
Serotype Distribution, Virulence Determinants and Antimicrobial Susceptibility of Streptococcus agalactiae Isolated from Young Infants
 , , and
, , and
Abstract
:1. Introduction
2. Methods and Materials
2.1. Clinical Isolates
2.2. Ethical Approvals
2.3. Laboratory Methods
2.4. Whole Genome Sequencing
2.5. Molecular Subtyping
2.6. Multilocus Sequence Type
3. Results
3.1. General Characteristics
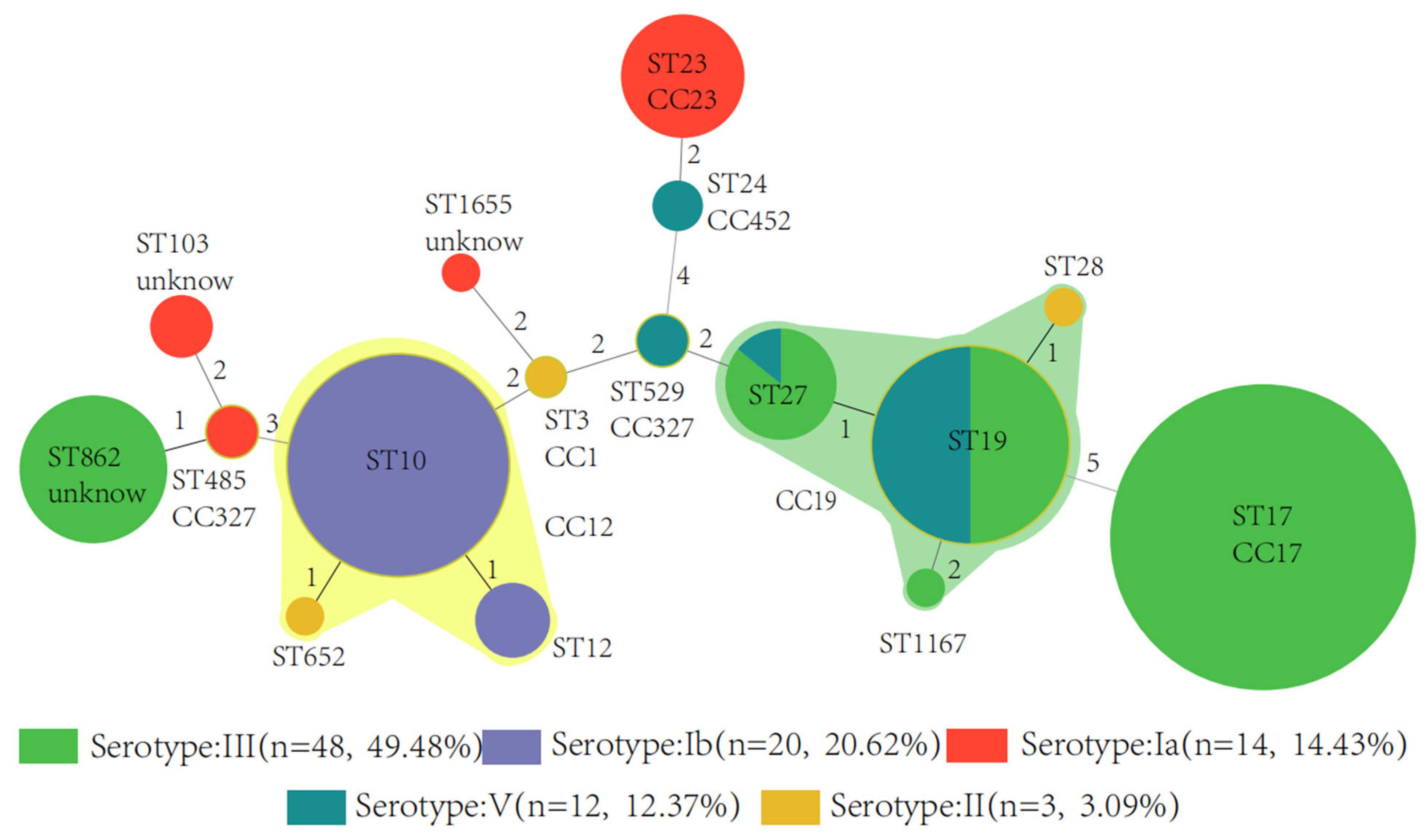
3.2. Serotype, Sequence Type, and Clonal Complex


3.3. Surface Protein Genes
3.4. Antimicrobial Resistance Genes
3.5. Virulence Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Seale, A.C.; Bianchi-Jassir, F.; Russell, N.J.; Kohli-Lynch, M.; Tann, C.J.; Hall, J.; Madrid, L.; Blencowe, H.; Cousens, S.; Baker, C.J.; et al. Estimates of the Burden of Group B Streptococcal Disease Worldwide for Pregnant Women, Stillbirths, and Children. Clin. Infect. Dis. 2017, 65, S200–S219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Goffau, M.C.; Lager, S.; Sovio, U.; Gaccioli, F.; Cook, E.; Peacock, S.J.; Parkhill, J.; Charnock-Jones, D.S.; Smith, G.C.S. Human placenta has no microbiome but can contain potential pathogens. Nature 2019, 574, E15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncalves, B.P.; Procter, S.R.; Paul, P.; Chandna, J.; Lewin, A.; Seedat, F.; Koukounari, A.; Dangor, Z.; Leahy, S.; Santhanam, S.; et al. Group B Streptococcus infection during pregnancy and infancy: Estimates of regional and global burden. Lancet. Global Health 2022, 10, e807–e819. [Google Scholar] [CrossRef]
- Ji, W.J.; Liu, H.Y.; Madhi, S.A.; Cunnington, M.; Zhang, Z.L.; Dangor, Z.; Zhou, H.J.; Mu, X.P.; Jin, Z.J.; Wang, A.M.; et al. Clinical and Molecular Epidemiology of Invasive Group B Streptococcus Disease among Infants, China. Emerg. Infect. Dis. 2019, 25, 2021–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madrid, L.; Seale, A.C.; Kohli-Lynch, M.; Edmond, K.M.; Lawn, J.E.; Heath, P.T.; Madhi, S.A.; Baker, C.J.; Bartlett, L.; Cutland, C.; et al. Infant Group B Streptococcal Disease Incidence and Serotypes Worldwide: Systematic Review and Meta-analyses. Clin. Infect. Dis. 2017, 65, S160–S172. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.J.; Wang, Y.J.; Hsia, Y.F.; Russell, N.; Heath, P.T. Systematic Review and Meta-Analyses of Incidence for Group B Streptococcus Disease in Infants and Antimicrobial Resistance, China. Emerg. Infect. Dis. 2020, 26, 2651–2659. [Google Scholar] [CrossRef]
- Schrag, S.J.; Zywicki, S.; Farley, M.M.; Reingold, A.L.; Harrison, L.H.; Lefkowitz, L.B.; Hadler, J.L.; Danila, R.; Cieslak, P.R.; Schuchat, A. Group B streptococcal disease in the era of intrapartum antibiotic prophylaxis. N. Engl. J. Med. 2000, 342, 15–20. [Google Scholar] [CrossRef]
