA Sequence Analysis
3.2. Expression and Purification of Full-Length AdcA and Its Fragments
3.3. AdcA Interacts with Plasminogen
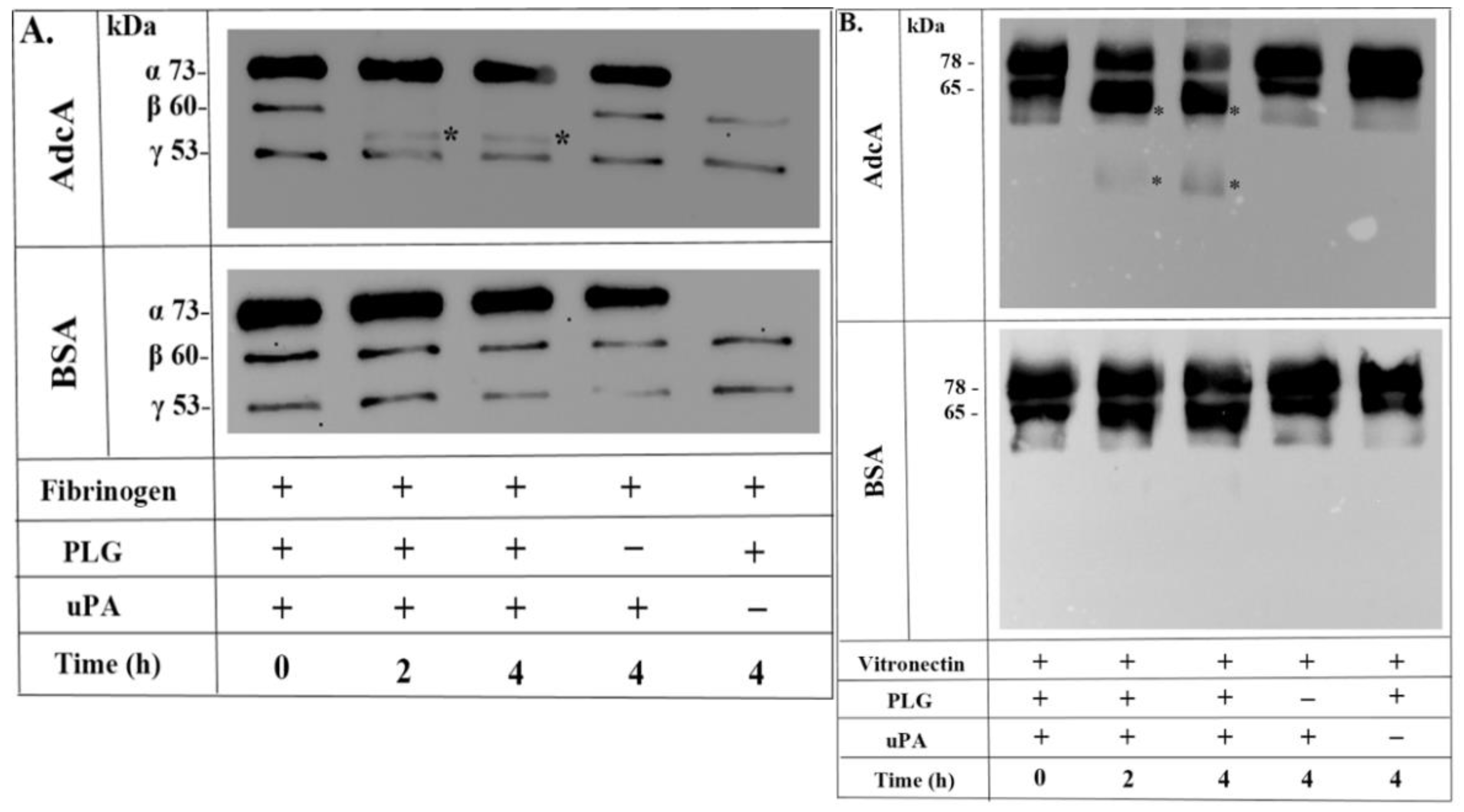
3.4. AdcA-Bound Plasminogen Is Activated to Plasmin
3.5. Plasmin Bound to AdcA Cleaves Fibrinogen and Vitronectin
3.6. AdcA Interacts with Plasminogen and FH through Its C-Terminal Domain
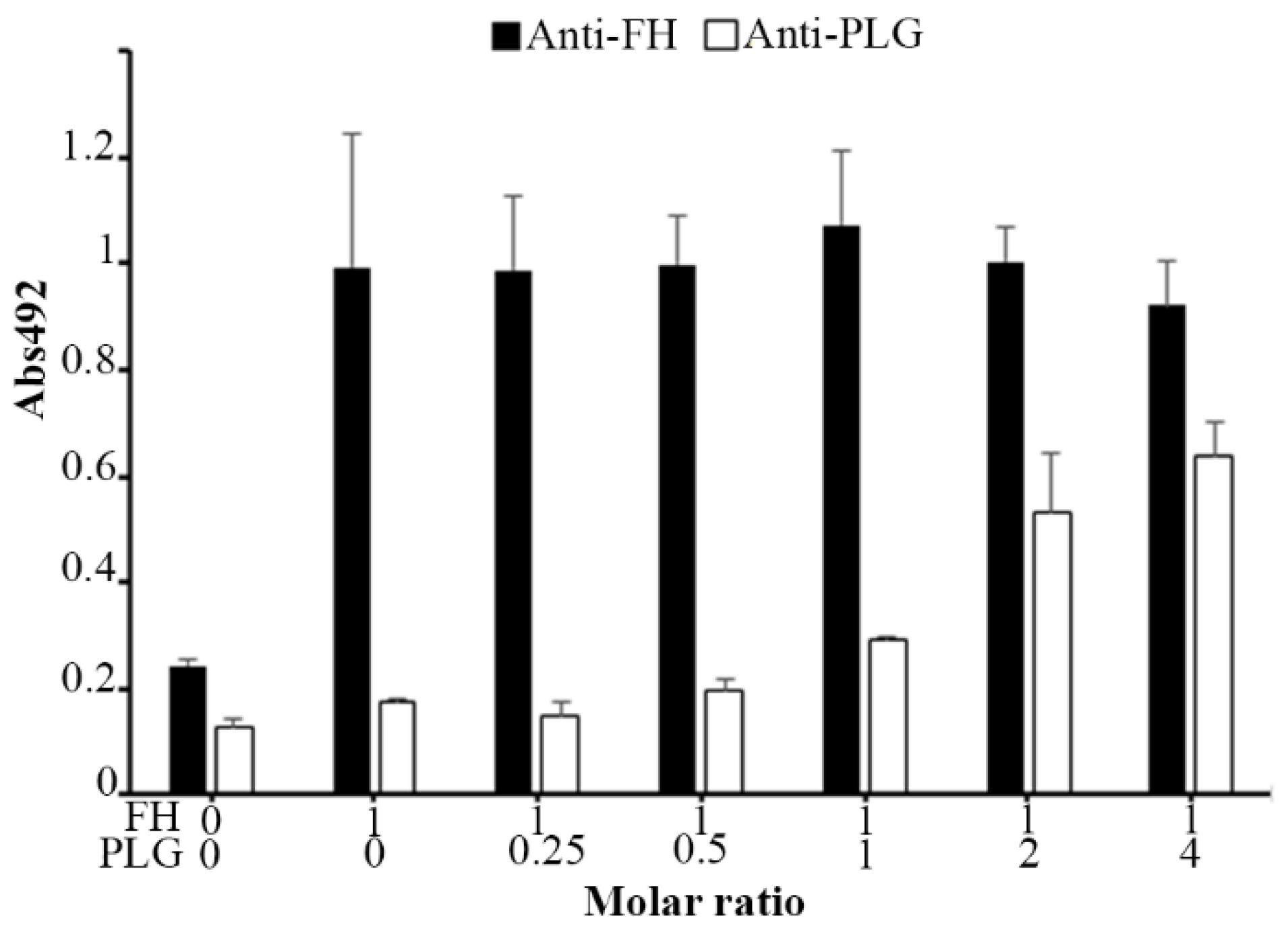
3.7. Plasminogen and FH Do Not Share Binding Sites on AdcA
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cassat, J.E.; Skaar, E.P. Metal ion acquisition in Staphylococcus aureus: Overcoming nutritional immunity. Semin. Immunopathol. 2012, 34, 215–235. [Google Scholar] [CrossRef] [Green Version]
- Shafeeq, S.; Kuipers, O.P.; Kloosterman, T.G. The role of zinc in the interplay between pathogenic streptococci and their hosts. Mol. Microbiol. 2013, 88, 1047–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Zinc through the Three Domains of Life. Proteome Res. 2006, 5, 3173–3178. [Google Scholar] [CrossRef]
- Formosa-Dague, C.; Speziale, P.; Foster, T.J.; Geoghegan, J.A.; Dufrêne, Y.F. Zinc-dependent mechanical properties of Staphylococcus aureus biofilm-forming surface protein SasG. Proc. Natl. Acad. Sci. USA 2016, 113, 410–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Terwilliger, A.; Maresso, A.W. Iron and zinc exploitation during bacterial pathogenesis. Metallomics 2015, 7, 1541–1554. [Google Scholar] [CrossRef] [Green Version]
- Cuajungco, M.P.; Ramirez, M.S.; Marcelo, E.; Tolmasky, M.E. Zinc: Multidimensional Effects on Living Organisms. Biomedicines 2021, 9, 208. [Google Scholar] [CrossRef] [PubMed]
- Capdevila, D.A.; Wang, J.; Giedroc, D.P. Bacterial Strategies to Maintain Zinc Metallostasis at the Host-Pathogen Interface. J. Biol. Chem. 2016, 291, 20858–20868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghssein, G.; Brutesco, C.; Ouerdane, L.; Fojcik, C.; Izaute, A.; Wang, S.; Hajja, C.; Lobinski, R.; Lemaire, D.; Richaud, P.; et al. Biosynthesis of a broad-spectrum nicotianamine-like metallophore in Staphylococcus aureus. Science 2016, 352, 1105–1109. [Google Scholar] [CrossRef]
- Grim, K.P.; San Francisco, B.; Radin, J.N.; Brazel, E.B.; Kelliher, J.L.; Solórzano, P.K.P.; Kim, P.C.; McDevitt, C.A.; Kehl-Fie, T.E. The metallophore staphylopine enables Staphylococcus aureus to compete with the host for zinc and overcome nutritional immunity. mBio 2017, 8, e01281-17. [Google Scholar] [CrossRef] [Green Version]
