
Transmission Cycle of Tick-Borne Infections and Co-Infections, Animal Models and Diseases

Abstract
:1. Introduction
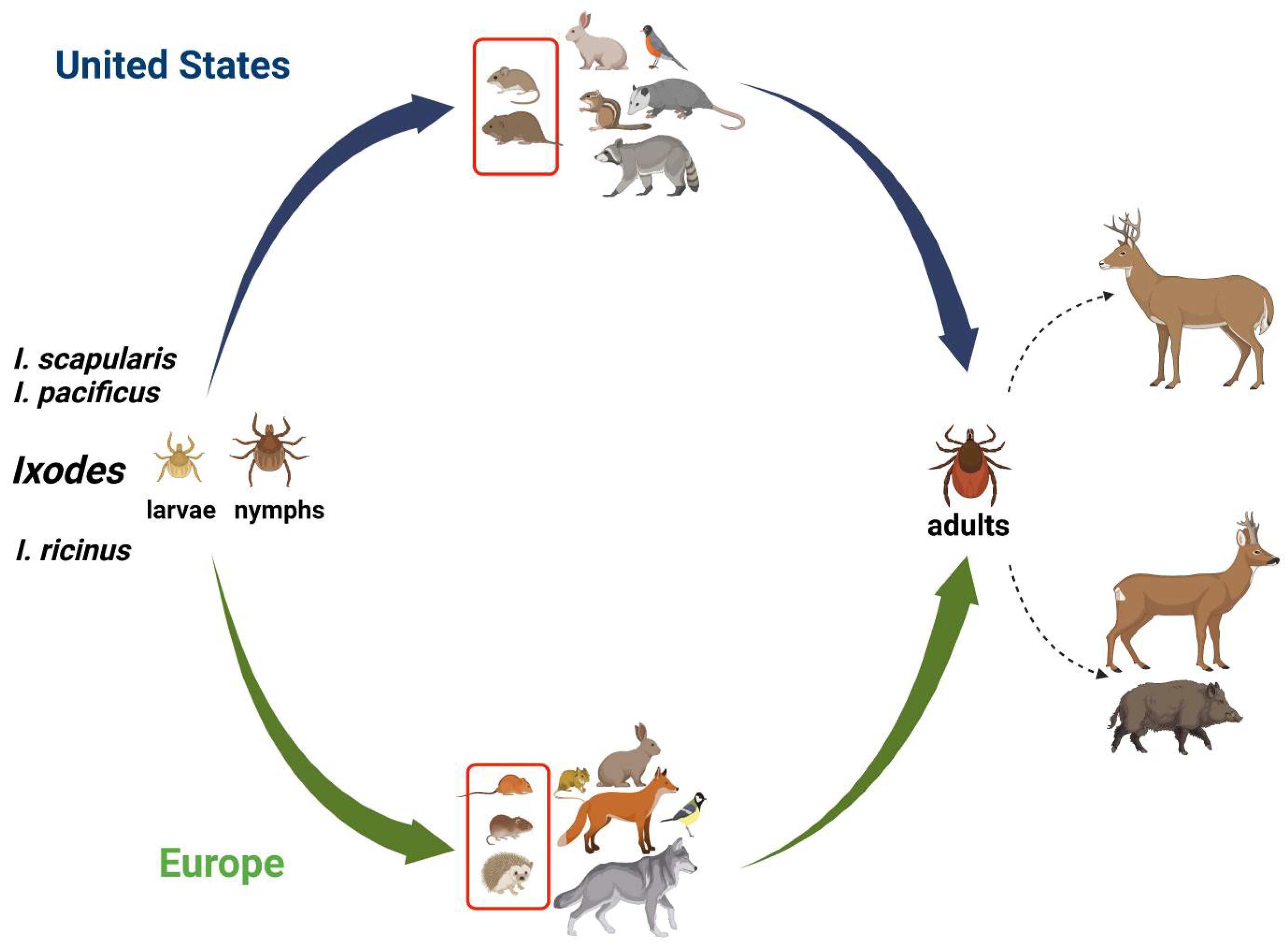
2. Ticks as Vectors of Transmission of Co-Infections
3. Wildlife as a Reservoir Host for TBCI
3.1. Small Mammalian Species
3.2. Meso-Mammal Species
3.3. Large Mammal Species
3.4. Birds
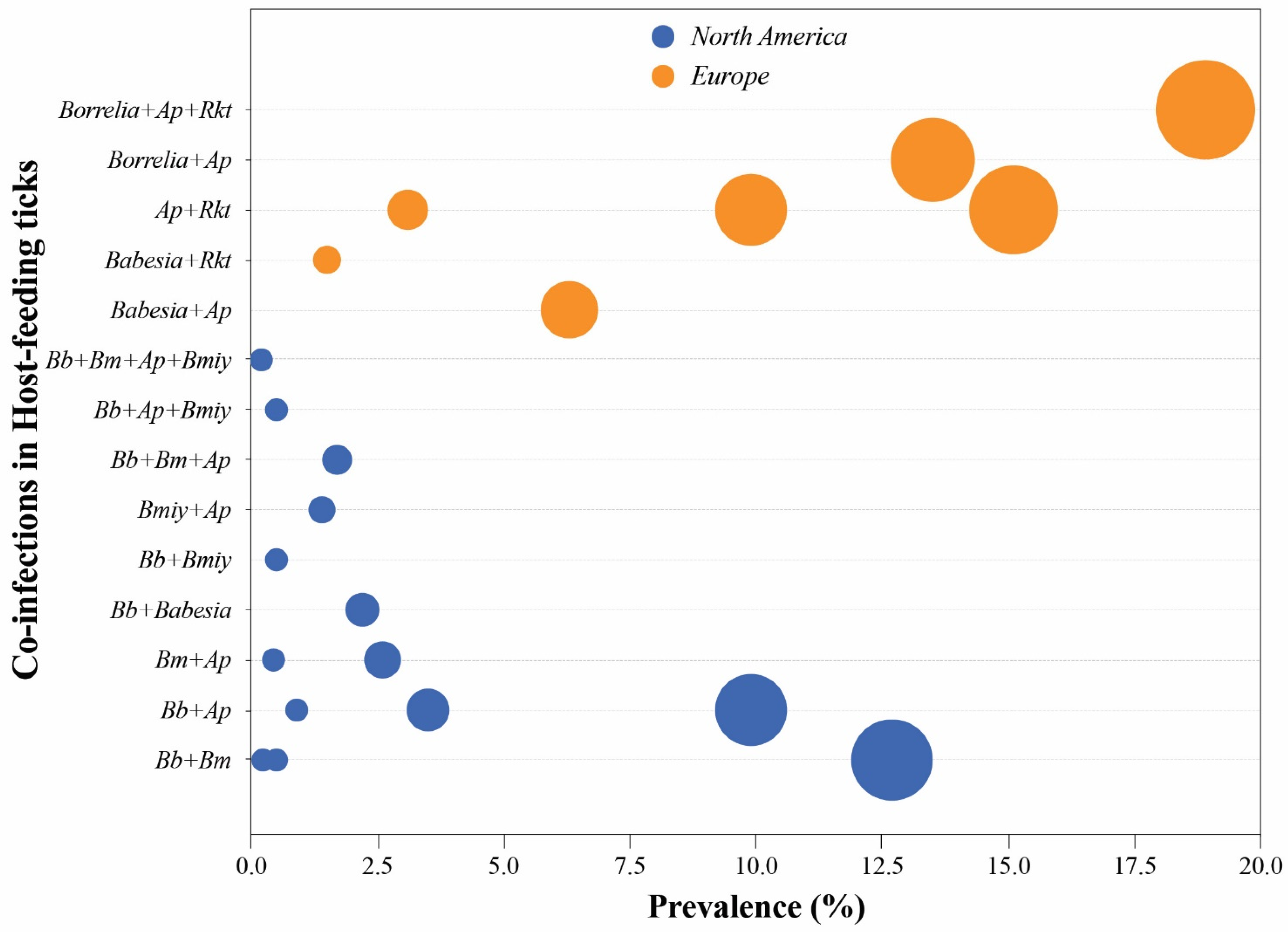
4. TBPs Prevalence in Reservoir Wildlife versus Surveillance of Ticks
5. Major Virulence Factors of Pathogens That Cause Co-Infection
6. Animal Models for Investigation of TBD and TBCI
7. Evaluation of Vaccine Candidates for Major Pathogens
8. TBCI in Farm and Companion Animals
9. Incidence Rate of TBCI in Humans and Impact of Co-Infections on Severity of Each Disease
10. Vaccines against TBDs for Human Use
11. Economic Impact of TBDs
12. Summary
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ASF | African swine fever |
| A. phagocytophilum | Anaplasma phagocytophilum |
| AMA1 | Apical membrane antigen 1 |
| ABRF | Argasidae-borne relapsing fever |
| Ba. microti | Babesia microti |
| BMSA | Babesia microti merozoites |
| Ba. bovis | Babesia bovis |
| Ba. divergens | Babesia divergens |
| B. afzelii | Borrelia afzelii |
| B. bavariensis | Borrelia bavariensis |
| B. burgdorferi | Borrelia burgdorferi |
| B. garinii | Borrelia garinii |
| B. mayonii | Borrelia mayonii |
| B. miyamotoi | Borrelia miyamotoi |
| B. valaisiana | Borrelia valaisiana |
| CDC | Centers for Disease Control and Prevention |
| Ech | Ehrlichia chaffeensis |
| EM | Erythema migrans |
| ECM | Extracellular matrix components |
| EUR | Euro |
| Hc | Hepatozoon canis |
| HGA | Human Granulocytic Anaplasmosis |
| HGE | Human Granulocytic Ehrlichiosis |
| hLFA-1 | Human lymphocyte function–associated antigen-1 |
| I. | Ixodes |
| LD | Lyme disease |
| MSA-2 | Merozoite surface antigen 2 |
| MAb | Monoclonal antibodies |
| Nm | Neoehrlichia mikurensis |
| NHP | Nonhuman primate |
| Osp | Outer surface protein |
| PCR | Polymerase chain reaction |
| PIM | Polymorphic immunodominant molecule |
| R. helvetica | Rickettsia helvetica |
| R. monacensis | Rickettsia monacensis |
| R. rickettsia | Rickettsia rickettsia |
| Rkt | Rickettsia spp. |
| RLB | Reverse Line Blot hybridization assay |
| RON2 | Rhoptry neck protein 2 |
| s.l. | sensu lato |
| spp. | Species |
| FDA | The United States Food and Drug Administration |
| USIC | The United States inflation calculator |
| US | The United States of America |
| TBI | Tick-borne infection |
| TBCI | Tick-borne co-infection |
| TBE | Tick-borne encephalitis |
| TBEV | Tick-borne encephalitis virus |
| TBD | Tick-borne disease |
| TBP | Tick-borne pathogen |
| TROSPA | Tick receptor for OspA |
| USD | Unites States dollar |
| UUK | Uukuniemi |
| VESA | Variant erythrocyte surface antigen |
| VlsE | Variable lipoprotein surface-exposed protein |
References
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, S3–S14. [Google Scholar] [CrossRef] [PubMed]
- Baneth, G. Tick-borne infections of animals and humans: A common ground. Int. J. Parasitol. 2014, 44, 591–596. [Google Scholar] [CrossRef]
- Madison-Antenucci, S.; Kramer, L.D.; Gebhardt, L.L.; Kauffman, E. Emerging Tick-Borne Diseases. Clin. Microbiol. Rev. 2020, 33, e00083-18. [Google Scholar] [CrossRef]
- Pritt, B.S.; Mead, P.S.; Johnson, D.K.H.; Neitzel, D.F.; Respicio-Kingry, L.B.; Davis, J.P.; Schiffman, E.; Sloan, L.M.; Schriefer, M.E.; Replogle, A.J.; et al. Identification of a novel pathogenic Borrelia species causing Lyme borreliosis with unusually high spirochaetaemia: A descriptive study. Lancet Infect. Dis. 2016, 16, 556–564. [Google Scholar] [CrossRef] [Green Version]
- CDC. Tickborne Disease Surveillance Data Summary. Available online: https://www.cdc.gov/ticks/data-summary/index.html (accessed on 18 April 2022).