- Hayes, K.; O’Halloran, F.; Cotter, L. A review of antibiotic resistance in Group B Streptococcus: The story so far. Crit. Rev. Microbiol. 2020, 46, 253–269. [Google Scholar] [CrossRef]
- Plainvert, C.; Hays, C.; Touak, G.; Joubrel-Guyot, C.; Dmytruk, N.; Frigo, A.; Poyart, C.; Tazi, A. Multidrug-Resistant Hypervirulent Group B Streptococcus in Neonatal Invasive Infections, France, 2007–2019. Emerg. Infect. Dis. 2020, 26, 2721–2724. [Google Scholar] [CrossRef]
- Gizachew, M.; Tiruneh, M.; Moges, F.; Tessema, B. Streptococcus agalactiae maternal colonization, antibiotic resistance and serotype profiles in Africa: A meta-analysis. Ann. Clin. Microb. Anti. 2019, 18, 14. [Google Scholar] [CrossRef]
- Vornhagen, J.; Waldorf, K.M.A.; Rajagopal, L. Perinatal Group B Streptococcal Infections: Virulence Factors, Immunity, and Prevention Strategies. Trends Microbiol. 2017, 25, 919–931. [Google Scholar] [CrossRef]
- Lo, C.W.; Liu, H.C.; Lee, C.C.; Lin, C.L.; Chen, C.L.; Jeng, M.J.; Chiu, C.H. Serotype distribution and clinical correlation of Streptococcus agalactiae causing invasive disease in infants and children in Taiwan. J. Microbiol. Immunol. Infect. 2019, 52, 578–584. [Google Scholar] [CrossRef]
- Jiang, S.M.; Park, S.E.; Yadav, P.; Paoletti, L.C.; Wessels, M.R. Regulation and Function of Pilus Island 1 in Group B Streptococcus. J. Bacteriol. 2012, 194, 2479–2490. [Google Scholar] [CrossRef]
- Lazzarin, M.; Mu, R.; Fabbrini, M.; Ghezzo, C.; Rinaudo, C.D.; Doran, K.S.; Margarit, I. Contribution of pilus type 2b to invasive disease caused by a Streptococcus agalactiae ST-17 strain. BMC Microbiol. 2017, 17, 8. [Google Scholar] [CrossRef] [Green Version]
- Shabayek, S.; Spellerberg, B. Group B Streptococcal Colonization, Molecular Characteristics, and Epidemiology. Front. Microbiol. 2018, 9, 14. [Google Scholar] [CrossRef]
- Bianchi-Jassir, F.; Paul, P.; To, K.N.; Carreras-Abad, C.; Seale, A.C.; Jauneikaite, E.; Madhi, S.A.; Russell, N.J.; Hall, J.; Madrid, L.; et al. Systematic review of Group B Streptococcal capsular types, sequence types and surface proteins as potential vaccine candidates. Vaccine 2020, 38, 6682–6694. [Google Scholar] [CrossRef]
- Song, J.Y.; Lim, J.H.; Lim, S.; Yong, Z.; Seo, H.S. Progress toward a group B streptococcal vaccine. Hum. Vaccines Immunother. 2018, 14, 2669–2681. [Google Scholar] [CrossRef]
- Absalon, J.; Segall, N.; Block, S.L.; Center, K.J.; Scully, I.L.; Giardina, P.C.; Peterson, J.; Watson, W.J.; Gruber, W.C.; Jansen, K.U.; et al. Safety and immunogenicity of a novel hexavalent Group B Streptococcus conjugate vaccine in healthy, non-pregnant adults: A phase 1/2, randomised, placebo-controlled, observer-blinded, dose-escalation trial. Lancet Infect. Dis. 2021, 21, 263–274. [Google Scholar] [CrossRef]