- Janulczyk, R.; Pallon, J.; Bjorck, L. Identification and characterization of a Streptococcus pyogenes ABC transporter with multiple specificity for metal cations. Mol. Microbiol. 1999, 34, 596–606. [Google Scholar] [CrossRef] [Green Version]
- Klein, J.S.; Lewinson, O. Bacterial ATP-driven transporters of transition metals: Physiological roles, mechanisms of action, and roles in bacterial virulence. Metallomics 2011, 3, 1098–1108. [Google Scholar] [CrossRef]
- Palmer, L.D.; Skaar, E.P. Transition Metals and Virulence in Bacteria. Annu. Rev. Genet. 2016, 23, 67–91. [Google Scholar] [CrossRef] [Green Version]
- Petrarca, P.; Ammendola, S.; Pasquali, P.; Battistoni, A. The Zur-regulated ZinT protein is na auxiliary component of the high-affinity ZnuABC zinc transporter that facilitates metal recruitment during severe zinc shortage. J. Bacteriol. 2010, 192, 1553–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panina, E.M.; Mironov, A.A.; Gelfand, M.S. Comparative genomics of bacterial zinc regulons: Enhanced ion transport, pathogenesis, and rearrangement of ribosomal proteins. Proc. Natl. Acad. Sci. USA 2003, 100, 9912–9917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, K.; Li, N.; Wang, H.; Cao, X.; He, J.; Zhang, B.; He, Q.-Y.; Zhang, G.; Sun, X. Two zinc-binding domains in the transporter AdcA from Streptococcus pyogenes facilitate high-affinity binding and fast transport of zinc. J. Biol. Chem. 2018, 293, 6075–6089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, A.S.; Scully, I.L.; Timofeyeva, Y.; Murphy, E.; McNeil, L.K.; Mininni, T.; Nuñez, L.; Carriere, M.; Singer, C.; Dilts, D.A.; et al. Staphylococcus aureus manganese transport protein C is a highly conserved cell surface protein that elicits protective immunity against S. aureus and Staphylococcus epidermidis. J. Infect. 2012, 205, 1688–1696. [Google Scholar] [CrossRef]
- Salazar, N.; Castiblanco-Valencia, M.M.; da Silva, L.B.; de Castro, Í.A.; Monaris, D.; Masuda, H.P.; Barbosa, A.S.; Arêas, A.P.M. Staphylococcus aureus manganese transport protein C (MntC) is an extracellular matrix-and plasminogen-binding protein. PLoS ONE 2014, 9, e112730. [Google Scholar] [CrossRef] [Green Version]