- Kugeler, K.J.; Schwartz, A.M.; Delorey, M.J.; Mead, P.S.; Hinckley, A.F. Estimating the Frequency of Lyme Disease Diagnoses, United States, 2010–2018. Emerg. Infect. Dis. 2021, 27, 616–619. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, A.M.; Kugeler, K.J.; Nelson, C.A.; Marx, G.E.; Hinckley, A.F. Use of Commercial Claims Data for Evaluating Trends in Lyme Disease Diagnoses, United States, 2010–2018. Emerg. Infect. Dis. 2021, 27, 499–507. [Google Scholar] [CrossRef]
- Boyer, P.H.; Lenormand, C.; Jaulhac, B.; Talagrand-Reboul, E. Human Co-Infections between Borrelia burgdorferi s.L. and Other Ixodes-Borne Microorganisms: A Systematic Review. Pathogens 2022, 11, 282. [Google Scholar] [CrossRef]
- Benach, J.L.; Coleman, J.L.; Habicht, G.S.; MacDonald, A.; Grunwaldt, E.; Giron, J.A. Serological evidence for simultaneous occurrences of Lyme disease and babesiosis. J. Infect. Dis. 1985, 152, 473–477. [Google Scholar] [CrossRef]
- Moniuszko, A.; Dunaj, J.; Swiecicka, I.; Zambrowski, G.; Chmielewska-Badora, J.; Zukiewicz-Sobczak, W.; Zajkowska, J.; Czupryna, P.; Kondrusik, M.; Grygorczuk, S.; et al. Co-infections with Borrelia species, Anaplasma phagocytophilum and Babesia spp. in patients with tick-borne encephalitis. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1835–1841. [Google Scholar] [CrossRef] [Green Version]
- Velušček, M.; Blagus, R.; Kišek, T.C.; Ružić-Sabljić, E.; Avšič-Županc, T.; Bajrović, F.F.; Stupica, D. Antibiotic Use and Long-Term Outcome in Patients with Tick-Borne Encephalitis and Co-Infection with Borrelia Burgdorferi Sensu Lato in Central Europe. A Retrospective Cohort Study. J. Clin. Med. 2019, 8, 1740. [Google Scholar] [CrossRef]
- Lotric-Furlan, S.; Strle, F. Peripheral facial palsy in patients with tick-borne encephalitis. Clin. Microbiol. Infect. 2012, 18, 1027–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moutailler, S.; Valiente Moro, C.; Vaumourin, E.; Michelet, L.; Tran, F.H.; Devillers, E.; Cosson, J.F.; Gasqui, P.; Van, V.T.; Mavingui, P.; et al. Co-infection of Ticks: The Rule Rather Than the Exception. PLoS Negl. Trop. Dis. 2016, 10, e0004539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traavik, T. Experimental mixed CNS infections in mice caused by three Ixodes ricinus transmitted Arboviruses. Acta Pathol. Microbiol. Scand. B 1978, 86, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.F.; Johnson, R.C.; Magnarelli, L.A.; Hyde, F.W.; Myers, J.E. Peromyscus leucopus and Microtus pennsylvanicus simultaneously infected with Borrelia burgdorferi and Babesia microti. J. Clin. Microbiol. 1986, 23, 135–137. [Google Scholar] [CrossRef] [Green Version]
- Grunwaldt, E.; Barbour, A.G.; Benach, J.L. Simultaneous occurrence of babesiosis and Lyme disease. N. Engl. J. Med. 1983, 308, 1166. [Google Scholar] [CrossRef]
- Marcus, L.C.; Steere, A.C.; Duray, P.H.; Anderson, A.E.; Mahoney, E.B. Fatal pancarditis in a patient with coexistent Lyme disease and babesiosis. Demonstration of spirochetes in the myocardium. Ann. Intern. Med. 1985, 103, 374–376. [Google Scholar] [CrossRef]
- Steketee, R.W.; Eckman, M.R.; Burgess, E.C.; Kuritsky, J.N.; Dickerson, J.; Schell, W.L.; Godsey, M.S., Jr.; Davis, J.P. Babesiosis in Wisconsin. A new focus of disease transmission. JAMA 1985, 253, 2675–2678. [Google Scholar] [CrossRef]
- Kristoferitsch, W.; Stanek, G.; Kunz, C. Doppelinfektion mit Frühsommermeningoenzephalitis-(FSME-) Virus und Borrelia burgdorferi. DMW-Dtsch. Med. Wochenschr. 1986, 111, 861–864. [Google Scholar] [CrossRef]
- Duplaix, L.; Wagner, V.; Gasmi, S.; Lindsay, L.R.; Dibernardo, A.; Thivierge, K.; Fernandez-Prada, C.; Arsenault, J. Exposure to Tick-Borne Pathogens in Cats and Dogs Infested With Ixodes scapularis in Quebec: An 8-Year Surveillance Study. Front. Vet. Sci. 2021, 8, 696815. [Google Scholar] [CrossRef]
- Namina, A.; Capligina, V.; Seleznova, M.; Krumins, R.; Aleinikova, D.; Kivrane, A.; Akopjana, S.; Lazovska, M.; Berzina, I.; Ranka, R. Tick-borne pathogens in ticks collected from dogs, Latvia, 2011–2016. BMC Vet. Res. 2019, 15, 398. [Google Scholar] [CrossRef]
- Nimo-Paintsil, S.C.; Mosore, M.; Addo, S.O.; Lura, T.; Tagoe, J.; Ladzekpo, D.; Addae, C.; Bentil, R.E.; Behene, E.; Dafeamekpor, C.; et al. Ticks and prevalence of tick-borne pathogens from domestic animals in Ghana. Parasite Vectors 2022, 15, 86. [Google Scholar] [CrossRef]
- Hartlaub, J.; Daodu, O.B.; Sadeghi, B.; Keller, M.; Olopade, J.; Oluwayelu, D.; Groschup, M.H. Cross-Reaction or Co-Infection? Serological Discrimination of Antibodies Directed against Dugbe and Crimean-Congo Hemorrhagic Fever Orthonairovirus in Nigerian Cattle. Viruses 2021, 13, 1398. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Liu, Q.; Liu, J.Q.; Xu, B.L.; Lv, S.; Xia, S.; Zhou, X.N. Tick-borne pathogens and associated co-infections in ticks collected from domestic animals in central China. Parasite Vectors 2014, 7, 237. [Google Scholar] [CrossRef] [Green Version]
- Nieto, N.C.; Foley, J.E. Meta-analysis of coinfection and coexposure with Borrelia burgdorferi and Anaplasma phagocytophilum in humans, domestic animals, wildlife, and Ixodes ricinus-complex ticks. Vector Borne Zoonotic Dis. 2009, 9, 93–102. [Google Scholar] [CrossRef]
- Miranda, E.A.; Han, S.W.; Cho, Y.K.; Choi, K.S.; Chae, J.S. Co-Infection with Anaplasma Species and Novel Genetic Variants Detected in Cattle and Goats in the Republic of Korea. Pathogens 2021, 10, 28. [Google Scholar] [CrossRef]
- Colunga-Salas, P.; Sanchez-Montes, S.; Grostieta, E.; Verde-Arregoitia, L.D.; Cabrera-Garrido, M.Y.; Becker, I.; Leon-Paniagua, L. What do studies in wild mammals tell us about human emerging viral diseases in Mexico? Transbound. Emerg. Dis. 2020, 67, 33–45. [Google Scholar] [CrossRef]
- Borsan, S.D.; Ionica, A.M.; Galon, C.; Toma-Naic, A.; Pestean, C.; Sandor, A.D.; Moutailler, S.; Mihalca, A.D. High Diversity, Prevalence, and Co-infection Rates of Tick-Borne Pathogens in Ticks and Wildlife Hosts in an Urban Area in Romania. Front. Microbiol. 2021, 12, 645002. [Google Scholar] [CrossRef] [PubMed]
- Klompen, J.S.; Black, W.C.T.; Keirans, J.E.; Oliver, J.H., Jr. Evolution of ticks. Ann. Rev. Entomol. 1996, 41, 141–161. [Google Scholar] [CrossRef]
- Kazim, A.R.; Houssaini, J.; Ehlers, J.; Tappe, D.; Heo, C.C. Soft ticks (Acari: Argasidae) in the island nations of Southeast Asia: A review on their distribution, associated hosts and potential pathogens. Acta Trop. 2021, 223, 106085. [Google Scholar] [CrossRef] [PubMed]
- Talagrand-Reboul, E.; Boyer, P.H.; Bergstrom, S.; Vial, L.; Boulanger, N. Relapsing Fevers: Neglected Tick-Borne Diseases. Front. Cell. Infect. Microbiol. 2018, 8, 98. [Google Scholar] [CrossRef]
- Boinas, F.S.; Wilson, A.J.; Hutchings, G.H.; Martins, C.; Dixon, L.J. The persistence of African swine fever virus in field-infected Ornithodoros erraticus during the ASF endemic period in Portugal. PLoS ONE 2011, 6, e20383. [Google Scholar] [CrossRef]
- Lippi, C.A.; Ryan, S.J.; White, A.L.; Gaff, H.D.; Carlson, C.J. Trends and Opportunities in Tick-Borne Disease Geography. J. Med. Entomol. 2021, 58, 2021–2029. [Google Scholar] [CrossRef] [PubMed]
- Diuk-Wasser, M.A.; Vannier, E.; Krause, P.J. Coinfection by Ixodes Tick-Borne Pathogens: Ecological, Epidemiological, and Clinical Consequences. Trends Parasitol. 2016, 32, 30–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leger, E.; Vourc’h, G.; Vial, L.; Chevillon, C.; McCoy, K.D. Changing distributions of ticks: Causes and consequences. Exp. Appl. Acarol. 2013, 59, 219–244. [Google Scholar] [CrossRef]
- Randolph, S.E. Ticks are not Insects: Consequences of Contrasting Vector Biology for Transmission Potential. Parasitol. Today 1998, 14, 186–192. [Google Scholar] [CrossRef]
- Eisen, L. Pathogen transmission in relation to duration of attachment by Ixodes scapularis ticks. Ticks Tick Borne Dis. 2018, 9, 535–542. [Google Scholar] [CrossRef]
- Militzer, N.; Bartel, A.; Clausen, P.H.; Hoffmann-Kohler, P.; Nijhof, A.M. Artificial Feeding of All Consecutive Life Stages of Ixodes ricinus. Vaccines 2021, 9, 385. [Google Scholar] [CrossRef]
- Piesman, J.; Mather, T.N.; Donahue, J.G.; Levine, J.; Campbell, J.D.; Karakashian, S.J.; Spielman, A. Comparative prevalence of Babesia microti and Borrelia burgdorferi in four populations of Ixodes dammini in eastern Massachusetts. Acta Trop. 1986, 43, 263–270. [Google Scholar]
