
The Use of Pythium oligandrum in the Biological Control of Roundworm Infection in Dogs and Cats


 ,
,
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Egg Collection
4.2. Sampling, Analysis and Preparation of Soil Samples
- n = 50 (Petri dishes with Toxocara canis eggs and soil): n = 25—samples exposed to Ecosin® (n = 5—samples for each soil type) and n = 25—control samples;
- n = 50 (Petri dishes with Toxocara cati eggs and soil): n = 25—samples exposed to Ecosin® (n = 5—samples for each soil type) and n = 25—control samples.
4.3. Preparation of Sandstone Media
- n = 10 (Toxocara canis eggs): n = 5—samples exposed to Ecosin® and n = 5—control samples;
- n = 10 (Toxocara cati eggs): n = 5—samples exposed to Ecosin® and n = 5—control samples.
4.4. Preparation of Pythium oligandrum Solution
4.5. Egg Exposure
4.6. Statistical Interpretation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Crompton, D.W. Ascaris and ascariasis. Adv. Parasitol. 2001, 48, 285–375. [Google Scholar] [CrossRef]
- Etewa, S.E.; Abdel-Rahman, S.A.; Abd El-Aal, N.F.; Fathy, G.M.; El-Shafey, M.A.; Ewis, A.M.G. Geohelminths distribution as affected by soil properties, physicochemical factors and climate in Sharkyia governorate Egypt. J. Parasit. Dis. 2016, 40, 496–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umar, A.A.; Bassey, S.E. Incidence of Strongyloides stercoralis infection in Ungogo, Nassarawa, Dala and Fagge local government areas of Kano state, Nigeria. Bayero J. Pure Appl. Sci. 2010, 3, 76–80. [Google Scholar] [CrossRef]
- Ciarmela, M.; Arambarri, A.; Basualdo, J.; Minvielle, M. Effect of saprotrophic soil fungi on Toxocara canis eggs. Malays. J. Microbiol. 2010, 6, 75–80. [Google Scholar] [CrossRef]
- De Souza Maia Filho, F.; de Oliveira da Silva Fonseca, A.; de Souza Silveira Valente, J.; Telles Baptista, C.; da Silva Moreira, A.; de Avila Botton, S.; Pötter, L.; Brayer Pereira, D.I. Exposure of Toxocara canis eggs to Purpureocillium lilacinum as a biocontrol strategy: An experimental model evaluation. Rev. Bras. Parasitol. Vet. 2019, 28, 91–96. [Google Scholar] [CrossRef]
- Hiura, E.; Del Carmen Garcia Lopes, A.; da Paz, J.S.; Gava, M.G.; Flecher, M.C.; Colares, M.; de Freitas Soares, F.E.; da Fonseca, L.A.; Lacerda, T.; de Araújo, J.V.; et al. Fungi predatory activity on embryonated Toxocara canis eggs inoculated in domestic chickens (Gallus gallus domesticus) and destruction of second stage larvae. Parasitol. Res. 2015, 114, 3301–3308. [Google Scholar] [CrossRef] [PubMed]
