
Impact of Sainfoin (Onobrychis viciifolia) Pellets on Parasitological Status, Antibody Responses, and Antioxidant Parameters in Lambs Infected with Haemonchus contortus

 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Results
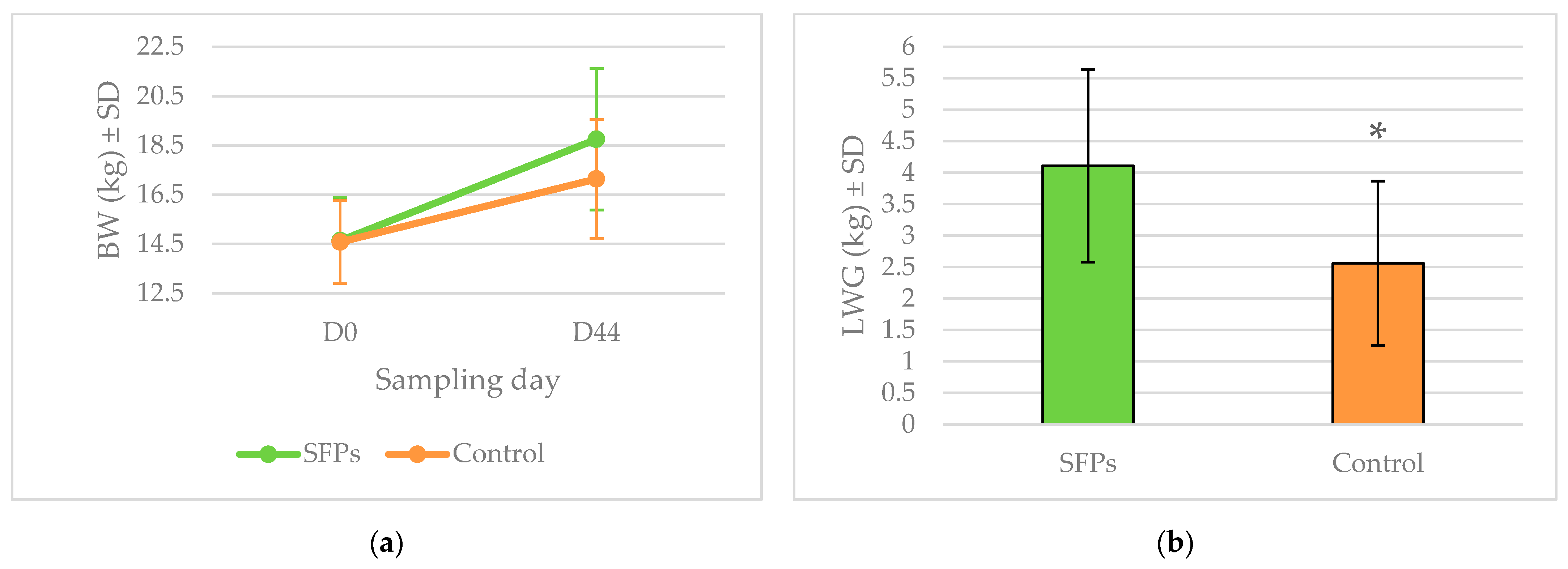
2.1. Fecal Egg Count (FEC) and Weight Gains
2.2. Egg Hatch Test (EHT)
2.3. Antibody Responses
2.4. Antioxidant Status
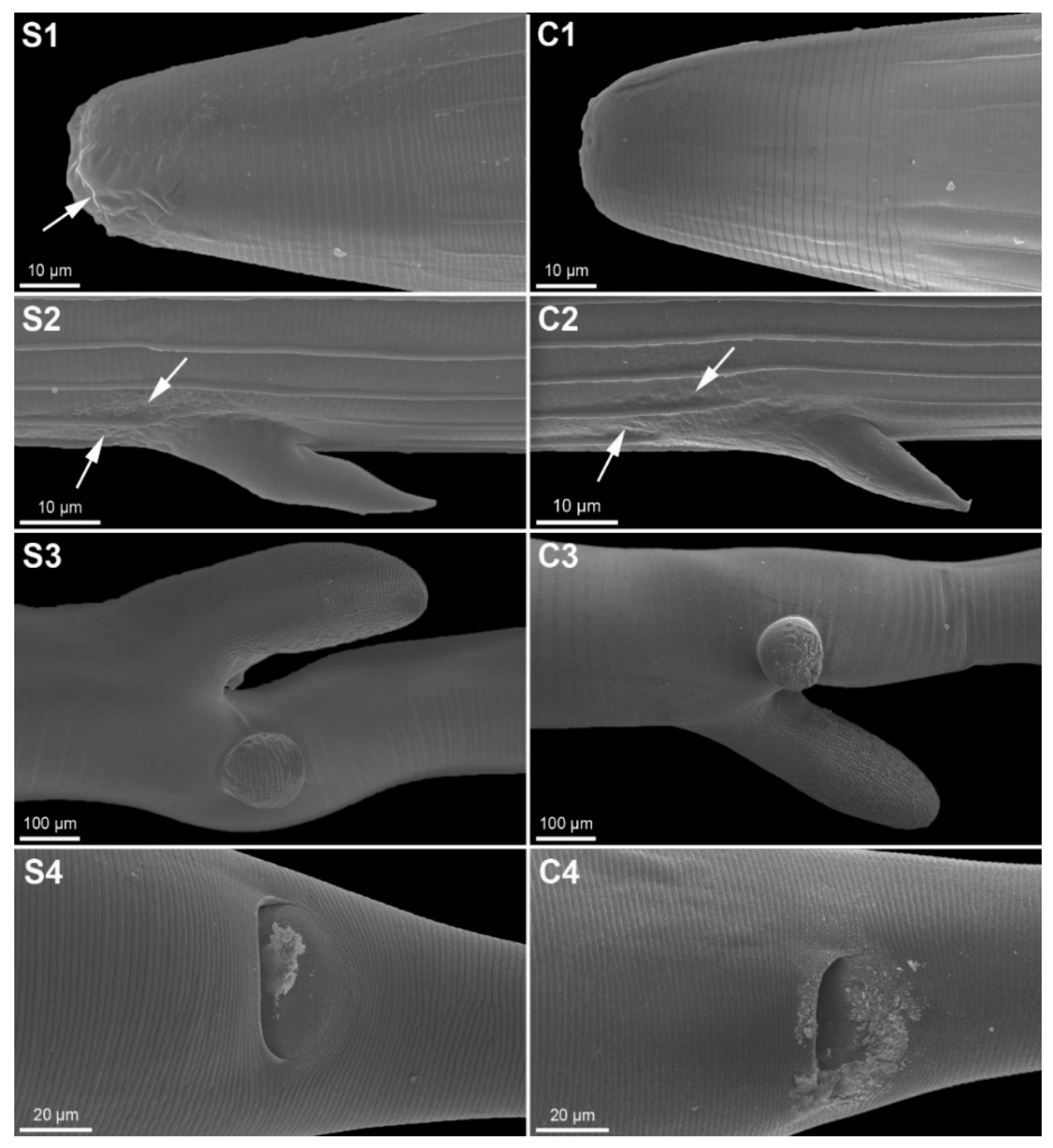
2.5. Scanning Electron Microscopy (SEM)
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Experimental Design, Diets, and Experimental Infection
4.3. Chemical Tests of Sainfoin Pellets (SFPs)
4.4. Fecal Egg Count and Animal Weighing
4.5. Counts of Adult Worms
4.6. Egg Hatch Test
4.7. Antibody Responses
4.8. Antioxidant Parameters
4.9. Scanning Electron Microscopy (SEM)
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Papadopoulos, E. Anthelmintic resistance in sheep nematodes. Small Rumin. Res. 2008, 76, 99–103. [Google Scholar] [CrossRef]
- Waller, P.J. From discovery to development: Current industry perspectives for the development of novel methods of helminth control in livestock. Vet. Parasitol. 2006, 139, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Besier, R.B.; Kahn, L.P.; Sargison, N.D.; Van Wyk, J.A. The pathophysiology, ecology and epidemiology of Haemonchus contortus infection in small ruminants. Adv. Parasitol. 2016, 93, 95–143. [Google Scholar] [CrossRef] [PubMed]
- Balic, A.; Bowles, V.M.; Meeusen, E.N. The immunobiology of gastrointestinal nematode infections in ruminants. Adv. Parasitol. 2000, 45, 181–241. [Google Scholar] [CrossRef]
- Colditz, I.G. Six costs of immunity to gastrointestinal nematode infections. Parasite Immunol. 2008, 30, 63–70. [Google Scholar] [CrossRef]
- Coop, R.L.; Holmes, P.H. Nutrition and parasite interaction. Int. J. Parasitol. 1996, 26, 951–962. [Google Scholar] [CrossRef]
- Hoste, H.; Torres-Acosta, J.F.; Quijada, J.; Chan-Perez, I.; Dakheel, M.M.; Kommuru, D.S.; Mueller-Harvey, I.; Terrill, T.H. Interactions between nutrition and infections with Haemonchus contortus and related gastrointestinal nematodes in small ruminants. Adv. Parasitol. 2016, 93, 239–351. [Google Scholar] [CrossRef]
- Williams, A.R.; Klaver, E.J.; Laan, L.C.; Ramsay, A.; Fryganas, C.; Difborg, R.; Kringel, H.; Reed, J.D.; Mueller-Harvey, I.; Skov, S.; et al. Co-operative suppression of inflammatory responses in human dendritic cells by plant proanthocyanidins and products from the parasitic nematode Trichuris suis. Immunology 2017, 150, 312–328. [Google Scholar] [CrossRef] [Green Version]