- Mather, T.N.; Telford, S.R., 3rd; Moore, S.I.; Spielman, A. Borrelia burgdorferi and Babesia microti: Efficiency of transmission from reservoirs to vector ticks (Ixodes dammini). Exp. Parasitol. 1990, 70, 55–61. [Google Scholar] [CrossRef]
- Mather, T.N.; Ginsberg, H.S. Vector-host-pathogen relationships: Transmission dynamics of tick-borne infections. In Ecological Dynamics of Tick-Borne Zoonoses; Oxford University Press: New York, NY, USA, 1994; pp. 68–90. [Google Scholar]
- Gomez-Chamorro, A.; Hodžić, A.; King, K.C.; Cabezas-Cruz, A. Ecological and evolutionary perspectives on tick-borne pathogen co-infections. Curr. Res. Parasitol. Vector-Borne Dis. 2021, 1, 100049. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.J.; Vayssier-Taussat, M.; Estrada-Peña, A.; Potkonjak, A.; Mihalca, A.D.; Zeller, H. Tick-borne diseases and co-infection: Current considerations. Ticks Tick-Borne Dis. 2021, 12, 101607. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Schauber, E.M.; Canham, C.D.; Keesing, F.; Jones, C.G.; Wolff, J.O. Effects of acorn production and mouse abundance on abundance and Borrelia burgdorferi infection prevalence of nymphal Ixodes scapularis ticks. Vector Borne Zoonotic Dis. 2001, 1, 55–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krawczyk, A.I.; van Duijvendijk, G.L.A.; Swart, A.; Heylen, D.; Jaarsma, R.I.; Jacobs, F.H.H.; Fonville, M.; Sprong, H.; Takken, W. Effect of rodent density on tick and tick-borne pathogen populations: Consequences for infectious disease risk. Parasite Vectors 2020, 13, 34. [Google Scholar] [CrossRef] [Green Version]
- Ueti, M.W.; Knowles, D.P.; Davitt, C.M.; Scoles, G.A.; Baszler, T.V.; Palmer, G.H. Quantitative differences in salivary pathogen load during tick transmission underlie strain-specific variation in transmission efficiency of Anaplasma marginale. Infect. Immun. 2009, 77, 70–75. [Google Scholar] [CrossRef] [Green Version]
- Chamberlain, R.W.; Corristan, E.C.; Sikes, R.K. Studies on the North American arthropod-borne encephalitides. V. The extrinsic incubation of eastern and western equine encephalitis in mosquitoes. Am. J. Hyg. 1954, 60, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, C.; Gern, L. Survival of Ixodes ricinus (Acari: Ixodidae) under challenging conditions of temperature and humidity is influenced by Borrelia burgdorferi sensu lato infection. J. Med. Entomol. 2010, 47, 1196–1204. [Google Scholar] [CrossRef]
- Ross, D.E.; Levin, M.L. Effects of Anaplasma phagocytophilum infection on the molting success of Ixodes scapularis (Acari: Ixodidae) larvae. J. Med. Entomol. 2004, 41, 476–483. [Google Scholar] [CrossRef] [Green Version]
- Ginsberg, H.S. Potential effects of mixed infections in ticks on transmission dynamics of pathogens: Comparative analysis of published records. Exp. Appl. Acarol. 2008, 46, 29–41. [Google Scholar] [CrossRef]
- Thomas, V.; Anguita, J.; Barthold, S.W.; Fikrig, E. Coinfection with Borrelia burgdorferi and the agent of human granulocytic ehrlichiosis alters murine immune responses, pathogen burden, and severity of Lyme arthritis. Infect. Immun. 2001, 69, 3359–3371. [Google Scholar] [CrossRef] [Green Version]
- Holden, K.; Hodzic, E.; Feng, S.; Freet, K.J.; Lefebvre, R.B.; Barthold, S.W. Coinfection with Anaplasma phagocytophilum alters Borrelia burgdorferi population distribution in C3H/HeN mice. Infect. Immun. 2005, 73, 3440–3444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klitgaard, K.; Kjaer, L.J.; Isbrand, A.; Hansen, M.F.; Bodker, R. Multiple infections in questing nymphs and adult female Ixodes ricinus ticks collected in a recreational forest in Denmark. Ticks Tick Borne Dis. 2019, 10, 1060–1065. [Google Scholar] [CrossRef]
- Lommano, E.; Bertaiola, L.; Dupasquier, C.; Gern, L. Infections and coinfections of questing Ixodes ricinus ticks by emerging zoonotic pathogens in Western Switzerland. Appl. Environ. Microbiol. 2012, 78, 4606–4612. [Google Scholar] [CrossRef] [Green Version]
- Capligina, V.; Seleznova, M.; Akopjana, S.; Freimane, L.; Lazovska, M.; Krumins, R.; Kivrane, A.; Namina, A.; Aleinikova, D.; Kimsis, J.; et al. Large-scale countrywide screening for tick-borne pathogens in field-collected ticks in Latvia during 2017–2019. Parasite Vectors 2020, 13, 351. [Google Scholar] [CrossRef] [PubMed]
- Reye, A.L.; Hubschen, J.M.; Sausy, A.; Muller, C.P. Prevalence and seasonality of tick-borne pathogens in questing Ixodes ricinus ticks from Luxembourg. Appl. Environ. Microbiol. 2010, 76, 2923–2931. [Google Scholar] [CrossRef] [Green Version]
- Lehane, A.; Maes, S.E.; Graham, C.B.; Jones, E.; Delorey, M.; Eisen, R.J. Prevalence of single and coinfections of human pathogens in Ixodes ticks from five geographical regions in the United States, 2013–2019. Ticks Tick Borne Dis. 2021, 12, 101637. [Google Scholar] [CrossRef] [PubMed]
- Magnarelli, L.A.; Stafford, K.C., 3rd; Ijdo, J.W.; Fikrig, E. Antibodies to whole-cell or recombinant antigens of Borrelia burgdorferi, Anaplasma phagocytophilum, and Babesia microti in white-footed mice. J. Wildl. Dis. 2006, 42, 732–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnarelli, L.A.; Anderson, J.F.; Stafford, K.C., 3rd; Dumler, J.S. Antibodies to multiple tick-borne pathogens of babesiosis, ehrlichiosis, and Lyme borreliosis in white-footed mice. J. Wildl. Dis. 1997, 33, 466–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, M.; Scherman, K.; Råberg, L. Infection dynamics of the tick-borne pathogen “Candidatus Neoehrlichia mikurensis” and coinfections with Borrelia afzelii in bank voles in Southern Sweden. Appl. Environ. Microbiol. 2014, 80, 1645–1649. [Google Scholar] [CrossRef] [Green Version]
- Hamšíková, Z.; Coipan, C.; Mahríková, L.; Minichová, L.; Sprong, H.; Kazimírová, M. Borrelia miyamotoi and Co-Infection with Borrelia afzelii in Ixodes ricinus Ticks and Rodents from Slovakia. Microb. Ecol. 2017, 73, 1000–1008. [Google Scholar] [CrossRef] [PubMed]
- Martello, E.; Mannelli, A.; Grego, E.; Ceballos, L.A.; Ragagli, C.; Stella, M.C.; Tomassone, L. Borrelia burgdorferi sensu lato and spotted fever group rickettsiae in small rodents and attached ticks in the Northern Apennines, Italy. Ticks Tick Borne Dis. 2019, 10, 862–867. [Google Scholar] [CrossRef]
- Hodzic, A.; Georges, I.; Postl, M.; Duscher, G.G.; Jeschke, D.; Szentiks, C.A.; Ansorge, H.; Heddergott, M. Molecular survey of tick-borne pathogens reveals a high prevalence and low genetic variability of Hepatozoon canis in free-ranging grey wolves (Canis lupus) in Germany. Ticks Tick Borne Dis. 2020, 11, 101389. [Google Scholar] [CrossRef]
- Mierzejewska, E.J.; Dwuznik, D.; Koczwarska, J.; Stanczak, L.; Opalinska, P.; Krokowska-Paluszak, M.; Wierzbicka, A.; Gorecki, G.; Bajer, A. The red fox (Vulpes vulpes), a possible reservoir of Babesia vulpes, B. canis and Hepatozoon canis and its association with the tick Dermacentor reticulatus occurrence. Ticks Tick Borne Dis. 2021, 12, 101551. [Google Scholar] [CrossRef]
- Beristain-Ruiz, D.M.; Vital-Garcia, C.; Figueroa-Millan, J.V.; Lira-Amaya, J.J.; Garza-Hernandez, J.A.; Sanchez-Ayala, J.R.; Flores-Ceballos, S.; Rodriguez-Alarcon, C.A.; Olivas-Sanchez, M.P.; Pons-Monarrez, G. Molecular Detection of Tick-Borne Pathogens in American Bison (Bison bison) at El Uno Ecological Reserve, Janos, Chihuahua, Mexico. Pathogens 2021, 10, 1428. [Google Scholar] [CrossRef]
- Overzier, E.; Pfister, K.; Herb, I.; Mahling, M.; Bock, G., Jr.; Silaghi, C. Detection of tick-borne pathogens in roe deer (Capreolus capreolus), in questing ticks (Ixodes ricinus), and in ticks infesting roe deer in southern Germany. Ticks Tick Borne Dis. 2013, 4, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Razanske, I.; Rosef, O.; Radzijevskaja, J.; Bratchikov, M.; Griciuviene, L.; Paulauskas, A. Prevalence and co-infection with tick-borne Anaplasma phagocytophilum and Babesia spp. in red deer (Cervus elaphus) and roe deer (Capreolus capreolus) in Southern Norway. Int. J. Parasitol. Parasites Wildl. 2019, 8, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Cafiso, A.; Bazzocchi, C.; Cavagna, M.; Di Lorenzo, E.; Serra, V.; Rossi, R.; Comazzi, S. Molecular Survey of Babesia spp. and Anaplasma phagocytophilum in Roe Deer from a Wildlife Rescue Center in Italy. Animals 2021, 11, 3335. [Google Scholar] [CrossRef]
- Johnson, N.; Golding, M.; Phipps, L.P. Detection of Tick-Borne Pathogens in Red Deer (Cervus elaphus), United Kingdom. Pathogens 2021, 10, 640. [Google Scholar] [CrossRef]