- Ramos, C.R.; Abreu, P.A.; Nascimento, A.L.; Ho, P.L. A high-copy T7 Escherichia coli expression vector for the production of recombinant proteins with a minimal N-terminal His-tagged fusion peptide. Braz. J. Med. Biol. 2004, 37, 1103e9. [Google Scholar] [CrossRef] [Green Version]
- Castiblanco-Valencia, M.M.; Fraga, T.R.; Pagotto, A.H.; de Toledo Serrano, S.M.; Abreu, P.A.E. Plasmin cleaves fibrinogen and the human complement proteins C3b and C5 in the presence of Leptospira interrogans proteins: A new role of LigA and LigB in invasion and complement immune evasion. Immunobiology 2016, 221, 679–689. [Google Scholar] [CrossRef]
- Castiblanco-Valencia, M.M.; Fraga, T.R.; Silva, L.B.; Monaris, D.; Abreu, P.A.; Strobel, S.; Józsi, M.; Isaac, L.; Barbosa, A.S. Leptospiral immunoglobulin-like proteins interact with human complement regulators factor H, FHL-1, FHR-1, and C4BP. J. Infect. Dis. 2012, 205, 995–1004. [Google Scholar] [CrossRef]
- Fulde, M.; Steinert, M.; Bergmann, S. Interaction of streptococcal plasminogen binding proteins with the host fibrinolytic system. Front. Cell. Infect. Microbiol. 2013, 3, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danø, K.; Andreasen, P.A.; Grondahl-Hansen, J.; Kristensen, P.; Nielsen, L.S.; Skriver, L. Plasminogen activators, tissue degradation, and cancer. Adv. Cancer Res. 1985, 44, 139–266. [Google Scholar] [PubMed]
- Wu, G.; Quek, A.J.; Caradoc-Davies, T.T.; Ekkel, S.M.; Mazzitelli, B.; Whisstock, J.C.; Law, R.H.P. Structural studies of plasmin inhibition. Biochem. Soc. Trans. 2019, 47, 541–557. [Google Scholar] [CrossRef]
- Barber-Zucker, S.; Shaanan, B.; Zarivach, R. Transition metal binding selectivity in proteins and its correlation with the phylogenomic classification of the cation diffusion facilitator protein family. Sci. Rep. 2017, 7, 16381–16392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roche, F.M.; Massey, R.; Peacock, S.J.; Day, N.P.; Visai, L.; Speziale, P.; Lam, A.; Pallen, M.; Foster, T.J. Characterization of novel LPXTG-containing proteins of Staphylococcus aureus identified from genome sequences. Microbiology 2003, 149, 643–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsompanidou, E.; Denham, E.L.; Sibbald, M.J.; Yang, X.M.; Seinen, J.; Friedrich, A.W.; Buist, G.; van Dijl, J.M. The sortase A substrates FnbpA, FnbpB, ClfA and ClfB antagonize colony spreading of Staphylococcus aureus. PLoS ONE 2012, 7, e44646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazmanian, S.K.; Liu, G.; Ton-That, H.; Schneewind, O. Staphylococcus aureus sortase, an enzyme that anchors surface proteins to the cell wall. Science 1999, 285, 760–763. [Google Scholar] [CrossRef]