- Tedeschi, L.O.; Muir, J.P.; Naumann, H.D.; Norris, A.B.; Ramírez-Restrepo, C.A.; Mertens-Talcott, S.U. Nutritional aspects of ecologically relevant phytochemicals in ruminant production. Front. Vet. Sci. 2021, 8, 628445. [Google Scholar] [CrossRef]
- Machado, V.; Da Silva, A.S.; Schafer, A.S.; Aires, A.R.; Tonin, A.A.; Oliveira, C.B.; Hermes, C.L.; Almeida, T.C.; Moresco, R.N.; Stefani, L.M.; et al. Relationship between oxidative stress and pathological findings in abomasum of infected lambs by Haemonchus contortus. Pathol. Res. Pract. 2014, 210, 812–817. [Google Scholar] [CrossRef]
- Čobanová, K.; Váradyová, Z.; Grešáková, Ľ.; Kucková, K.; Mravčáková, D.; Várady, M. Does herbal and/or zinc dietary supplementation improve the antioxidant and mineral status of lambs with parasite infection? Antioxidants 2020, 9, 1172. [Google Scholar] [CrossRef] [PubMed]
- Mravčáková, D.; Sobczak-Filipiak, M.; Váradyová, Z.; Kucková, K.; Čobanová, K.; Maršík, P.; Tauchen, J.; Vadlejch, J.; Mickiewicz, M.; Kaba, J.; et al. Effect of Artemisia absinthium and Malva sylvestris on antioxidant parameters and abomasal histopathology in lambs experimentally infected with Haemonchus contortus. Animals 2021, 11, 462. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The immunomodulatory and anti-inflammatory role of polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paolini, V.; De La Farge, F.; Prevot, F.; Dorchies, P.; Hoste, H. Effects of the repeated distribution of sainfoin hay on the resistance and the resilience of goats naturally infected with gastrointestinal nematodes. Vet. Parasitol. 2005, 127, 277–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, A.R.; Fryganas, C.; Ramsay, A.; Mueller-Harvey, I.; Thamsborg, S.M. Direct anthelmintic effects of condensed tannins from diverse plant sources against Ascaris suum. PLoS ONE 2014, 9, e97053. [Google Scholar] [CrossRef] [Green Version]
- Hoste, H.; Torres-Acosta, J.F.; Sandoval-Castro, C.A.; Mueller-Harvey, I.; Sotiraki, S.; Louvandini, H.; Thamsborg, S.M.; Terrill, T.H. Tannin containing legumes as a model for nutraceuticals against digestive parasites in livestock. Vet. Parasitol. 2015, 212, 5–17. [Google Scholar] [CrossRef]
- Spiegler, V.; Liebau, E.; Hensel, A. Medicinal plant extracts and plant-derived polyphenols with anthelmintic activity against intestinal nematodes. Nat. Prod. Rep. 2017, 34, 627–643. [Google Scholar] [CrossRef]
- Gaudin, E.; Simon, M.; Quijada, J.; Schelcher, F.; Sutra, J.-F.; Lespine, A.; Hoste, H. Efficacy of sainfoin (Onobrychis viciifolia) pellets against multi resistant Haemonchus contortus and interaction with oral ivermectin: Implications for on-farm control. Vet. Parasitol. 2016, 227, 122–129. [Google Scholar] [CrossRef]