- Hersh, M.H.; Ostfeld, R.S.; McHenry, D.J.; Tibbetts, M.; Brunner, J.L.; Killilea, M.E.; LoGiudice, K.; Schmidt, K.A.; Keesing, F. Co-infection of blacklegged ticks with Babesia microti and Borrelia burgdorferi is higher than expected and acquired from small mammal hosts. PLoS ONE 2014, 9, e99348. [Google Scholar] [CrossRef]
- Milholland, M.T.; Xu, G.; Rich, S.M.; Machtinger, E.T.; Mullinax, J.M.; Li, A.Y. Pathogen Coinfections Harbored by Adult Ixodes scapularis from White-Tailed Deer Compared with Questing Adults Across Sites in Maryland, USA. Vector Borne Zoonotic Dis. 2021, 21, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.D.; Clark, K.L.; Coble, N.M.; Ballantyne, T.R. Detection and Transstadial Passage of Babesia Species and Borrelia burgdorferi Sensu Lato in Ticks Collected from Avian and Mammalian Hosts in Canada. Healthcare 2019, 7, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Fuente, J.; Naranjo, V.; Ruiz-Fons, F.; Vicente, J.; Estrada-Peña, A.; Almazán, C.; Kocan, K.M.; Martín, M.P.; Gortázar, C. Prevalence of tick-borne pathogens in ixodid ticks (Acari: Ixodidae) collected from European wild boar (Sus scrofa) and Iberian red deer (Cervus elaphus hispanicus) in central Spain. Eur. J. Wildl. Res. 2004, 50, 187–196. [Google Scholar] [CrossRef]
- Magalhaes-Matos, P.C.; Araujo, I.M.; Valim, J.R.A.; Ogrzewalska, M.; Guterres, A.; Cordeiro, M.D.; Cepeda, M.B.; Fonseca, A.H.D. Detection of Rickettsia spp. in ring-tailed coatis (Nasua nasua) and ticks of the Iguacu National Park, Brazilian Atlantic Rainforest. Ticks Tick Borne Dis. 2022, 13, 101891. [Google Scholar] [CrossRef] [PubMed]
- Willi, B.; Meli, M.L.; Cafarelli, C.; Gilli, U.O.; Kipar, A.; Hubbuch, A.; Riond, B.; Howard, J.; Schaarschmidt, D.; Regli, W.; et al. Cytauxzoon europaeus infections in domestic cats in Switzerland and in European wildcats in France: A tale that started more than two decades ago. Parasite Vectors 2022, 15, 19. [Google Scholar] [CrossRef] [PubMed]
- Egan, S.L.; Taylor, C.L.; Banks, P.B.; Northover, A.S.; Ahlstrom, L.A.; Ryan, U.M.; Irwin, P.J.; Oskam, C.L. The bacterial biome of ticks and their wildlife hosts at the urban-wildland interface. Microb. Genom. 2021, 7, 000730. [Google Scholar] [CrossRef]
- Masika, S.J.; Muchemi, G.M.; Okumu, T.A.; Mutura, S.; Zimmerman, D.; Kamau, J. Molecular evidence of Anaplasma phagocytophilum in olive baboons and vervet monkeys in Kenya. BMC Vet. Res. 2021, 17, 385. [Google Scholar] [CrossRef]
- Zhao, H.Q.; Liu, P.P.; Xue, F.; Lu, M.; Qin, X.C.; Li, K. Molecular Detection and Identification of Candidatus Ehrlichia hainanensis, A Novel Ehrlichia Species in Rodents from Hainan Province, China. Biomed. Environ. Sci. 2021, 34, 1020–1023. [Google Scholar] [CrossRef]
- Telfer, S.; Lambin, X.; Birtles, R.; Beldomenico, P.; Burthe, S.; Paterson, S.; Begon, M. Species interactions in a parasite community drive infection risk in a wildlife population. Science 2010, 330, 243–246. [Google Scholar] [CrossRef] [Green Version]
- Hersh, M.H.; Tibbetts, M.; Strauss, M.; Ostfeld, R.S.; Keesing, F. Reservoir Competence of Wildlife Host Species for Babesia microti. Emerg. Infect. Dis. 2012, 18, 1951–1957. [Google Scholar] [CrossRef]
- Keesing, F.; Hersh, M.H.; Tibbetts, M.; McHenry, D.J.; Duerr, S.; Brunner, J.; Killilea, M.; LoGiudice, K.; Schmidt, K.A.; Ostfeld, R.S. Reservoir Competence of Vertebrate Hosts for Anaplasma phagocytophilum. Emerg. Infect. Dis. 2012, 18, 2013–2016. [Google Scholar] [CrossRef] [PubMed]
- Boni, L.D. Occurrence of Tick-Borne Haemoparasites in Selected South African Wild Rodent Species in the Mnisi Communal Area. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 2017. [Google Scholar]
- Levi, T.; Kilpatrick, A.M.; Mangel, M.; Wilmers, C.C. Deer, predators, and the emergence of Lyme disease. Proc. Natl. Acad. Sci. USA 2012, 109, 10942–10947. [Google Scholar] [CrossRef] [Green Version]
- Campbell, T.A.; VerCauteren, K.C. Diseases and Parasites [of White-tailed Deer]. In Biology and Management of White-Tailed Deer; Hewitt, D.G., Ed.; CRC Press: Boca Raton, FL, USA, 2011; pp. 219–249. [Google Scholar]
- Ruiz-Fons, F.; Fernandez-de-Mera, I.G.; Acevedo, P.; Gortazar, C.; de la Fuente, J. Factors driving the abundance of Ixodes ricinus ticks and the prevalence of zoonotic I. ricinus-borne pathogens in natural foci. Appl. Environ. Microbiol. 2012, 78, 2669–2676. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Pena, A.; George, J.C.; Golovljova, I.; Jaenson, T.G.; Jensen, J.K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasite Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasle, G. Transport of ixodid ticks and tick-borne pathogens by migratory birds. Front. Cell. Infect. Microbiol. 2013, 3, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson, S.J.; Neitzel, D.; Reed, K.D.; Belongia, E.A. Coinfections acquired from Ixodes ticks. Clin. Microbiol. Rev. 2006, 19, 708–727. [Google Scholar] [CrossRef] [Green Version]
- Bumann, D. Pathogen proteomes during infection: A basis for infection research and novel control strategies. J. Proteom. 2010, 73, 2267–2276. [Google Scholar] [CrossRef]
- Marcelino, I.; de Almeida, A.M.; Ventosa, M.; Pruneau, L.; Meyer, D.F.; Martinez, D.; Lefrancois, T.; Vachiery, N.; Coelho, A.V. Tick-borne diseases in cattle: Applications of proteomics to develop new generation vaccines. J. Proteom. 2012, 75, 4232–4250. [Google Scholar] [CrossRef] [PubMed]
- Toye, P.; Musoke, A.; Naessens, J. Role of the polymorphic immunodominant molecule in entry of Theileria parva sporozoites into bovine lymphocytes. Infect. Immun. 2014, 82, 1786–1792. [Google Scholar] [CrossRef] [Green Version]
- Dobbelaere, D.A.E.; Spooner, P.R.; Barry, W.C.; Irvin, A.D. Monoclonal-Antibody Neutralizes the Sporozoite Stage of Different Theileria-Parva Stocks. Parasite Immunol. 1984, 6, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Knight, P.; Musoke, A.J.; Gachanja, J.N.; Nene, V.; Katzer, F.; Boulter, N.; Hall, R.; Brown, C.G.D.; Williamson, S.; Kirvar, E.; et al. Conservation of neutralizing determinants between the sporozoite surface antigens of Theileria annulata and Theileria parva. Exp. Parasitol. 1996, 82, 229–241. [Google Scholar] [CrossRef]
- Ijdo, J.W.; Carlson, A.C.; Kennedy, E.L. Anaplasma phagocytophilum AnkA is tyrosine-phosphorylated at EPIYA motifs and recruits SHP-1 during early infection. Cell. Microbiol. 2007, 9, 1284–1296. [Google Scholar] [CrossRef]
- Yang, X.L.; Promnares, K.; Qin, J.H.; He, M.; Shroder, D.Y.; Kariu, T.; Wang, Y.; Pal, U. Characterization of Multiprotein Complexes of the Borrelia burgdorferi Outer Membrane Vesicles. J. Proteome Res. 2011, 10, 4556–4566. [Google Scholar] [CrossRef] [PubMed]
- Kenedy, M.R.; Lenhart, T.R.; Akins, D.R. The role of Borrelia burgdorferi outer surface proteins. FEMS Immunol. Med. Microbiol. 2012, 66, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.F.F.; Pal, U.; Alani, S.M.; Fikrig, E.; Norgard, M.V. Essential role for OspA/B in the life cycle of the Lyme disease spirochete. J. Exp. Med. 2004, 199, 641–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, U.; Li, X.; Wang, T.; Montgomery, R.R.; Ramamoorthi, N.; Desilva, A.M.; Bao, F.; Yang, X.; Pypaert, M.; Pradhan, D.; et al. TROSPA, an Ixodes scapularis receptor for Borrelia burgdorferi. Cell 2004, 119, 457–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilly, K.; Krum, J.G.; Bestor, A.; Jewett, M.W.; Grimm, D.; Bueschel, D.; Byram, R.; Dorward, D.; VanRaden, M.J.; Stewart, P.; et al. Borrelia burgdorferi OspC protein required exclusively in a crucial early stage of mammalian infection. Infect. Immun. 2006, 74, 3554–3564. [Google Scholar] [CrossRef] [Green Version]
- Onder, O.; Humphrey, P.T.; McOmber, B.; Korobova, F.; Francella, N.; Greenbaum, D.C.; Brisson, D. OspC Is Potent Plasminogen Receptor on Surface of Borrelia burgdorferi. J. Biol. Chem. 2012, 287, 16860–16868. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.R.; Hardham, J.M.; Barbour, A.G.; Norris, S.J. Antigenic variation in Lyme disease borreliae by promiscuous recombination of VMP-like sequence cassettes. Cell 1997, 89, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Skare, J.T.; Mirzabekov, T.A.; Shang, E.S.; Blanco, D.R.; ErdjumentBromage, H.; Bunikis, J.; Bergstrom, S.; Tempst, P.; Kagan, B.L.; Miller, J.N.; et al. The Oms66 (p66) protein is a Borrelia burgdorferi porin. Infect. Immun. 1997, 65, 3654–3661. [Google Scholar] [CrossRef] [Green Version]
- Noppa, L.; Ostberg, Y.; Lavrinovicha, M.; Bergstrom, S. P13, an integral membrane protein of Borrelia burgdorferi, is C-terminally processed and contains surface-exposed domains. Infect. Immun. 2001, 69, 3323–3334. [Google Scholar] [CrossRef] [Green Version]