- Josse, J.; Laurent, F.; Diot, A. Staphylococcal Adhesion and Host Cell Invasion: Fibronectin-Binding and Other Mechanisms. Front. Microbiol. 2017, 8, 2433. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.; Ko, Y.P.; Liang, X.; Ross, C.L.; Liu, Q.; Murray, B.E.; Höök, M. Collagen-binding microbial surface components recognizing adhesive matrix molecule (MSCRAMM) of Gram-positive bacteria inhibit complement activation via the classical pathway. J. Biol. Chem. 2013, 288, 20520–20531. [Google Scholar] [CrossRef] [Green Version]
- Fulde, M.; Rohde, M.; Hitzmann, A.; Preissner, K.T.; Nitsche-Schmitz, D.P.; Nerlich, A.; Chhatwal, G.S.; Bergmann, S. SCM, a novel M-like protein from Streptococcus canis, binds (mini)-plasminogen with high affinity and facilitates bacterial transmigration. Biochem. J. 2011, 434, 523–535. [Google Scholar] [CrossRef] [Green Version]
- Peetermans, M.; Vanassche, T.; Liesenborghs, L.; Claes, J.; Velde, G.V.; Kwiecinksi, J.; Jin, T.; De Geest, B.; Hoylaerts, M.F.; Lijnen, R.H.; et al. Plasminogen activation by staphylokinase enhances local spreading of S. aureus in skin infections. BMC Microbiol. 2014, 14, 310–321. [Google Scholar] [CrossRef] [Green Version]
- Hemmadi, V.; Biswas, M. An overview of moonlighting proteins in Staphylococcus aureus infection. Arch. Microbiol. 2021, 203, 481–498. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, C.; Wenzel, E.; Heiden, M. Plasminogen: A brief introduction into its biochemistry and function. Haemostasis 1988, 18, 7–13. [Google Scholar] [CrossRef]
- Pancholi, V.; Fischetti, V.A. Alpha-enolase, a novel strong plasmin(ogen) binding protein on the surface of pathogenic streptococci. J. Biol. Chem. 1998, 273, 14503–14515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, S.; Wild, D.; Diekmann, O.; Frank, R.; Bracht, D.; Chhatwal, G.S.; Hammerschmidt, S. Identification of a novel plasmin(ogen)-binding motif in surface displayed a-enolase of Streptococcus pneumoniae. Mol. Microbiol. 2003, 49, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, R.H.; McVernon, J.; Geard, N. Implications of asymptomatic carriers for infectious disease transmission and control. R. Soc. Open Sci. 2018, 5, 172341. [Google Scholar] [CrossRef] [Green Version]