- Singh, S.; Pathak, A.K.; Sharma, R.K.; Khan, M. Effect of tanniferous leaf meal based multi-nutrient blocks on feed intake, hematological profile, immune response, and body weight changes in Haemonchus contortus infected goats. Vet. World 2015, 8, 572–579. [Google Scholar] [CrossRef] [Green Version]
- Tufarelli, V.; Casalino, E.; D’Alessandro, A.G.; Laudadio, V. Dietary phenolic compounds: Biochemistry, metabolism and significance in animal and human health. Curr. Drug Metab. 2017, 18, 905–913. [Google Scholar] [CrossRef]
- Váradyová, Z.; Kišidayová, S.; Čobanová, K.; Grešáková, Ľ.; Babják, M.; Königová, A.; Urda Dolinská, M.; Várady, M. The impact of a mixture of medicinal herbs on ruminal fermentation, parasitological status and hematological parameters of the lambs experimentally infected with Haemonchus contortus. Small Rumin. Res. 2017, 151, 124–132. [Google Scholar] [CrossRef]
- Váradyová, Z.; Mravčáková, D.; Babják, M.; Bryszak, M.; Grešáková, Ľ.; Čobanová, K.; Kišidayová, S.; Plachá, I.; Königová, A.; Cieslak, A.; et al. Effects of herbal nutraceuticals and/or zinc against Haemonchus contortus in lambs experimentally infected. BMC Vet. Res. 2018, 14, 78. [Google Scholar] [CrossRef] [PubMed]
- Mravčáková, D.; Váradyová, Z.; Kopčáková, A.; Čobanová, K.; Grešáková, Ľ.; Kišidayová, S.; Babják, M.; Urda Dolinská, M.; Dvorožňáková, E.; Königová, A.; et al. Natural chemotherapeutic alternatives for controlling of haemonchosis in sheep. BMC Vet. Res. 2019, 15, 302. [Google Scholar] [CrossRef] [PubMed]
- Komáromyová, M.; Mravčáková, D.; Petrič, D.; Kucková, K.; Babják, M.; Dolinská, M.U.; Königová, A.; Maďarová, M.; Pruszyńska-Oszmałek, E.; Cieslak, A.; et al. Effects of medicinal plants and organic selenium against ovine haemonchosis. Animals 2021, 11, 1319. [Google Scholar] [CrossRef]
- Váradyová, Z.; Pisarčíková, J.; Babják, M.; Hodges, A.; Mravčáková, D.; Kišidayová, S.; Königová, A.; Vadlejch, J.; Várady, M. Ovicidal and larvicidal activity of extracts from medicinal-plants against Haemonchus contortus. Exp. Parasitol. 2018, 195, 71–77. [Google Scholar] [CrossRef]
- Mravčáková, D.; Komáromyová, M.; Babják, M.; Urda Dolinská, M.; Königová, A.; Petrič, D.; Čobanová, K.; Ślusarczyk, S.; Cieslak, A.; Várady, M.; et al. Anthelmintic activity of wormwood (Artemisia absinthium L.) and mallow (Malva sylvestris L.) against Haemonchus contortus in sheep. Animals 2020, 10, 219. [Google Scholar] [CrossRef] [Green Version]