- Coburn, J.; Leong, J.M.; Erban, J.K. Integrin alpha IIb beta 3 mediates binding of the Lyme disease agent Borrelia burgdorferi to human platelets. Proc. Natl. Acad. Sci. USA 1993, 90, 7059–7063. [Google Scholar] [CrossRef] [Green Version]
- Guo, B.P.; Brown, E.L.; Dorward, D.W.; Rosenberg, L.C.; Höök, M. Decorin-binding adhesins from Borrelia burgdorferi. Mol. Microbiol. 1998, 30, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Parveen, N.; Caimano, M.; Radolf, J.D.; Leong, J.M. Adaptation of the Lyme disease spirochaete to the mammalian host environment results in enhanced glycosaminoglycan and host cell binding. Mol. Microbiol. 2003, 47, 1433–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parveen, N.; Leong, J.M. Identification of a candidate glycosaminoglycan-binding adhesin of the Lyme disease spirochete Borrelia burgdorferi. Mol. Microbiol. 2000, 35, 1220–1234. [Google Scholar] [CrossRef]
- Fischer, J.R.; Parveen, N.; Magoun, L.; Leong, J.M. Decorin-binding proteins A and B confer distinct mammalian cell type-specific attachment by Borrelia burgdorferi, the Lyme disease spirochete. Proc. Natl. Acad. Sci. USA 2003, 100, 7307–7312. [Google Scholar] [CrossRef] [Green Version]
- Fischer, J.R.; LeBlanc, K.T.; Leong, J.M. Fibronectin binding protein BBK32 of the Lyme disease spirochete promotes bacterial attachment to glycosaminoglycans. Infect. Immun. 2006, 74, 435–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutten, E.L.; Norimine, J.; Beare, P.A.; Heinzen, R.A.; Lopez, J.E.; Morse, K.; Brayton, K.A.; Gillespie, J.J.; Brown, W.C. Anaplasma marginale Type IV Secretion System Proteins VirB2, VirB7, VirB11, and VirD4 Are Immunogenic Components of a Protective Bacterial Membrane Vaccine. Infect. Immun. 2010, 78, 1314–1325. [Google Scholar] [CrossRef] [Green Version]
- Rikihisa, Y.; Lin, M.Q.; Niu, H.; Cheng, Z.H. Type IV Secretion System of Anaplasma phagocytophilum and Ehrlichia chaffeensis. Ann. N. Y. Acad. Sci. 2009, 1166, 106–111. [Google Scholar] [CrossRef]
- Allred, D.R.; Al-Khedery, B. Antigenic variation as an exploitable weakness of babesial parasites. Vet. Parasitol. 2006, 138, 50–60. [Google Scholar] [CrossRef]
- Allred, D.R.; Carlton, J.M.R.; Satcher, R.L.; Long, J.A.; Brown, W.C.; Patterson, P.E.; O’Connor, R.M.; Stroup, S.E. The ves multigene family of B-bovis encodes components of rapid antigenic variation at the infected erythrocyte surface. Mol. Cell. 2000, 5, 153–162. [Google Scholar] [CrossRef]
- Mosqueda, J.; McElwain, T.F.; Palmer, G.H. Babesia bovis merozoite surface antigen 2 proteins are expressed on the merozoite and sporozoite surface, and specific antibodies inhibit attachment and invasion of erythrocytes. Infect. Immun. 2002, 70, 6448–6455. [Google Scholar] [CrossRef] [Green Version]
- Man, S.Q.; Fu, Y.F.; Guan, Y.; Feng, M.; Qiao, K.; Li, X.P.; Gao, H.J.; Cheng, X.J. Evaluation of a Major Surface Antigen of Babesia microti Merozoites as a Vaccine Candidate against Babesia Infection. Front. Microbiol. 2017, 8, 2545. [Google Scholar] [CrossRef] [PubMed]
- Tyler, J.S.; Treeck, M.; Boothroyd, J.C. Focus on the ringleader: The role of AMA1 in apicomplexan invasion and replication. Trends Parasitol. 2011, 27, 410–420. [Google Scholar] [CrossRef] [Green Version]
- Barthold, S.W.; Beck, D.S.; Hansen, G.M.; Terwilliger, G.A.; Moody, K.D. Lyme Borreliosis in Selected Strains and Ages of Laboratory Mice. J. Infect. Dis. 1990, 162, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Seiler, K.P.; Eichwald, E.J.; Weis, J.H.; Teuscher, C.; Weis, J.J. Distinct Characteristics of Resistance to Borrelia burgdorferi Induced Arthritis in C57BL/6N Mice. Infect. Immun. 1998, 66, 161–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barthold, S.W.; Sidman, C.L.; Smith, A.L. Lyme Borreliosis in Genetically Resistant and Susceptible Mice with Severe Combined Immunodeficiency. Am. J. Trop. Med. Hyg. 1992, 47, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Lynn, G.E.; Breuner, N.E.; Hojgaard, A.; Oliver, J.; Eisen, L.; Eisen, R.J. A comparison of horizontal and transovarial transmission efficiency of Borrelia miyamotoi by Ixodes scapularis. Ticks Tick-Borne Dis. 2022, 13, 102003. [Google Scholar] [CrossRef]
- Breuner, N.E.; Dolan, M.C.; Replogle, A.J.; Sexton, C.; Hojgaard, A.; Boegler, K.A.; Clark, R.J.; Eisen, L. Transmission of Borrelia miyamotoi sensu lato relapsing fever group spirochetes in relation to duration of attachment by Ixodes scapularis nymphs. Ticks Tick Borne Dis. 2017, 8, 677–681. [Google Scholar] [CrossRef]
- Breuner, N.E.; Hojgaard, A.; Replogle, A.J.; Boegler, K.A.; Eisen, L. Transmission of the relapsing fever spirochete, Borrelia miyamotoi, by single transovarially-infected larval Ixodes scapularis ticks. Ticks Tick Borne Dis. 2018, 9, 1464–1467. [Google Scholar] [CrossRef]
- Campfield, B.T.; Nolder, C.L.; Davis, A.; Hirsch, R.; Nowalk, A.J. The DBA/1 strain is a novel mouse model for experimental Borrelia burgdorferi infection. Clin. Vaccine Immunol. 2012, 19, 1567–1573. [Google Scholar] [CrossRef]
- Casselli, T.; Divan, A.; Vomhof-DeKrey, E.E.; Tourand, Y.; Pecoraro, H.L.; Brissette, C.A. A murine model of Lyme disease demonstrates that Borrelia burgdorferi colonizes the dura mater and induces inflammation in the central nervous system. PLoS Pathog. 2021, 17, e1009256. [Google Scholar] [CrossRef]
- Schmitz, J.L.; Schell, R.F.; Hejka, A.; England, D.M.; Konick, L. Induction of lyme arthritis in LSH hamsters. Infect. Immun. 1988, 56, 2336–2342. [Google Scholar] [CrossRef] [PubMed]
- Goodman, J.L.; Jurkovich, P.; Kodner, C.; Johnson, R.C. Persistent cardiac and urinary tract infections with Borrelia burgdorferi in experimentally infected Syrian hamsters. J. Clin. Microbiol. 1991, 29, 894–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley, D.M.; Gayek, R.J.; Skare, J.T.; Wagar, E.A.; Champion, C.I.; Blanco, D.R.; Lovett, M.A.; Miller, J.N. Rabbit model of Lyme borreliosis: Erythema migrans, infection-derived immunity, and identification of Borrelia burgdorferi proteins associated with virulence and protective immunity. J. Clin. Investig. 1995, 96, 965–975. [Google Scholar] [CrossRef] [Green Version]
- Embers, M.E.; Hasenkampf, N.R.; Jacobs, M.B.; Tardo, A.C.; Doyle-Meyers, L.A.; Philipp, M.T.; Hodzic, E. Variable manifestations, diverse seroreactivity and post-treatment persistence in non-human primates exposed to Borrelia burgdorferi by tick feeding. PLoS ONE 2017, 12, e0189071. [Google Scholar] [CrossRef] [Green Version]
- Philipp, M.T.; Aydintug, M.K.; Bohm, R.P., Jr.; Cogswell, F.B.; Dennis, V.A.; Lanners, H.N.; Lowrie, R.C., Jr.; Roberts, E.D.; Conway, M.D.; Karaçorlu, M.; et al. Early and early disseminated phases of Lyme disease in the rhesus monkey: A model for infection in humans. Infect. Immun. 1993, 61, 3047–3059. [Google Scholar] [CrossRef] [Green Version]
- Roberts, E.D.; Bohm, R.P., Jr.; Cogswell, F.B.; Lanners, H.N.; Lowrie, R.C., Jr.; Povinelli, L.; Piesman, J.; Philipp, M.T. Chronic lyme disease in the rhesus monkey. Lab. Invest. 1995, 72, 146–160. [Google Scholar] [PubMed]
- England, J.D.; Bohm, R.P., Jr.; Roberts, E.D.; Philipp, M.T. Lyme neuroborreliosis in the rhesus monkey. Semin. Neurol. 1997, 17, 53–56. [Google Scholar] [CrossRef]
- Pachner, A.R.; Amemiya, K.; Bartlett, M.; Schaefer, H.; Reddy, K.; Zhang, W.F. Lyme borreliosis in rhesus macaques: Effects of corticosteroids on spirochetal load and isotype switching of anti-borrelia burgdorferi antibody. Clin. Diagn. Lab. Immunol. 2001, 8, 225–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borggraefe, I.; Yuan, J.; Telford, S.R.; Menon, S.; Hunter, R.; Shah, S.; Spielman, A.; Gelfand, J.A.; Wortis, H.H.; Vannier, E. Babesia microti Primarily Invades Mature Erythrocytes in Mice. Infect. Immun. 2006, 74, 3204–3212. [Google Scholar] [CrossRef] [Green Version]
- Vannier, E.; Borggraefe, I.; Telford, S.R., 3rd; Menon, S.; Brauns, T.; Spielman, A.; Gelfand, J.A.; Wortis, H.H. Age-associated decline in resistance to Babesia microti is genetically determined. J. Infect. Dis. 2004, 189, 1721–1728. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.A.; Kuntz, R.E. Babesia microti Infections in Nonhuman Primates. J. Parasitol. 1981, 67, 454–456. [Google Scholar] [CrossRef] [PubMed]