- Beran, J.; Leroux-Roels, G.; Van Damme, P.; de Hoon, J.; Vandermeulen, C.; Al-Ibrahim, M.; Johnson, C.; Peterson, J.; Baker, S.; Seidl, C.; et al. Safety and immunogenicity of fully liquid and lyophilized formulations of an investigational trivalent Group B Streptococcus vaccine in healthy non-pregnant women: Results from a randomized comparative phase II trial. Vaccine 2020, 38, 3227–3234. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Le Doare, K.; O’Driscoll, M.; Turner, K.; Seedat, F.; Russell, N.J.; Seale, A.C.; Heath, P.T.; Lawn, J.E.; Baker, C.J.; Bartlett, L.; et al. Intrapartum Antibiotic Chemoprophylaxis Policies for the Prevention of Group B Streptococcal Disease Worldwide: Systematic Review. Clin. Infect. Dis. 2017, 65, S143–S151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.M. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Jones, N.; Bohnsack, J.F.; Takahashi, S.; Oliver, K.A.; Chan, M.S.; Kunst, F.; Glaser, P.; Rusniok, C.; Crook, D.W.; Harding, R.M.; et al. Multilocus sequence typing system for Group B streptococcus. J. Clin. Microbiol. 2003, 41, 2530–2536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, S.P.; Lu, X.D.; Jin, Z.J.; Gao, J.Y.; Ma, D.L.; Deng, J.K.; Wu, X.L.; Hu, Y.W.; Zheng, L.; Wang, Q. Characterization of Group B Streptococcus isolated from sterile and non-sterile specimens in China. Diagn. Microbiol. Infect. Dis. 2018, 92, 56–61. [Google Scholar] [CrossRef]
- Guo, H.; Fu, M.; Peng, Q.; Chen, Z.; Liu, J.; Qiu, Y.; Huang, Y. Antimicrobial resistance and molecular characterization of Streptococcus agalactiae from pregnant women in southern China. J. Infect. Dev. Ctries. 2019, 13, 802–809. [Google Scholar] [CrossRef] [Green Version]
- Musilova, I.; Pliskova, L.; Kutova, R.; Jacobsson, B.; Paterova, P.; Kacerovsky, M. Streptococcus agalactiae in pregnancies complicated by preterm prelabor rupture of membranes. J. Matern.-Fetal. Neonatal. Med. 2016, 29, 1036–1040. [Google Scholar] [CrossRef]
- Yaseen, S.; Asghar, S.; Shahzadi, I.; Qayyum, A. Ascertaining the Prevalence of Group B Streptococcal Infection in Patients with Preterm Premature Rupture of Membranes: A Cross-Sectional Analysis from Pakistan. Cureus 2021, 13, 6. [Google Scholar] [CrossRef]
- Dzanibe, S.; Madhi, S.A. Systematic review of the clinical development of Group B Streptococcus serotype-specific capsular polysaccharide-based vaccines. Expert. Rev. Vaccines 2018, 17, 635–651. [Google Scholar] [CrossRef]
- Paoletti, L.C.; Madoff, L.C. Vaccines to prevent neonatal GBS infection. Semin. Neonatol. SN 2002, 7, 315–323. [Google Scholar] [CrossRef]
- Nuccitelli, A.; Rinaudo, C.D.; Maione, D. Group B Streptococcus vaccine: State of the art. Ther. Adv. Vaccines 2015, 3, 76–90. [Google Scholar] [CrossRef] [Green Version]
- Le Doare, K.; Heath, P.T. An overview of global GBS epidemiology. Vaccine 2013, 31, D7–D12. [Google Scholar] [CrossRef]