- Hoste, H.; Martinez-Ortiz-De-Montellano, C.; Manolaraki, F.; Brunet, S.; Ojeda-Robertos, N.; Fourquaux, I.; Torres-Acosta, J.F.; Sandoval-Castro, C.A. Direct and indirect effects of bioactive tannin-rich tropical and temperate legumes against nematode infections. Vet. Parasitol. 2012, 186, 18–27. [Google Scholar] [CrossRef]
- Heckendorn, F.; Häring, D.A.; Maurer, V.; Zinsstag, J.; Langhans, W.; Hertzberg, H. Effect of sainfoin (Onobrychis viciifolia) silage and hay on established populations of Haemonchus contortus and Cooperia curticei in lambs. Vet. Parasitol. 2006, 142, 293–300. [Google Scholar] [CrossRef]
- Valderrábano, J.; Calvete, C.; Uriarte, J. Effect of feeding bioactive forages on infection and subsequent development of Haemonchus contortus in lamb faeces. Vet. Parasitol. 2010, 172, 89–94. [Google Scholar] [CrossRef]
- Desrues, O.; Peña-Espinoza, M.; Hansen, T.V.; Enemark, H.L.; Thamsborg, S.M. Anti-parasitic activity of pelleted sainfoin (Onobrychis viciifolia) against Ostertagia ostertagi and Cooperia oncophora in calves. Parasites Vectors 2016, 9, 329. [Google Scholar] [CrossRef] [Green Version]
- Athanasiadou, S.; Kyriazakis, F.; Jackson, F.; Coop, R.I. Direct anthelmintic effects of condensed tannins towards different gastrointestinal nematodes of sheep: In vitro and in vivo studies. Vet. Parasitol. 2001, 99, 205–219. [Google Scholar] [CrossRef]
- Paolini, V.; Fouraste, I.; Hoste, H. In vitro effects of three woody plant and sainfoin extracts on 3rd-stage larvae and adult worms of three gastrointestinal nematodes. Parasitology 2004, 129, 69–77. [Google Scholar] [CrossRef]
- Hoste, H.; Jackson, F.; Athanasiadou, S.; Thamsborg, S.M.; Hoskin, S.O. The effects of tannin-rich plants on parasitic nematodes in ruminants. Trends Parasitol. 2006, 22, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Desrues, O.; Fryganas, C.; Ropiak, H.M.; Mueller-Harvey, I.; Enemark, H.L.; Thamsborg, S.M. Impact of chemical structure of flavanol monomers and condensed tannins on in vitro anthelmintic activity against bovine nematodes. Parasitology 2016, 143, 444–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klongsiriwet, C.; Quijada, J.; Williams, A.R.; Mueller-Harvey, I.; Williamson, E.M.; Hoste, H. Synergistic inhibition of Haemonchus contortus exsheathment by flavonoid monomers and condensed tannins. Int. J. Parasitol. Drugs Drug Resist. 