- Ruebush, T.K., 2nd; Piesman, J.; Collins, W.E.; Spielman, A.; Warren, M. Tick transmission of Babesia microti to rhesus monkeys (Macaca mulatta). Am. J. Trop. Med. Hyg. 1981, 30, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Ruebush, T.K.; Collins, W.E.; Warren, M. Experimental Babesia microti infections in Macaca mulatta: Recurrent parasitemia before and after splenectomy. Am. J. Trop. Med. Hyg. 1981, 30, 304–307. [Google Scholar] [CrossRef] [PubMed]
- Gumber, S.; Nascimento, F.S.; Rogers, K.A.; Bishop, H.S.; Rivera, H.N.; Xayavong, M.V.; Devare, S.G.; Schochetman, G.; Amancha, P.K.; Qvarnstrom, Y.; et al. Experimental transfusion-induced Babesia microti infection: Dynamics of parasitemia and immune responses in a rhesus macaque model. Transfusion 2016, 56, 1508–1519. [Google Scholar] [CrossRef] [Green Version]
- Kocan, K.M.; Busby, A.T.; Allison, R.W.; Breshears, M.A.; Coburn, L.; Galindo, R.C.; Ayllon, N.; Blouin, E.F.; de la Fuente, J. Sheep experimentally infected with a human isolate of Anaplasma phagocytophilum serve as a host for infection of Ixodes scapularis ticks. Ticks Tick Borne Dis. 2012, 3, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Reppert, E.; Galindo, R.C.; Ayllon, N.; Breshears, M.A.; Kocan, K.M.; Blouin, E.F.; de la Fuente, J. Studies of Anaplasma phagocytophilum in sheep experimentally infected with the human NY-18 isolate: Characterization of tick feeding sites. Ticks Tick Borne Dis. 2014, 5, 744–752. [Google Scholar] [CrossRef]
- Urbanova, V.; Kalinova, E.; Kopacek, P.; Sima, R. Experimental Infection of Mice and Ticks with the Human Isolate of Anaplasma phagocytophilum NY-18. Pathogens 2022, 11, 820. [Google Scholar] [CrossRef]
- Naimi, W.A.; Green, R.S.; Cockburn, C.L.; Carlyon, J.A. Differential Susceptibility of Male Versus Female Laboratory Mice to Anaplasma phagocytophilum Infection. Trop. Med. Infect. Dis. 2018, 3, 78. [Google Scholar] [CrossRef] [Green Version]
- Telford, S.R., 3rd; Dawson, J.E.; Katavolos, P.; Warner, C.K.; Kolbert, C.P.; Persing, D.H. Perpetuation of the agent of human granulocytic ehrlichiosis in a deer tick-rodent cycle. Proc. Natl. Acad. Sci. USA 1996, 93, 6209–6214. [Google Scholar] [CrossRef] [Green Version]
- Hodzic, E.; IJdo, J.W.I.; Feng, S.; Katavolos, P.; Sun, W.; Maretzki, C.H.; Fish, D.; Fikrig, E.; Telford, S.R., III.; Barthold, S.W. Granulocytic Ehrlichiosis in the Laboratory Mouse. J. Infect. Dis. 1998, 177, 737–745. [Google Scholar] [CrossRef] [Green Version]
- Bunnell, J.E.; Magnarelli, L.A.; Dumler, J.S. Infection of Laboratory Mice with the Human Granulocytic Ehrlichiosis Agent Does Not Induce Antibodies to Diagnostically Significant Borrelia burgdorferi Antigens. J. Clin. Microbiol. 1999, 37, 2077–2079. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.D.; Cheng, C.; Jaworski, D.C.; Willard, L.H.; Sanderson, M.W.; Ganta, R.R. Ehrlichia chaffeensis infection in the reservoir host (white-tailed deer) and in an incidental host (dog) is impacted by its prior growth in macrophage and tick cell environments. PLoS ONE 2014, 9, e109056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbott, R.C.; Chomel, B.B.; Kasten, R.W.; Floyd-Hawkins, K.A.; Kikuchi, Y.; Koehler, J.E.; Pedersen, N.C. Experimental and natural infection with Bartonella henselae in domestic cats. Comp. Immunol. Microbiol. Infect. Dis. 1997, 20, 41–51. [Google Scholar] [CrossRef]
- O’Reilly, K.L.; Bauer, R.W.; Freeland, R.L.; Foil, L.D.; Hughes, K.J.; Rohde, K.R.; Roy, A.F.; Stout, R.W.; Triche, P.C. Acute Clinical Disease in Cats following Infection with a Pathogenic Strain of Bartonella henselae (LSU16). Infect. Immun. 1999, 67, 3066–3072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, M.N.; Vieira-Damiani, G.; Ericson, M.E.; Gupta, K.; de Almeida, A.R.; Drummond, M.R.; Soares, T.C.; Lania, B.G.; Gilioli, R.; Velho, P.E. Acute and Late Bartonella henselae Murine Model Infection. Vector Borne Zoonotic Dis. 2017, 17, 206–208. [Google Scholar] [CrossRef] [Green Version]
- Siewert, L.K.; Dehio, C.; Pinschewer, D.D. Adaptive immune defense prevents Bartonella persistence upon trans-placental transmission. PLoS Pathog. 2022, 18, e1010489. [Google Scholar] [CrossRef] [PubMed]
- Chiaraviglio, L.; Duong, S.; Brown, D.A.; Birtles, R.J.; Kirby, J.E. An Immunocompromised Murine Model of Chronic Bartonella Infection. Am. J. Pathol. 2010, 176, 2753–2763. [Google Scholar] [CrossRef]
- Boulanger, N.; Wikel, S. Induced Transient Immune Tolerance in Ticks and Vertebrate Host: A Keystone of Tick-Borne Diseases? Front. Immunol. 2021, 12, 625993. [Google Scholar] [CrossRef]
- Belongia, E.A. Epidemiology and impact of coinfections acquired from Ixodes ticks. Vector Borne Zoonotic Dis. 2002, 2, 265–273. [Google Scholar] [CrossRef]
- Djokic, V.; Akoolo, L.; Primus, S.; Schlachter, S.; Kelly, K.; Bhanot, P.; Parveen, N. Protozoan Parasite Babesia microti Subverts Adaptive Immunity and Enhances Lyme Disease Severity. Front. Microbiol. 2019, 10, 1596. [Google Scholar] [CrossRef]
- Djokic, V.; Primus, S.; Akoolo, L.; Chakraborti, M.; Parveen, N. Age-Related Differential Stimulation of Immune Response by Babesia microti and Borrelia burgdorferi During Acute Phase of Infection Affects Disease Severity. Front. Immunol. 2018, 9, 2891. [Google Scholar] [CrossRef] [PubMed]
- Akoolo, L.; Djokic, V.; Rocha, S.C.; Parveen, N. Pathogenesis of Borrelia burgdorferi and Babesia microti in TLR4-Competent and TLR4-dysfunctional C3H mice. Cell. Microbiol. 2021, 23, e13350. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.L.; LeVine, D.; Thill, C.; Kuhlow, C.; Benach, J.L. Babesia microti and Borrelia burgdorferi follow independent courses of infection in mice. J. Infect. Dis. 2005, 192, 1634–1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moro, M.H.; Zegarra-Moro, O.L.; Bjornsson, J.; Hofmeister, E.K.; Bruinsma, E.; Germer, J.J.; Persing, D.H. Increased Arthritis Severity in Mice Coinfected with Borrelia burgdorferi and Babesia microti. J. Infect. Dis. 2002, 186, 428–431. [Google Scholar] [CrossRef] [Green Version]
- Bhanot, P.; Parveen, N. Investigating disease severity in an animal model of concurrent babesiosis and Lyme disease. Int. J. Parasitol. 2019, 49, 145–151. [Google Scholar] [CrossRef]
- Zeidner, N.S.; Dolan, M.C.; Massung, R.; Piesman, J.; Fish, D. Coinfection with Borrelia burgdorferi and the agent of human granulocytic ehrlichiosis suppresses IL-2 and IFN gamma production and promotes an IL-4 response in C3H/HeJ mice. Parasite Immunol. 2000, 22, 581–588. [Google Scholar] [CrossRef]
- De la Fuente, J.; Contreras, M.; Estrada-Peña, A.; Cabezas-Cruz, A. Targeting a global health problem: Vaccine design and challenges for the control of tick-borne diseases. Vaccine 2017, 35, 5089–5094. [Google Scholar] [CrossRef]
- Merino, O.; Alberdi, P.; de la Lastra, J.M.P.; de la Fuente, J. Tick vaccines and the control of tick-borne pathogens. Front. Cell. Infect. Microbiol. 2013, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Opdebeeck, J.P.; Wong, J.Y.; Jackson, L.A.; Dobson, C. Vaccines to protect Hereford cattle against the cattle tick, Boophilus microplus. Immunology 1988, 63, 363–367. [Google Scholar]
- Carreón, D.; de la Lastra, J.M.; Almazán, C.; Canales, M.; Ruiz-Fons, F.; Boadella, M.; Moreno-Cid, J.A.; Villar, M.; Gortázar, C.; Reglero, M.; et al. Vaccination with BM86, subolesin and akirin protective antigens for the control of tick infestations in white tailed deer and red deer. Vaccine 2012, 30, 273–279. [Google Scholar] [CrossRef]
- De la Fuente, J.; Rodriguez, M.; Montero, C.; Redondo, M.; Garcia-Garcia, J.C.; Mendez, L.; Serrano, E.; Valdes, M.; Enriquez, A.; Canales, M.; et al. Vaccination against ticks (Boophilus spp.): The experience with the Bm86-based vaccine Gavac. Genet. Anal. 1999, 15, 143–148. [Google Scholar] [CrossRef]
- Gomes-Solecki, M. Blocking pathogen transmission at the source: Reservoir targeted OspA-based vaccines against Borrelia burgdorferi. Front. Cell. Infect. Microbiol. 2014, 4, 136. [Google Scholar] [CrossRef]
- Richer, L.M.; Brisson, D.; Melo, R.; Ostfeld, R.S.; Zeidner, N.; Gomes-Solecki, M. Reservoir Targeted Vaccine Against Borrelia burgdorferi: A New Strategy to Prevent Lyme Disease Transmission. J. Infect. Dis. 2014, 209, 1972–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luke, C.J.; Carner, K.; Liang, X.; Barbour, A.G. An OspA-based DNA vaccine protects mice against infection with Borrelia burgdorferi. J. Infect. Dis. 1997, 175, 91–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sajid, A.; Matias, J.; Arora, G.; Kurokawa, C.; DePonte, K.; Tang, X.; Lynn, G.; Wu, M.-J.; Pal, U.; Strank, N.O.; et al. mRNA vaccination induces tick resistance and prevents transmission of the Lyme disease agent. Sci. Transl. Med. 2021, 13, eabj9827. [Google Scholar] [CrossRef] [PubMed]