- Sudhakar, N.R.; Samanta, S.; Sahu, S.; Raina, O.K.; Gupta, S.C.; Madhu, D.N.; Kumar, A. Prevalence of Toxocara species eggs in soil samples of public health importance in and around Bareilly, Uttar Pradesh, India. Vet. World 2013, 6, 87–90. [Google Scholar] [CrossRef]
- Xavier, R.I.G.; Ramos, B.C.; Santarem, V.A. Recovery threshold of Toxocara canis eggs from soil. Vet. Parasitol. 2010, 167, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Cazapal-Monteiro, C.F.; Hernández, J.A.; Arroyo, F.L.; Miguélez, S.; Romasanta, Á.; Paz-Silva, A.; Sánchez-Andrade, R.; Arias, M.S. Analysis of the effect of soil saprophytic fungi on the eggs of Baylisascaris procyonis. Parasitol. Res. 2015, 114, 2443–2450. [Google Scholar] [CrossRef]
- Hernández, J.A.; Cazapal-Monteiro, C.F.; Arroyo, F.L.; Silva, M.I.; Palomero, A.M.; Paz-Silva, A.; Sánchez-Andrade, R.; Arias, M.S. Biological control of soil transmitted helminths (STHs) in a zoological park by using saprophytic fungi. Biol. Control 2018, 122, 24–30. [Google Scholar] [CrossRef]
- Bělonožníková, K.; Hýsková, V.; Chmelík, J.; Kavan, D.; Čeřovská, N.; Ryšlavá, H. Pythium oligandrum in plant protection and growth promotion: Secretion of hydrolytic enzymes, elicitors and tryptamine as auxin precursor. Microbiol. Res. 2022, 258, 126976. [Google Scholar] [CrossRef] [PubMed]
- Araújo, J.V.; Braga, F.R.; Mendoza-de-Gives, P.; Paz-Silva, A.; Vilela, V.L.R. Recent Advances in the Control of Helminths of Domestic Animals by Helminthophagous Fungi. Parasitologia 2021, 1, 168–176. [Google Scholar] [CrossRef]
- Ghimire, B.K.; Yu, C.Y.; Kim, S.H.; Chung, I.M. Diversity in Accessions of Panicum miliaceum L. Based on Agro-Morphological, Antioxidative, and Genetic Traits. Molecules 2019, 24, 1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, M.Y.; Seo, D.W.; Lee, J.Y.; Sung, M.K.; Lee, Y.M.; Jang, H.H.; Choi, H.Y.; Kim, J.H.; Park, D.S. Effects of Panicum miliaceum L. extract on adipogenic transcription factors and fatty acid accumulation in 3T3-L1 adipocytes. Nutr. Res. Pract. 2011, 5, 192–197. [Google Scholar] [CrossRef] [Green Version]
- Kolodziejczyk, L.; Mazurkiewicz-Zapa£owicz, K.; Twaruek, M.; Grajewski, J.; Lopusiewicz, L.; Rybiñska, A.; Dzika, E.; Pilarczyk, B. The Ovistatic Effect of Saprotrophic Soil Fungi on Ascaris suum Eggs. Folia Biol. 2019, 67, 1734–9168. [Google Scholar] [CrossRef]
- Thapa, S.; Meyling, N.V.; Katakam, K.K.; Thamsborg, S.M.; Mejer, H. A method to evaluate relative ovicidal effects of soil microfungi on thick-shelled eggs of animal-parasitic nematodes. Biocontrol Sci. Technol. 2015, 25, 756–767. [Google Scholar] [CrossRef]