- Kost, C.; Benner, K.; Stockmann, A.; Linder, D.; Preissner, K.T. Limited plasmin proteolysis of vitronectin: Characterization of the adhesion protein as morpho-regulatory and angiostatin-binding factor. Eur. J. Biochem. 1996, 236, 682–688. [Google Scholar] [CrossRef]
- Zipfel, P.F.; Hallström, T.; Riesbeck, K. Human complement control and complement evasion by pathogenic microbes–tipping the balance. Mol. Immun. 2013, 56, 152–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burman, J.D.; Leung, E.; Atkins, K.L.; O’Seaghdha, M.N.; Lango, L. Interaction of human complement with Sbi, a staphylococcal immunoglobulin-binding protein indications of a novel mechanism of complement evasion by Staphylococcus aureus. J. Biol. Chem. 2008, 283, 17579–17593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, B.L.; Ramyar, K.X.; Ricklin, D.; Lambris, J.D.; Geisbrecht, B.V. Advances in understanding the structure, function, and mechanism of the SCIN and Efb families of Staphylococcal immune evasion proteins. In Current Topics in Innate Immunity II; Springer: New York, NY, USA, 2012; pp. 113–133. [Google Scholar] [CrossRef] [Green Version]
- Sharp, J.A.; Echague, C.G.; Hair, P.S.; Ward, M.D.; Nyalwidhe, J.O.; Geoghegan, J.A.; Foster, T.J.; Cunnion, K.M. Staphylococcus aureus surface protein SdrE binds complement regulator factor H as an immune evasion tactic. PLoS ONE 2012, 7, e38407. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Origin | Molecular Weight (kDa) | UniProtKBs |
|---|---|---|---|
| AdcA (S. aureus) | This study | 59.1 | A0A0D3QA93 |
| AdcA N-terminal fragment (S. aureus) | This study | 19 | - |
| AdcA intermediate fragment (S. aureus) | This study | 22 | - |
| AdcA C-terminal fragment (S. aureus) | This study | 24 | - |
| MntC | Salazar et al., 2014 | 35 | Q8VQS9 |
| LigA C-terminal fragment | Castiblanco-Valencia et al., 2016 | 63 | - |
| LigB C-terminal fragment | Castiblanco-Valencia et al., 2016 | 56 | - |
| Human plasminogen | Commercial (Sigma-Aldrich) | 92 | P00747 |
| Commercial (Sigma-Aldrich) | α-73 | α-P02671 | |