- VanBlargan, L.A.; Himansu, S.; Foreman, B.M.; Ebel, G.D.; Pierson, T.C.; Diamond, M.S. An mRNA Vaccine Protects Mice against Multiple Tick-Transmitted Flavivirus Infections. Cell. Rep. 2018, 25, 3382–3392. [Google Scholar] [CrossRef] [Green Version]
- Gehrke, R.; Ecker, M.; Aberle, S.W.; Allison, S.L.; Heinz, F.X.; Mandl, C.W. Incorporation of tick-borne encephalitis virus replicons into virus-like particles by a packaging cell line. J. Virol. 2003, 77, 8924–8933. [Google Scholar] [CrossRef] [Green Version]
- Estrada-Peña, A.; de la Fuente, J.; Ostfeld, R.S.; Cabezas-Cruz, A. Interactions between tick and transmitted pathogens evolved to minimise competition through nested and coherent networks. Sci. Rep. 2015, 5, 10361. [Google Scholar] [CrossRef] [Green Version]
- Estrada-Peña, A.; Sprong, H.; Cabezas-Cruz, A.; de la Fuente, J.; Ramo, A.; Coipan, E.C. Nested coevolutionary networks shape the ecological relationships of ticks, hosts, and the Lyme disease bacteria of the Borrelia burgdorferi (s.L.) complex. Parasites Vectors 2016, 9, 517. [Google Scholar] [CrossRef] [Green Version]
- Randolph, S.E.; Gern, L.; Nuttall, P.A. Co-feeding ticks: Epidemiological significance for tick-borne pathogen transmission. Parasitol. Today 1996, 12, 472–479. [Google Scholar] [CrossRef]
- Contreras, M.; de la Fuente, J. Control of infestations by Ixodes ricinus tick larvae in rabbits vaccinated with aquaporin recombinant antigens. Vaccine 2017, 35, 1323–1328. [Google Scholar] [CrossRef] [PubMed]
- Salih, D.; El Hussein, A.; Singla, L. Diagnostic approaches for tick-borne haemoparasitic diseases in livestock. J. Vet. Med. Anim. Health 2015, 7, 45–56. [Google Scholar]
- Beugnet, F.; Marie, J.L. Emerging arthropod-borne diseases of companion animals in Europe. Vet. Parasitol. 2009, 163, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Bajer, A.; Rodo, A.; Bednarska, M.; Mierzejewska, E.; Welc-Faleciak, R. Babesia canis and tick-borne encephalitis virus (TBEV) co-infection in a sled dog. Ann. Agric. Environ. Med. 2013, 20, 426–430. [Google Scholar]
- Espinaze, M.P.A.; Hellard, E.; Horak, I.G.; Cumming, G.S. Domestic mammals facilitate tick-borne pathogen transmission networks in South African wildlife. Biol. Conserv. 2018, 221, 228–236. [Google Scholar] [CrossRef] [Green Version]
- Wyk, C.-L.v.; Mtshali, K.; Taioe, M.O.; Terera, S.; Bakkes, D.; Ramatla, T.; Xuan, X.; Thekisoe, O. Detection of Ticks and Tick-Borne Pathogens of Urban Stray Dogs in South Africa. Pathogens 2022, 11, 862. [Google Scholar] [CrossRef]
- Kolo, A.O.; Sibeko-Matjila, K.P.; Maina, A.N.; Richards, A.L.; Knobel, D.L.; Matjila, P.T. Molecular Detection of Zoonotic Rickettsiae and Anaplasma spp. in Domestic Dogs and Their Ectoparasites in Bushbuckridge, South Africa. Vector Borne Zoonotic Dis. 2016, 16, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Woldehiwet, Z. Anaplasma phagocytophilum in ruminants in Europe. Ann. N. Y. Acad. Sci. 2006, 1078, 446–460. [Google Scholar] [CrossRef]
- Andersson, M.O.; Vichova, B.; Tolf, C.; Krzyzanowska, S.; Waldenstrom, J.; Karlsson, M.E. Co-infection with Babesia divergens and Anaplasma phagocytophilum in cattle (Bos taurus), Sweden. Ticks Tick Borne Dis. 2017, 8, 933–935. [Google Scholar] [CrossRef]
- Alvarez, J.A.; Rojas, C.; Figueroa, J.V. Diagnostic Tools for the Identification of Babesia sp. in Persistently Infected Cattle. Pathogens 2019, 8, 143. [Google Scholar] [CrossRef] [Green Version]
- Uilenberg, G. International Collaborative Research—Significance of Tick-Borne Hemoparasitic Diseases to World Animal Health. Vet. Parasitol. 1995, 57, 19–41. [Google Scholar] [CrossRef]
- Bursakov, S.A.; Kovalchuk, S.N. Co-infection with tick-borne disease agents in cattle in Russia. Ticks Tick Borne Dis. 2019, 10, 709–713. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubalek, Z.; Foldvari, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Spitalska, E.; et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: New hazards and relevance for public health. Front. Public Health 2014, 2, 251. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J.; Telford, S.R., III; Spielman, A.; Sikand, V.; Ryan, R.; Christianson, D.; Burke, G.; Brassard, P.; Pollack, R.; Peck, J.; et al. Concurrent Lyme Disease and Babesiosis: Evidence for Increased Severity and Duration of Illness. JAMA 1996, 275, 1657–1660. [Google Scholar] [CrossRef]
- Saetre, K.; Godhwani, N.; Maria, M.; Patel, D.; Wang, G.; Li, K.I.; Wormser, G.P.; Nolan, S.M. Congenital Babesiosis After Maternal Infection with Borrelia burgdorferi and Babesia microti. J. Pediatr. Infect. Dis. Soc. 2017, 7, e1–e5. [Google Scholar] [CrossRef] [Green Version]
- Nyarko, E.; Grab, D.J.; Dumler, J.S. Anaplasma phagocytophilum-infected neutrophils enhance transmigration of Borrelia burgdorferi across the human blood brain barrier in vitro. Int. J. Parasitol. 2006, 36, 601–605. [Google Scholar] [CrossRef]
- Horowitz, H.W.; Aguero-Rosenfeld, M.E.; Holmgren, D.; McKenna, D.; Schwartz, I.; Cox, M.E.; Wormser, G.P. Lyme disease and human granulocytic anaplasmosis coinfection: Impact of case definition on coinfection rates and illness severity. Clin. Infect. Dis. 2013, 56, 93–99. [Google Scholar] [CrossRef]
- Jahfari, S.; Hofhuis, A.; Fonville, M.; van der Giessen, J.; van Pelt, W.; Sprong, H. Molecular Detection of Tick-Borne Pathogens in Humans with Tick Bites and Erythema Migrans, in the Netherlands. PLoS Negl. Trop. Dis. 2016, 10, e0005042. [Google Scholar] [CrossRef] [Green Version]
- Franke, J.; Hildebrandt, A.; Dorn, W. Exploring gaps in our knowledge on Lyme borreliosis spirochaetes--updates on complex heterogeneity, ecology, and pathogenicity. Ticks Tick Borne Dis. 2013, 4, 11–25. [Google Scholar] [CrossRef]
- Steere, A.C.; Sikand, V.K.; Meurice, F.; Parenti, D.L.; Fikrig, E.; Schoen, R.T.; Nowakowski, J.; Schmid, C.H.; Laukamp, S.; Buscarino, C.; et al. Vaccination against Lyme Disease with Recombinant Borrelia burgdorferi Outer-Surface Lipoprotein A with Adjuvant. New Engl. J. Med. 1998, 339, 209–215. [Google Scholar] [CrossRef]
- Sigal, L.H.; Zahradnik, J.M.; Lavin, P.; Patella, S.J.; Bryant, G.; Haselby, R.; Hilton, E.; Kunkel, M.; Adler-Klein, D.; Doherty, T.; et al. A vaccine consisting of recombinant Borrelia burgdorferi outer-surface protein A to prevent Lyme disease. Recombinant Outer-Surface Protein A Lyme Disease Vaccine Study Consortium. N. Engl. J. Med. 1998, 339, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Gross, D.M.; Forsthuber, T.; Tary-Lehmann, M.; Etling, C.; Ito, K.; Nagy, Z.A.; Field, J.A.; Steere, A.C.; Huber, B.T. Identification of LFA-1 as a candidate autoantigen in treatment-resistant Lyme arthritis. Science 1998, 281, 703–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steere, A.C.; Drouin, E.E.; Glickstein, L.J. Relationship between immunity to Borrelia burgdorferi outer-surface protein A (OspA) and Lyme arthritis. Clin. Infect. Dis. 2011, 52, s259–s265. [Google Scholar] [CrossRef] [PubMed]
- Comstedt, P.; Schüler, W.; Meinke, A.; Lundberg, U. The novel Lyme borreliosis vaccine VLA15 shows broad protection against Borrelia species expressing six different OspA serotypes. PLoS ONE 2017, 12, e0184357. [Google Scholar] [CrossRef] [PubMed]
- Wressnigg, N.; Barrett, P.N.; Pöllabauer, E.-M.; O’Rourke, M.; Portsmouth, D.; Schwendinger, M.G.; Crowe, B.A.; Livey, I.; Dvorak, T.; Schmitt, B.; et al. A Novel Multivalent OspA Vaccine against Lyme Borreliosis Is Safe and Immunogenic in an Adult Population Previously Infected with Borrelia burgdorferi Sensu Lato. Clin. Vaccine Immunol. 2014, 21, 1490–1499. [Google Scholar] [CrossRef]
- Amicizia, D.; Domnich, A.; Panatto, D.; Lai, P.L.; Cristina, M.L.; Avio, U.; Gasparini, R. Epidemiology of tick-borne encephalitis (TBE) in Europe and its prevention by available vaccines. Hum. Vaccines Immunother. 2013, 9, 1163–1171. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Tick-Borne Encephalitis (TBE) Vaccine; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2021. [Google Scholar]
- U.S. Food and Drug Administration Website. TICOVAC (Tick-Borne Encephalitis Vaccine) [Package Insert]. Pfizer Ireland Pharmaceuticals. Available online: https://www.fda.gov/media/151502/download (accessed on 13 May 2022).