- Tiruvayipati, S.; Tang, W.Y.; Barkham, T.M.S.; Chen, S.L. GBS-SBG-GBS Serotyping by Genome Sequencing. Microb. Genom. 2021, 7, 000688. [Google Scholar] [CrossRef]
- Chinthamani, R.; Pathirage, S.; Chandrasiri, S. Serotyping of invasive and colonizing Group B Streptococcus (GBS) isolates at selected hospitals in Sri Lanka: A multicenter study. Ceylon Med. J. 2021, 66, 144–150. [Google Scholar] [CrossRef]
- Liu, J.Y.; Xu, R.R.; Zhong, H.M.; Zhong, Y.K.; Xie, Y.Q.; Li, L.; Li, B.; Chen, D.Q.; Xu, Z.B. Prevalence of GBS serotype III and identification of a ST 17-like genotype from neonates with invasive diseases in Guangzhou, China. Microb. Pathog. 2018, 120, 213–218. [Google Scholar] [CrossRef]
- Vekemans, J.; Moorthy, V.; Friede, M.; Alderson, M.R.; Sobanjo-Ter Meulen, A.; Baker, C.J.; Heath, P.T.; Madhi, S.A.; Mehring-Le Doare, K.; Saha, S.K.; et al. Maternal immunization against Group B Streptococcus: World Health Organization research and development technological roadmap and preferred product characteristics. Vaccine 2019, 37, 7391–7393. [Google Scholar] [CrossRef]
- Heath, P.T. Status of vaccine research and development of vaccines for GBS. Vaccine 2016, 34, 2876–2879. [Google Scholar] [CrossRef] [Green Version]
- Papasergi, S.; Brega, S.; Mistou, M.Y.; Firon, A.; Oxaran, V.; Dover, R.; Teti, G.; Shai, Y.; Trieu-Cuot, P.; Dramsi, S. The GBS PI-2a Pilus Is Required for Virulence in Mice Neonates. PLoS ONE 2011, 6, 8. [Google Scholar] [CrossRef]
- Koide, S.; Nagano, Y.; Takizawa, S.; Sakaguchi, K.; Soga, E.; Hayashi, W.; Tanabe, M.; Denda, T.; Kimura, K.; Arakawa, Y.; et al. Genomic Traits Associated with Virulence and Antimicrobial Resistance of Invasive Group B Streptococcus Isolates with Reduced Penicillin Susceptibility from Elderly Adults. Microbiol. Spectr. 2022, 10, e0056822. [Google Scholar] [CrossRef]
- Forquin, M.P.; Tazi, A.; Rosa-Fraile, M.; Poyart, C.; Trieu-Cuot, P.; Dramsi, S. The putative glycosyltransferase-encoding gene cylJ and the Group B Streptococcus (GBS)-specific gene cylK modulate hemolysin production and virulence of GBS. Infect. Immun. 2007, 75, 2063–2066. [Google Scholar] [CrossRef]
- Vornhagen, J.; Quach, P.; Boldenow, E.; Merillat, S.; Whidbey, C.; Ngo, L.Y.; Waldorf, K.M.A.; Rajagopal, L. Bacterial Hyaluronidase Promotes Ascending GBS Infection and Preterm Birth. mBio 2016, 10, e00781-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, B.; Wang, D.; Zhou, H.; Zhu, F.; Li, D.; Zhang, S.; Shi, Y.; Cui, Y.; Huang, L.; Wu, H. Distribution of pilus islands and alpha-like protein genes of Group B Streptococcus colonized in pregnant women in Beijing, China. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1173–1179. [Google Scholar] [CrossRef] [PubMed]