2015, 5, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Barrau, E.; Fabre, N.; Fouraste, I.; Hoste, H. Effect of bioactive compounds from sainfoin (Onobrychis viciifolia Scop.) on the in vitro larval migration of Haemonchus contortus: Role of tannins and flavonol glycosides. Parasitology 2005, 131, 531–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Díaz, M.A.; Torres-Acosta, J.F.J.; Sandoval-Castro, C.A.; Hoste, H. Comparing the sensitivity of two in vitro assays to evaluate the anthelmintic activity of tropical tannin rich plant extracts against Haemonchus contortus. Vet. Parasitol. 2011, 181, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Quijada, J.; Fryganas, C.; Ropiak, H.M.; Ramsay, A.; Mueller-Harvey, I.; Hoste, H. Anthelmintic activities against Haemonchus contortus or Trichostrongylus colubriformis from small ruminants are influenced by structural features of condensed tannins. J. Agric. Food Chem. 2015, 63, 6346–6354. [Google Scholar] [CrossRef] [Green Version]
- Ríos-De Álvarez, L.; Greer, A.; Jackson, F.; Athanasiadou, S.; Kyriazakis, I.; Huntley, J. The effect of dietary sainfoin (Onobrychis viciifolia) on local cellular responses to Trichostrongylus colubriformis in sheep. Parasitology 2008, 135, 1117–1124. [Google Scholar] [CrossRef] [Green Version]
- McRae, K.M.; Good, B.; Hanrahan, J.P.; Glynn, A.; O’Connell, M.J.; Keane, O.M. Response to Teladorsagia circumcincta infection in Scottish Blackface lambs with divergent phenotypes for nematode resistance. Vet. Parasitol. 2014, 206, 200–207. [Google Scholar] [CrossRef] [Green Version]
- De Cisneros, J.; Matthews, L.; Mair, C.; Stefan, T.; Stear, M. The transfer of IgA from mucus to plasma and the implications for diagnosis and control of nematode infections. Parasitology 2014, 141, 875–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doligalska, M.; Moskwa, B.; Stear, M.J. Relationships among peripheral eosinophilia, eosinophil peroxidase activity, interleukin-5 concentration and faecal nematode egg count during natural, mixed gastrointestinal nematode infection. Vet. Immunol. Immunopathol. 1999, 70, 299–308. [Google Scholar] [CrossRef]
- Jenvey, C.J.; Alenizi, D.; Almasi, F.; Cairns, C.; Holmes, A.; Sloan, S.; Stear, M.J. Bioinformatic analysis of eosinophil activity and its implications for model and target species. Parasitology 2020, 147, 393–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiedemann, M.; Voehringer, D. Immunomodulation and immune escape strategies of gastrointestinal helminths and schistosomes. Front. Immunol. 2020, 11, 2271. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Singh, S.; Prasad, S.M. Role of medicinal plants for health perspective: Special references to antioxidant potential. J. Chem. Biol. Ther. 2016, 1, 106. [Google Scholar] [CrossRef]