- Walker, D.H. The realities of biodefense vaccines against Rickettsia. Vaccine 2009, 27, D52–D55. [Google Scholar] [CrossRef] [Green Version]
- Osterloh, A. Vaccine Design and Vaccination Strategies against Rickettsiae. Vaccines 2021, 9, 896. [Google Scholar] [CrossRef]
- Medicine, I.o. Critical Needs and Gaps in Understanding Prevention, Amelioration, and Resolution of Lyme and Other Tick-Borne Diseases: The Short-Term and Long-Term Outcomes: Workshop Report; The National Academies Press: Washington, DC, USA, 2011; p. 560. [Google Scholar]
- Rochlin, I.; Ninivaggi, D.V.; Benach, J.L. Malaria and Lyme disease—The largest vector-borne US epidemics in the last 100 years: Success and failure of public health. BMC Public Heal. 2019, 19, 804. [Google Scholar] [CrossRef]
- Maes, E.; Lecomte, P.; Ray, N. A cost-of-illness study of Lyme disease in the United States. Clin. Ther. 1998, 20, 993–1008. [Google Scholar] [CrossRef]
- Zhang, X.; Meltzer, M.I.; Peña, C.A.; Hopkins, A.B.; Wroth, L.; Fix, A.D. Economic impact of Lyme disease. Emerg. Infect. Dis. 2006, 12, 653–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinckley, A.F.; Connally, N.P.; Meek, J.I.; Johnson, B.J.; Kemperman, M.M.; Feldman, K.A.; White, J.L.; Mead, P.S. Lyme disease testing by large commercial laboratories in the United States. Clin. Infect. Dis. 2014, 59, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Beldner, S.J.; Jadonath, R.L.; Altman, E.J. A Safe and Cost-Effective Approach to Treating Lyme Cardiac Disease in an Era of Health Care Reform. Pacing Clin. Electrophysiol. 2011, 34, 666–669. [Google Scholar] [CrossRef] [PubMed]
- Adrion, E.R.; Aucott, J.; Lemke, K.W.; Weiner, J.P. Health care costs, utilization and patterns of care following Lyme disease. PLoS ONE 2015, 10, e0116767. [Google Scholar] [CrossRef]
- Henningsson, A.J.; Malmvall, B.E.; Ernerudh, J.; Matussek, A.; Forsberg, P. Neuroborreliosis—An epidemiological, clinical and healthcare cost study from an endemic area in the south-east of Sweden. Clin. Microbiol. Infect. 2010, 16, 1245–1251. [Google Scholar] [CrossRef] [Green Version]
- Joss, A.W.; Davidson, M.M.; Ho-Yen, D.O.; Ludbrook, A. Lyme disease—What is the cost for Scotland? Public Heal. 2003, 117, 264–273. [Google Scholar] [CrossRef]
- Lohr, B.; Müller, I.; Mai, M.; Norris, D.E.; Schöffski, O.; Hunfeld, K.P. Epidemiology and cost of hospital care for Lyme borreliosis in Germany: Lessons from a health care utilization database analysis. Ticks Tick Borne Dis. 2015, 6, 56–62. [Google Scholar] [CrossRef]
- Van den Wijngaard, C.C.; Hofhuis, A.; Wong, A.; Harms, M.G.; de Wit, G.A.; Lugnér, A.K.; Suijkerbuijk, A.W.M.; Mangen, M.J.; van Pelt, W. The cost of Lyme borreliosis. Eur. J. Public Heal. 2017, 27, 538–547. [Google Scholar] [CrossRef]
- Müller, I.; Freitag, M.H.; Poggensee, G.; Scharnetzky, E.; Straube, E.; Schoerner, C.; Hlobil, H.; Hagedorn, H.J.; Stanek, G.; Schubert-Unkmeir, A.; et al. Evaluating frequency, diagnostic quality, and cost of Lyme borreliosis testing in Germany: A retrospective model analysis. Clin. Dev. Immunol. 2012, 2012, 595427. [Google Scholar] [CrossRef] [Green Version]
- Connally, N.P.; Hinckley, A.F.; Feldman, K.A.; Kemperman, M.; Neitzel, D.; Wee, S.-B.; White, J.L.; Mead, P.S.; Meek, J.I. Testing practices and volume of non-Lyme tickborne diseases in the United States. Ticks Tick Borne Dis. 2016, 7, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Bish, E.K.; Moritz, E.D.; El-Amine, H.; Bish, D.R.; Stramer, S.L. Cost-effectiveness of Babesia microti antibody and nucleic acid blood donation screening using results from prospective investigational studies. Transfusion 2015, 55, 2256–2271. [Google Scholar] [CrossRef] [PubMed]
- Guerrero Espejo, A.; Muñoz Parada, C.; Tomás Dols, S. Incidence of human babesiosis in Spain obtained from the diagnoses at hospital discharge. Med. Clin. 2017, 149, 84–85. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Pena, A.; Salman, M. Current Limitations in the Control and Spread of Ticks that Affect Livestock: A Review. Agriculture 2013, 3, 221–235. [Google Scholar] [CrossRef] [Green Version]
- Kivaria, F.M. Estimated direct economic costs associated with tick-borne diseases on cattle in Tanzania. Trop. Anim. Heal. Prod. 2006, 38, 291–299. [Google Scholar] [CrossRef]
- Minjauw, B.; McLeod, A. Tick-borne diseases and poverty. The impact of ticks and tickborne diseases on the livelihood of small-scale and marginal livestock owners in India and eastern and southern Africa. Research report, DFID Animal Health Programme. Centre for Tropical Veterinary Medicine, University of Edinburgh, UK, 2003, 124.
- Islam, M.K.; Jabbar, A.; Campbell, B.E.; Cantacessi, C.; Gasser, R.B. Bovine theileriosis—An emerging problem in south-eastern Australia? Infect. Genet. Evol. 2011, 11, 2095–2097. [Google Scholar] [CrossRef] [PubMed]
- Perera, P.K.; Gasser, R.B.; Firestone, S.M.; Anderson, G.A.; Malmo, J.; Davis, G.; Beggs, D.S.; Jabbar, A. Oriental theileriosis in dairy cows causes a significant milk production loss. Parasites Vectors 2014, 7, 73. [Google Scholar] [CrossRef] [Green Version]
- Dinkel, K.D.; Herndon, D.R.; Noh, S.M.; Lahmers, K.K.; Todd, S.M.; Ueti, M.W.; Scoles, G.A.; Mason, K.L.; Fry, L.M. A US isolate of Theileria orientalis, Ikeda genotype, is transmitted to cattle by the invasive Asian longhorned tick, Haemaphysalis longicornis. Parasites Vectors 2021, 14, 157. [Google Scholar] [CrossRef]
- DeCastro, J.J. Sustainable tick and tickborne disease control in Livestock improvement in developing countries. Vet. Parasitol. 1997, 71, 77–97. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Host | Region | Host Species | Co-Infections | Prevalence (%) | Ref. |
|---|---|---|---|---|---|
| Small rodents | US | P. leucopus; M. pennsylvanicus | Bm + Bb | 35.7 | [15] |
| P. leucopus | 9.4 | [58] | |||
| 12 | [59] | ||||
| Bm + Ap | <1 | [59] | |||
| 1 | [58] | ||||
| Bb + Ap | 22.5 | [58] | |||
| 1 | [59] | ||||
| Bb + Ech | 2 | [59] | |||
| Ap + Ech | <1 | [59] | |||
| Bm + Bb + Ap | 12.8 | [58] | |||
| 6 | [59] | ||||
| Bm + Bb + Ech | 2 | [59] | |||
| Bb + Ap + Ech | <1 | [59] | |||
| Bm + Bb + Ap + Ech | 2 | [59] | |||
| Europe | Myodes glareolus | Ba + CNm | 42 | [60] | |
| Apodemus flavicollis | Bb + Bmiy | 9.3 | [61] | ||
| Apodemus spp. | Rs + Bl/Bv | 2 | [62] | ||
| Meso mammals | Europe | Canis lupus (grey woof) | Ap + Hc | <1 | [63] |
| Vulpes vulpes (red fox) | Babesia spp. + Hc | 17.6 | [64] | ||
| Ungulates (Large mammals) | Mexico | Bison bison (American Bison) | Bbov + Bb | 7.6 | [65] |
| Europe | Capreolus capreolus (roe deer) | Babesia spp. + Ap | 88.4 | [66] | |
| 79.9 | [67] | ||||
| 18.92 | [68] | ||||
| Cervus elaphus (red deer) | 1.9 | [69] | |||
| 62.2 | [67] | ||||
| Vector | Vector species | ||||
| Questing ticks | US | I. scapularis | Bm + Bb | 6.68 | [70] |
| 4.2 | [71] | ||||
| Bb + Ap | 2.47 | [70] | |||
| 1.8 | [71] | ||||
| Bm + Ap | <1 | [70] | |||
| Bm + Bmiy | <1 | [71] | |||
| Bb + Bmiy | 1.5 | [71] | |||
| Bm + Bb + Ap | <1 | [71] | |||
| Bm + Bb + Bmiy | <1 | [71] | |||
| Canada | I. scapularis | Babesia spp. + Bb | 1.15 | [72] | |
| Europe | I. ricinus | Babesia spp. + Ap | <1 | [66] | |
| Babesia spp. + Rickettsia spp. | 1.5 | [66] | |||
| Ap + Rickettsia spp. | 1.5 | [66] | |||
| Engorged ticks (Collected from birds, rodents, meso mammals and ungulates) | US | I. scapularis | Bm + Bb | <1, 12.69 | [70] |
| <1 | [71] | ||||
| Bm + Ap | <1 | [70] | |||
| 2.6 | [71] | ||||
| Bb + Ap | 9.9 | [71] | |||
| <1, 3.5 | [70] | ||||
| Bb + Bmiy | <1 | [71] | |||
| Bmiy + Ap | 1.4 | [71] | |||
| Bm + Bb + Ap | 1.7 | [71] | |||
| Bb + Ap + Bmiy | <1 | [71] | |||
| Bm + Bb + Bmiy + Ap | <1 | [71] | |||
| Canada | Ixodes spp. and Haemaphysalis leporispalustris | Babesia spp. + Bb | 2.2 | [72] | |
| Europe | I. ricinus | Babesia spp. + Ap | 6.3 | [66] | |
| Babesia spp. + Rickettsia spp. | 1.5 | [66] | |||
| Ap + Rickettsia spp. | 15.1 | [66] | |||
| Ap + Theileria spp. | <1 | [28] | |||
| Ap + Hepatozoon spp. | <1 | [28] | |||
| Borrelia spp. + Ap + Rh + Theileria spp. | <1 | [28] | |||
| Bbav/Bg + Ap | <1 | [28] | |||
| Ba/Bv + Rickettsia spp. | <1 | [28] | |||
| Bmiy + Ap | <1 | [28] | |||
| Ixodes spp. | Ap + Rh | 9.9 | [28] | ||
| Bg + Ap | 3.1 | [28] | |||
| Borrelia spp. + Ap + Rickettsia spp. | 18.9 | [28] | |||
| Bbav/Bg + Ap + Nm | <1 | [28] | |||
| Ixodes spp. and Haemaphysalis | Ba + Ap | 9 | [28] | ||
| Punctata | Ap + Rm | 3.1 | [28] | ||
| Hyalomma marginatum | Prp + Ehrlichia spp. | 12.19 | [73] | ||
| marginatum, Rhipicephalus bursa | Ehrlichia spp. + Ap | 9.75, 12.76 | [73] | ||
| and Dermacentor marginatus | Ehrlichia spp. + Am | 4.88 | [73] | ||
| Ehrlichia spp. + Ap + Am | 4.88, 6.38 | [73] | |||
| Prp + Ehrlichia spp. + Ap + Am | 7.32 | [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocha, S.C.; Velásquez, C.V.; Aquib, A.; Al-Nazal, A.; Parveen, N. Transmission Cycle of Tick-Borne Infections and Co-Infections, Animal Models and Diseases. Pathogens 2022, 11, 1309. https://doi.org/10.3390/pathogens11111309
Rocha SC, Velásquez CV, Aquib A, Al-Nazal A, Parveen N. Transmission Cycle of Tick-Borne Infections and Co-Infections, Animal Models and Diseases. Pathogens. 2022; 11(11):1309. https://doi.org/10.3390/pathogens11111309
Chicago/Turabian StyleRocha, Sandra C., Clara Vásquez Velásquez, Ahmed Aquib, Aya Al-Nazal, and Nikhat Parveen. 2022. "Transmission Cycle of Tick-Borne Infections and Co-Infections, Animal Models and Diseases" Pathogens 11, no. 11: 1309. https://doi.org/10.3390/pathogens11111309