- Hernández, J.A.; Vázquez-Ruiz, R.A.; Cazapal-Monteiro, C.F.; Valderrábano, E.; Arroyo, F.L.; Francisco, I.; Miguélez, S.; Sánchez-Andrade, R.; Paz-Silva, A.; Arias, M.S. Isolation of Ovicidal Fungi from Fecal Samples of Captive Animals Maintained in a Zoological Park. J. Fungi 2017, 3, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bojanich, M.V.; Basualdo, J.A.; Giusiano, G. In vitro effect of Chrysosporium indicum and Chrysosporium keratinophylum on Toxocara canis eggs. Rev. Argent. Microbiol. 2018, 50, 249–254. [Google Scholar] [CrossRef]
- Abou-El-Naga, I.F. Developmental stages and viability of Toxocara canis eggs outside the host. Biomedica 2018, 38, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Viña, C.; Silva, M.I.; Palomero, A.M.; Voinot, M.; Vilá, M.; Hernández, J.Á.; Paz-Silva, A.; Sánchez-Andrade, R.; Cazapal-Monteiro, C.F.; Arias, M.S. The Control of Zoonotic Soil-Transmitted Helminthoses Using Saprophytic Fungi. Pathogens 2020, 9, 1071. [Google Scholar] [CrossRef] [PubMed]
- Lýsek, H.; Ondrus, J. Morphology of the uterus of Ascaris lumbricoides in the region where fertilization and formation of eggshell occur. Folia Parasitol. 1992, 39, 41–50. [Google Scholar]
- Gee, G.W.; Bauder, J.W. Physical and Mineralogical Methods, 2nd ed.; Agronomy Monograph No. 9; American Society of Agronomy/Soil Science Society of America: Madison, WI, USA, 1986; pp. 383–411. [Google Scholar]
- Mabaso, M.L.; Appleton, C.C.; Hughes, J.C.; Gouws, E. The effect of soil type and climate on hookworm (Necator americanus) distribution in KwaZulu-Natal, South Africa. Trop. Med. Int. Health 2003, 8, 722–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, H.H. A Simple Levitation Method for the Detection of Hookworm Ova. Med. J. Aust. 1921, 2, 375–376. [Google Scholar] [CrossRef]
- Gautam, S.; Petkeviciute, E.; Storm, N.T.; Thapa, S.; Mejer, H. Effect of disinfectants on viability of Ascaris suum and Ascaridia galli eggs. In Spring Symposium of the Danish Society for Parasitlogy; Organic eprints: Copenhagen, Denmark, 2014. [Google Scholar]



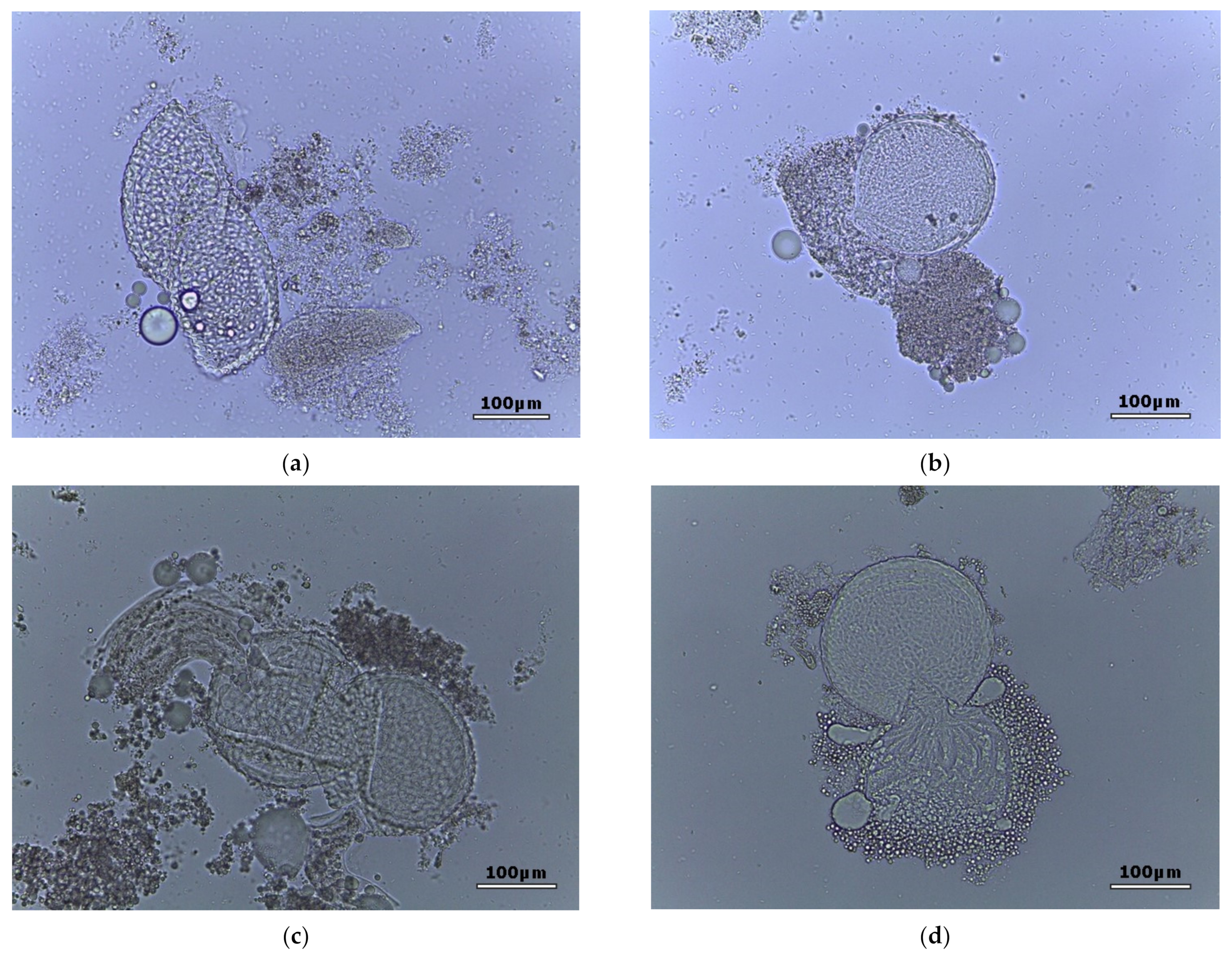{kind=link}
{kind=link}
| Sample | Media | Viable Eggs | Non-Viable Eggs | |||
|---|---|---|---|---|---|---|
| % Larvated (am) | % Developed (Prelarval Stages) (am) | % Undeveloped (am) | % Degenerate (am) | Mann-Whitney Test (p Value) | ||
| treated (n = 5) | sandy-clayey clay | 0 | 43.8 | 21.2 | 35 | 0.0079 (**) |
| control (n = 5) | sandy-clayey clay | 0 | 67.4 | 29.2 | 3.4 | |