- Bob-Manuel, M.; McGee, L.; Igunma, J.A.; Alex-Wele, M.A.; Obunge, O.K.; Wariso, K.T. Whole genome sequence based capsular typing and antimicrobial resistance prediction of Group B streptococcal isolates from colonized pregnant women in Nigeria. BMC Genom. 2021, 22, 6. [Google Scholar] [CrossRef] [PubMed]
- Shabayek, S.; Abdalla, S.; Abouzeid, A.M. Serotype and surface protein gene distribution of colonizing Group B Streptococcus in women in Egypt. Epidemiol. Infect. 2014, 142, 208–210. [Google Scholar] [CrossRef] [PubMed]
- Dangor, Z.; Kwatra, G.; Izu, A.; Khan, M.; Lala, S.G.; Madhi, S.A.; Streptococcus, G.B. Infant serotype specific anti-capsular immunoglobulin G antibody and risk of invasive group B Streptococcal disease. Vaccine 2021, 39, 6813–6816. [Google Scholar] [CrossRef]
- Martins, E.R.; Andreu, A.; Melo-Cristino, J.; Ramirez, M. Distribution of Pilus Islands in Streptococcus agalactiae That Cause Human Infections: Insights into Evolution and Implication for Vaccine Development. Clin. Vaccine Immunol. 2013, 20, 313–316. [Google Scholar] [CrossRef] [Green Version]
- Lu, B.; Wu, J.; Chen, X.; Gao, C.; Yang, J.; Li, Y.; Wang, J.; Zeng, J.; Fang, Y.; Wang, D.; et al. Microbiological and clinical characteristics of Group B Streptococcus isolates causing materno-neonatal infections: High prevalence of CC17/PI-1 and PI-2b sublineage in neonatal infections. J. Med. Microbiol. 2018, 67, 1551–1559. [Google Scholar] [CrossRef]
- Suresh, S.; Tyrrell, G.; Alhhazmi, A.; Escoredo, S.; Hawkes, M. Recurrent sepsis and neuroinvasive disease in a neonate culture-positive for a Group B Streptococcus CPS III serotype, hvgA+ strain. JMM Case Rep. 2016, 3, e005034. [Google Scholar] [CrossRef] [Green Version]
- Tazi, A.; Bellais, S.; Tardieux, I.; Dramsi, S.; Trieu-Cuot, P.; Poyart, C. Group B Streptococcus surface proteins as major determinants for meningeal tropism. Curr. Opin. Microbiol. 2012, 15, 44–49. [Google Scholar] [CrossRef]
- de Cambronne, R.D.; Fouet, A.; Picart, A.; Bourrel, A.S.; Anjou, C.; Bouvier, G.; Candeias, C.; Bouaboud, A.; Costa, L.; Boulay, A.C.; et al. CC17 Group B Streptococcus exploits integrins for neonatal meningitis development. J. Clin. Investig. 2021, 131, 16. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, L.Q.; Yan, Q.; Lee, C.C.; Hsu, M.H.; Liao, W.T.; Zhang, L.; Chiu, C.H. Genomic Analysis of Group B Streptococcus from Neonatal Sepsis Reveals Clonal CC17 Expansion and Virulence- and Resistance-Associated Traits After Intrapartum Antibiotic Prophylaxis. Clin. Infect. Dis. 2022, 29, ciac331. [Google Scholar] [CrossRef]
- Yang, Y.C.; Liu, Y.L.; Ding, Y.L.; Yi, L.; Ma, Z.; Fan, H.J.; Lu, C.P. Molecular Characterization of Streptococcus agalactiae Isolated from Bovine Mastitis in Eastern China. PLoS ONE 2013, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Kaczorek, E.; Malaczewska, J.; Wojcik, R.; Siwicki, A.K. Biofilm production and other virulence factors in Streptococcus spp. isolated from clinical cases of bovine mastitis in Poland. BMC Vet. Res. 2017, 13, 7. [Google Scholar] [CrossRef] [Green Version]