- Surai, P.F. Polyphenol compounds in the chicken/animal diet: From the past to the future. J. Anim. Physiol. Anim. Nutr. 2014, 98, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, W.L.C.; Andre, W.P.P.; Cavalcante, G.S.; de Araújo-Filho, J.V.; Santos, J.M.L.; Macedo, I.T.F.; de Melo, J.V.; de Morais, S.M.; Bevilaqua, C.M.L. Effects of Spigelia anthelmia decoction on sheep gastrointestinal nematodes. Small Rumin. Res. 2017, 153, 146–152. [Google Scholar] [CrossRef]
- Adamu, M.; Mukandiwa, L.; Awouafack, M.D.; Ahmed, A.S.; Eloff, J.N.; Naidoo, V. Ultrastructure changes induced by the phloroglucinol derivative agrimol G isolated from Leucosidea sericea in Haemonchus contortus. Exp. Parasitol. 2019, 207, 107780. [Google Scholar] [CrossRef] [PubMed]
- Cavalcante, G.S.; Morais, S.M.; André, W.P.P.; Araújo-Filho, J.V.; Muniz, C.R.; Rocha, L.O.; Ribeiro, W.L.C.; Rodrigues, A.L.M.; de Oliveira, L.M.B.; Bevilaqua, C.M.L.; et al. Chemical constituents of Calotropis procera latex and ultrastructural effects on Haemonchus contortus. Braz. J. Vet. Parasitol. 2020, 29, e001320. [Google Scholar] [CrossRef]
- Sambodo, P.; Prastowo, J.; Kurniasih, K.; Indarjulianto, S. In vitro potential anthelmintic activity of Biophytum petersianum on Haemonchus contortus. Vet. World 2018, 11, 1–4. [Google Scholar] [CrossRef]
- Brunet, S.; Fourquaux, I.; Hoste, H. Ultrastructural changes in the third-stage, infective larvae of ruminant nematodes treated with sainfoin (Onobrychis viciifolia) extract. Parasitol. Int. 2011, 60, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ortiz-de-Montellano, C.; Torres-Acosta, J.F.J.; Fourquaux, I.; Sandoval-Castro, C.A.; Hoste, H. Ultrastructural study of adult Haemonchus contortus exposed to polyphenol-rich materials under in vivo conditions in goats. Parasite 2019, 26, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Ortíz-de-Montellano, C.; Arroyo-López, C.; Fourquaux, I.; Torres-Acosta, J.F.; Sandoval-Castro, C.A.; Hoste, H. Scanning electron microscopy of Haemonchus contortus exposed to tannin-rich plants under in vivo and in vitro conditions. Exp. Parasitol. 2013, 133, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Kommuru, D.S.; Whitley, N.C.; Miller, J.E.; Mosjidis, J.A.; Burke, J.M.; Gujja, S.; Mechineni, A.; Terrill, T.H. Effect of Sericea lespedeza leaf meal pellets on adult female Haemonchus contortus in goats. Vet. Parasitol. 2015, 207, 170–175. [Google Scholar] [CrossRef]