| treated (n = 5) | medium clay | 0 | 32.8 | 44.6 | 22.6 | 0.0159 (*) |
| control (n = 5) | medium clay | 0 | 63.4 | 26.2 | 10.4 | |
| treated (n = 5) | medium sandy clay | 0 | 22.8 | 38.2 | 41 | 0.0079 (**) |
| control (n = 5) | medium sandy clay | 0 | 65.2 | 23.8 | 11 | |
| treated (n = 5) | loamy fine sand | 0 | 48.6 | 31.6 | 21.8 | 0.1032 (ns) |
| control (n = 5) | loamy fine sand | 0 | 69 | 20.6 | 10.4 | |
| treated (n = 5) | coarse sandy clay | 0 | 49.8 | 32.4 | 17.8 | 0.0952 (ns) |
| control (n = 5) | coarse sandy clay | 0 | 69.2 | 25.2 | 5.6 | |
| treated (n = 5) | sandstone media | 0 | 45.2 | 33.8 | 21 | 0.0079 (**) |
| control (n = 5) | sandstone media | 0 | 71 | 19.6 | 9.4 | |
| Sample | Media | Viable Eggs | Non-Viable Eggs | |||
|---|---|---|---|---|---|---|
| % Larvated (am) | % Developed (Prelarval Stages) (am) | % Undeveloped (am) | % Degenerate (am) | Mann-Whitney Test (p Value) | ||
| treated (n = 5) | sandy-clayey clay | 0 | 37.6 | 27.4 | 35 | 0.0079 (**) |
| control (n = 5) | sandy-clayey clay | 0 | 67.6 | 20 | 12.4 | |
| treated (n = 5) | medium clay | 0 | 30.2 | 48.6 | 21.2 | |
| control (n = 5) | medium clay | 0 | 69.6 | 20.2 | 10.2 | |
| treated (n = 5) | medium sandy clay | 0 | 29 | 24.6 | 46.4 | |
| control (n = 5) | medium sandy clay | 0 | 66.8 | 21.2 | 12 | |
| treated (n = 5) | loamy fine sand | 0 | 39.2 | 30.8 | 30 | |
| control (n = 5) | loamy fine sand | 0 | 69.2 | 21.2 | 9.6 | |
| treated (n = 5) | coarse sandy clay | 0 | 39 | 30.8 | 30.2 | |
| control (n = 5) | coarse sandy clay | 0 | 67.6 | 24.4 | 10 | |
| treated (n = 5) | sandstone media | 0 | 37.6 | 31.4 | 31 | |
| control (n = 5) | sandstone media | 0 | 68.6 | 22.8 | 8.6 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luca, I.; Ilie, M.S.; Florea, T.; Olariu-Jurca, A.; Stancu, A.; Dărăbuş, G. The Use of Pythium oligandrum in the Biological Control of Roundworm Infection in Dogs and Cats. Pathogens 2022, 11, 367. https://doi.org/10.3390/pathogens11030367
Luca I, Ilie MS, Florea T, Olariu-Jurca A, Stancu A, Dărăbuş G. The Use of Pythium oligandrum in the Biological Control of Roundworm Infection in Dogs and Cats. Pathogens. 2022; 11(3):367. https://doi.org/10.3390/pathogens11030367
Chicago/Turabian StyleLuca, Iasmina, Marius Stelian Ilie, Tiana Florea, Adrian Olariu-Jurca, Adrian Stancu, and Gheorghe Dărăbuş. 2022. "The Use of Pythium oligandrum in the Biological Control of Roundworm Infection in Dogs and Cats" Pathogens 11, no. 3: 367. https://doi.org/10.3390/pathogens11030367