- Emaneini, M.; Mirsalehian, A.; Beigvierdi, R.; Fooladi, A.A.I.; Asadi, F.; Jabalameli, F.; Taherikalani, M. High Incidence of Macrolide and Tetracycline Resistance among Streptococcus Agalactiae Strains Isolated from Clinical Samples in Tehran, Iran. Maedica 2014, 9, 157–161. [Google Scholar]
- Hraoui, M.; Boutiba-Ben Boubaker, I.; Rachdi, M.; Slim, A.; Ben Redjeb, S. Macrolide and tetracycline resistance in clinical strains of Streptococcus agalactiae isolated in Tunisia. J. Med. Microbiol. 2012, 61, 1109–1113. [Google Scholar] [CrossRef]
- Dinos, G.P. The macrolide antibiotic renaissance. Br. J. Pharmacol. 2017, 174, 2967–2983. [Google Scholar] [CrossRef] [Green Version]
- Morozumi, M.; Wajima, T.; Kuwata, Y.; Chiba, N.; Sunaoshi, K.; Sugita, K.; Sakata, H.; Iwata, S.; Ubukata, K. Associations between capsular serotype, multilocus sequence type, and macrolide resistance in Streptococcus agalactiae isolates from Japanese infants with invasive infections. Epidemiol. Infect. 2014, 142, 812–819. [Google Scholar] [CrossRef] [Green Version]
- Lopes, E.; Fernandes, T.; Machado, M.P.; Carrico, J.A.; Mello-Cristino, J.; Ramirez, M.; Martins, E.R.; Portuguese Grp Study, S. Increasing macrolide resistance among Streptococcus agalactiae causing invasive disease in non-pregnant adults was driven by a single capsular-transformed lineage, Portugal, 2009 to 2015. Eurosurveillance 2018, 23, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Lu, B.H.; Chen, X.C.; Wang, J.R.; Wang, D.C.; Zeng, J.; Li, Y.; Li, D.; Zhu, F.X.; Cui, Y.C.; Huang, L. Molecular characteristics and antimicrobial resistance in invasive and noninvasive Group B Streptococcus between 2008 and 2015 in China. Diagn. Microbiol. Infect. Dis. 2016, 86, 351–357. [Google Scholar] [CrossRef]
- Martel, A.; Baele, M.; Devriese, L.A.; Goossens, H.; Wisselink, H.J.; Decostere, A.; Haesebrouck, F. Prevalence and mechanism of resistance against macrolides and lincosamides in Streptococcus suis isolates. Vet. Microbiol. 2001, 83, 287–297. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EOD (n = 81) | LOD (n= 16) | Total (n = 97) | |
|---|---|---|---|
| Age (days) | |||
| Median (IQR) | 1 (1–1) | 20 (17–24) | 1 (1–1) |
| Gender (n, %) | |||
| Male | 47 (58.0) | 8 (50.0) | 55 (56.7) |
| Female | 34 (42.0) | 8 (50.0) | 42 (43.3) |
| Birth weight (grams) | |||
| All cases (median (IQR)) | 3150 (2600–3400) | 3200 (3000–3515) | 3150 (2757–3450) |
| Apgar-1min | |||
| Median (IQR) | 9 (9–10) | 10 (8–10) | 9 (9–10) |
| Apgar-5 min | |||
| Median (IQR) | 10 (10–10) | 10 (10–10) | 10 (10–10) |
| Single or twins (n, %) | |||