- Andre, W.P.; Ribeiro, W.L.; Cavalcante, G.S.; dos Santos, J.M.; Macedo, I.T.; de Paula, H.C.; de Freitas, R.M.; de Morais, S.M.; de Melo, J.V.; Bevilaqua, C.M. Comparative efficacy and toxic effects of carvacryl acetate and carvacrol on sheep gastrointestinal nematodes and mice. Vet. Parasitol. 2016, 218, 52–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Commission (EC). Council Regulation (EC) 1099/2009 of 24 September 2009 on the Protection of Animals at the Time of Killing. Off. J. Eur. Union 2009, L 303, 1–30. [Google Scholar]
- Jaradat, N.; Hussen, F.; Al Ali, A. Preliminary phytochemical screening, quantitative estimation of total flavonoids, total phenols and antioxidant activity of Ephedra alata Decne. J. Mater. Environ. Sci. 2015, 6, 1771–1778. [Google Scholar]
- Proestos, C.; Lytoudi, K.; Mavromelanidou, O.K.; Zoumpoulakis, P.; Sinanoglou, V.J. Antioxidant capacity of selected plant extracts and their essential oils. Antioxidants 2013, 2, 11–22. [Google Scholar] [CrossRef]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar] [CrossRef]
- Coles, G.C.; Bauer, C.; Borgsteede, F.H.M.; Geerts, S.; Klei, T.R.; Taylor, M.A.; Waller, P.J. World Association for the Advancement of Veterinary Parasitology (W.A.A.V.P.) methods for the detection of anthelmintic resistance in nematodes of veterinary importance. Vet. Parasitol. 1992, 44, 35–44. [Google Scholar] [CrossRef]
- Amarante, A.F.T.; Bricarello, P.A.; Huntley, J.F.; Mazzolin, L.P.; Gomes, J.C. Relationship of abomasal histology and parasite-specific immunoglobulin A with the resistance to Haemonchus contortus infection in three breeds of sheep. Vet. Parasitol. 2005, 128, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Schallig, H.; Van Leeuwen, M.; Hendrikx, W. Immune responses of Texel sheep to excretory/secretory products of adult Haemonchus contortus. Parasitology 1994, 108, 351–357. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Concentration (mg/mL) | Hatching (%) | Ovicidal Effect (%) | p |
|---|---|---|---|
| 50 | 5 ± 7.42 | 95 | <0.001 |
| 25 | 4 ± 6.33 | 96 | <0.001 |
| 12.5 | 11 ± 14.47 | 89 | <0.001 |
| 6.25 | 26 ± 28.74 | 74 | <0.001 |
| 3.125 | 76 ± 12.25 | 24 | <0.001 |
| 1.563 | 88 ± 3.71 | 12 | <0.001 |
| 0 | 98 ± 1.095 | - | - |
| Day | SFPs | Control | SD | p | |||
|---|---|---|---|---|---|---|---|
| Treatment (T) | Time | T × Time | |||||
| IgG (OD) | 23 | 0.357 | 0.361 | 0.0773 | 0.611 | 0.837 | 0.987 |
| 30 | 0.397 | 0.374 | 0.1512 | ||||
| 37 | 0.402 | 0.385 | 0.1207 | ||||
| 44 | 0.390 | 0.369 | 0.0763 | ||||
| IgA (OD) | 23 | 0.405 | 0.401 | 0.0771 | 0.475 | 0.035 | 0.704 |
| 30 | 0.398 | 0.411 | 0.0651 | ||||
| 37 | 0.417 | 0.376 | 0.0751 | ||||
| 44 | 0.467 | 0.451 | 0.0580 | ||||