| Human fibrinogen | β-60 | β-P02675 | |
| γ-53 | γ-P02679 | ||
| Human vitronectin | Commercial (Sigma-Aldrich) | 75/65 | P04004 |
| Human complement factor H | Commercial (Complement Technology) | 155 | Q03591 |
| Urokinase-type plasminogen activator | Commercial (Sigma-Aldrich) | 46.9 | P00749 |
| Plasminogen activator inhibitor 1 | Commercial (Sigma-Aldrich) | 43.4 | P05121 |
| Goat anti-human factor H | Commercial (Quidel) | - | - |
| Rabbit anti-human fibrinogen | Commercial (Calbiochem) | - | - |
| Rabbit anti-human plasminogen | Commercial (Sigma-Aldrich) | - | - |
| Rabbit anti-human vitronectin | Commercial (Complement Technology) | - | - |
| Peroxidase-conjugated anti-goat immunoglobulin G | Commercial (Sigma-Aldrich) | - | - |
| Peroxidase-conjugated goat anti-rabbit immunoglobulin G | Commercial (Sigma-Aldrich) | - | - |
| Bovine serum albumin | Commercial (Sigma-Aldrich) | 69.2 | P02769 |
| Full-length AdcA | F: 5′-GGATCCGGGAATGATGATGGAAAAGATAAAGATGGC-3′ |
| R: 5′-CTGCAGTTAATGCGCTAACATTTCTTCTTTG-3′ | |
| N-terminal fragment | F: 5′-GGATCCGGGAATGATGATGGAAAAGATAAAGATGGC-3′ |
| R: 5′-CCATGGTTATTTTTTCACTAATTCATCTTTAATTTCTTTAGCG-3′ | |
| Intermediate fragment | F: 5′-GGATCCCCTAAAATTAACCAAACTTTCGCTAAAG-3′ |
| R: 5′-CCATGGTTATTTACTTTCAGCTTTGTCGTCTTTCAC-3′ | |
| C-terminal fragment | F: 5′-GGATCCAGTGGTGTTAAAGTGAAAGACGAC-3′ |
| R: 5′-CCATGGTTAATGCGCTAACATTTCTTCTTTG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salazar, N.; Yamamoto, B.B.; Souza, M.C.L.d.; Silva, L.B.d.; Arêas, A.P.M.; Barbosa, A.S. The C-Terminal Domain of Staphylococcus aureus Zinc Transport Protein AdcA Binds Plasminogen and Factor H In Vitro. Pathogens 2022, 11, 240. https://doi.org/10.3390/pathogens11020240
Salazar N, Yamamoto BB, Souza MCLd, Silva LBd, Arêas APM, Barbosa AS. The C-Terminal Domain of Staphylococcus aureus Zinc Transport Protein AdcA Binds Plasminogen and Factor H In Vitro. Pathogens. 2022; 11(2):240. https://doi.org/10.3390/pathogens11020240
Chicago/Turabian StyleSalazar, Natália, Bruno Bernardi Yamamoto, Matilde Costa Lima de Souza, Ludmila Bezerra da Silva, Ana Paula Mattos Arêas, and Angela Silva Barbosa. 2022. "The C-Terminal Domain of Staphylococcus aureus Zinc Transport Protein AdcA Binds Plasminogen and Factor H In Vitro" Pathogens 11, no. 2: 240. https://doi.org/10.3390/pathogens11020240