| Single | 79 (97.5) | 15 (93.8) | 94 (96.9) |
| Twins or multiple | 2 (2.5) | 1 (6.3) | 3 (3.1) |
| Gestational age (weeks) | |||
| Median (IQR) | 39 (36–40) | 39 (38–40) | 39 (37–40) |
| Delivery mode (n, %) | |||
| Vaginal | 53 (65.4) | 9 (56.3) | 62 (63.9) |
| C-section | 28 (34.6) | 7 (43.8) | 35 (36.1) |
| Specimen source (n, %) | |||
| Blood | 31 (38.3) | 12 (75.0) | 43 (44.3) |
| Gastricus | 30 (37.0) | 0 (0.0) | 30 (30.9) |
| Sputum | 17 (21.0) | 0 (0.0) | 17 (17.5) |
| Cerebrospinal fluid | 1 (1.2) | 4 (25.0) | 5 (5.2) |
| Ear canal secretion | 1 (1.2) | 0 (0.0) | 1 (1.0) |
| Umbilical secretion | 1 (1.2) | 0 (0.0) | 1 (1.0) |
| Premature rupture of membranes (n, %) | |||
| No | 65 (80.2) | 15 (93.8) | 80 (82.5) |
| Yes | 16 (19.8) | 1 (6.3) | 17 (17.5) |
| Pregnancy type (n, %) | |||
| First time | 52 (64.2) | 8 (50.0) | 60 (61.9) |
| Non-first | 29 (35.8) | 8 (50.0) | 37 (38.1) |
| Length of stay in ICU (days) | |||
| Median (IQR) | 10 (7–15) | 14 (8–30) | 11 (7–16) |
| Serotype | Srr | Pili | HvgA | Macrolide/ Clindamycin Genes | Tetracycline Resistance Genes | LsaE/LsaC | Chloramphenicol Genes |
|---|---|---|---|---|---|---|---|
| Ia | srr1 (11) | pi-1 (1); pi-2a (8); pi-2b (6); pi-1:pi-2b (1) | ermB (1); mef(4); lnuB (1) | tetM (3); tetO (1); tetL (1) | lsaC (3); lsaE (1) | ||
| Ib | srr1 (17) | pi-1 (18); pi-2a (20); pi-1:pi-2a (18) | ermB (20); lnuB (1) | tetO (3) | lsaE (1) | ||
| II | srr1 (3) | pi-1 (3); pi-2a (3); pi-1:pi-2a (3) | ermB (1); ermTR(4) | tetM (2); tetL (1) | cat (1) | ||
| III | srr1 (14); srr2 (24) | pi-1 (18); pi-2a (14); pi-2b (34); pi-1:pi-2a (13); pi-1:pi-2b (5) | HvgA (24) | ermB (31); ermT (1); mef(13); lnuB (8) | tetM (7); tetO (26); tetL (1); | lsaC (1); lsaE (8) | |
| V | pi-1 (9); pi-2a (12); pi-1:pi-2a (9) | ermB (2); ermTR(8); mef(6); lnuB (1) | tetM (7); tetO (3); tetL (1); | lsaE (1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, Z.; Li, J.; Zhou, H.; Wang, Z.; Yi, L.; Liu, N.; Du, J.; Chang, C.-Y.; Ji, W. Serotype Distribution, Virulence Determinants and Antimicrobial Susceptibility of Streptococcus agalactiae Isolated from Young Infants. Pathogens 2022, 11, 1355. https://doi.org/10.3390/pathogens11111355
Jin Z, Li J, Zhou H, Wang Z, Yi L, Liu N, Du J, Chang C-Y, Ji W. Serotype Distribution, Virulence Determinants and Antimicrobial Susceptibility of Streptococcus agalactiae Isolated from Young Infants. Pathogens. 2022; 11(11):1355. https://doi.org/10.3390/pathogens11111355
Chicago/Turabian StyleJin, Zhengjiang, Juan Li, Haijian Zhou, Zhenhui Wang, Lu Yi, Nian Liu, Jiaxi Du, Chien-Yi Chang, and Wenjing Ji. 2022. "Serotype Distribution, Virulence Determinants and Antimicrobial Susceptibility of Streptococcus agalactiae Isolated from Young Infants" Pathogens 11, no. 11: 1355. https://doi.org/10.3390/pathogens11111355