| IgM (OD) | 23 | 0.458 | 0.509 | 0.1048 | 0.453 | 0.175 | 0.821 |
| 30 | 0.479 | 0.479 | 0.1265 | ||||
| 37 | 0.489 | 0.535 | 0.1243 | ||||
| 44 | 0.566 | 0.555 | 0.1169 | ||||
| EPx (ng/mL) | 23 | 22.7 | 18.2 | 13.70 | |||
| 30 | 28.6 | 25.3 | 15.76 | ||||
| 37 | 44.3 | 37.0 | 20.29 | 0.509 | 0.001 | 0.814 | |
| 44 | 47.6 | 51.5 | 16.02 | ||||
| Day | SFP | Control | p | |
|---|---|---|---|---|
| IgA (OD) | 44 | 0.401 ± 0.0928 | 0.441 ± 0.0916 | 0.280 |
| Day | SFPs | Control | SD | p | |||
|---|---|---|---|---|---|---|---|
| Treatment (T) | Time | T × Time | |||||
| TAC (mmol/L) | 0 | 0.578 | 0.527 | 0.062 | 0.919 | 0.004 | 0.004 |
| 23 | 0.536 | 0.547 | 0.060 | ||||
| 30 | 0.522 | 0.552 | 0.110 | ||||
| 37 | 0.471 | 0.518 | 0.054 | ||||
| 44 | 0.509 | 0.483 | 0.065 | ||||
| GPx (U/g Hb) | 0 | 486.4 | 506.7 | 115.3 | 0.022 | 0.001 | 0.857 |
| 23 | 518.2 | 530.7 | 117.8 | ||||
| 30 | 380.4 | 434.0 | 97.16 | ||||
| 37 | 254.5 | 316.9 | 75.62 | ||||
| 44 | 175.3 | 224.5 | 72.15 | ||||
| MDA (µmol/L) | 0 | 0.265 | 0.269 | 0.037 | 0.060 | 0.001 | 0.599 |
| 23 | 0.235 | 0.247 | 0.033 | ||||
| 30 | 0.242 | 0.269 | 0.028 | ||||
| 37 | 0.241 | 0.248 | 0.041 | ||||
| 44 | 0.273 | 0.297 | 0.035 | ||||
| Tannins | Saponins | Alkaloids | Terpenoids | Flavonoids | Steroids | |
|---|---|---|---|---|---|---|
| SFPs | + | − | − | + | + | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komáromyová, M.; Petrič, D.; Kucková, K.; Batťányi, D.; Babják, M.; Dolinská, M.U.; Königová, A.; Barčák, D.; Dvorožňáková, E.; Čobanová, K.; et al. Impact of Sainfoin (Onobrychis viciifolia) Pellets on Parasitological Status, Antibody Responses, and Antioxidant Parameters in Lambs Infected with Haemonchus contortus. Pathogens 2022, 11, 301. https://doi.org/10.3390/pathogens11030301
Komáromyová M, Petrič D, Kucková K, Batťányi D, Babják M, Dolinská MU, Königová A, Barčák D, Dvorožňáková E, Čobanová K, et al. Impact of Sainfoin (Onobrychis viciifolia) Pellets on Parasitological Status, Antibody Responses, and Antioxidant Parameters in Lambs Infected with Haemonchus contortus. Pathogens. 2022; 11(3):301. https://doi.org/10.3390/pathogens11030301
Chicago/Turabian StyleKomáromyová, Michaela, Daniel Petrič, Katarína Kucková, Dominika Batťányi, Michal Babják, Michaela Urda Dolinská, Alžbeta Königová, Daniel Barčák, Emília Dvorožňáková, Klaudia Čobanová, and et al. 2022. "Impact of Sainfoin (Onobrychis viciifolia) Pellets on Parasitological Status, Antibody Responses, and Antioxidant Parameters in Lambs Infected with Haemonchus contortus" Pathogens 11, no. 3: 301. https://doi.org/10.3390/pathogens11030301