
Repeated Tick Infestations Impair Borrelia burgdorferi Transmission in a Non-Human Primate Model of Tick Feeding
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement for Animal Use
2.2. Generation of Ixodes Scapularis Pathogen-Free and B. burgdorferi-Infected Nymphs
2.3. Tick Infestation of Rhesus Monkeys
2.4. Tick Transmission of B. burgdorferi
2.5. Skin Biopsy, Histology, Necropsy, and B. burgdorferi Evaluation
2.6. Skin Biopsy, Necropsy, and B. burgdorferi Evaluation
2.7. Statistical Analysis
3. Results
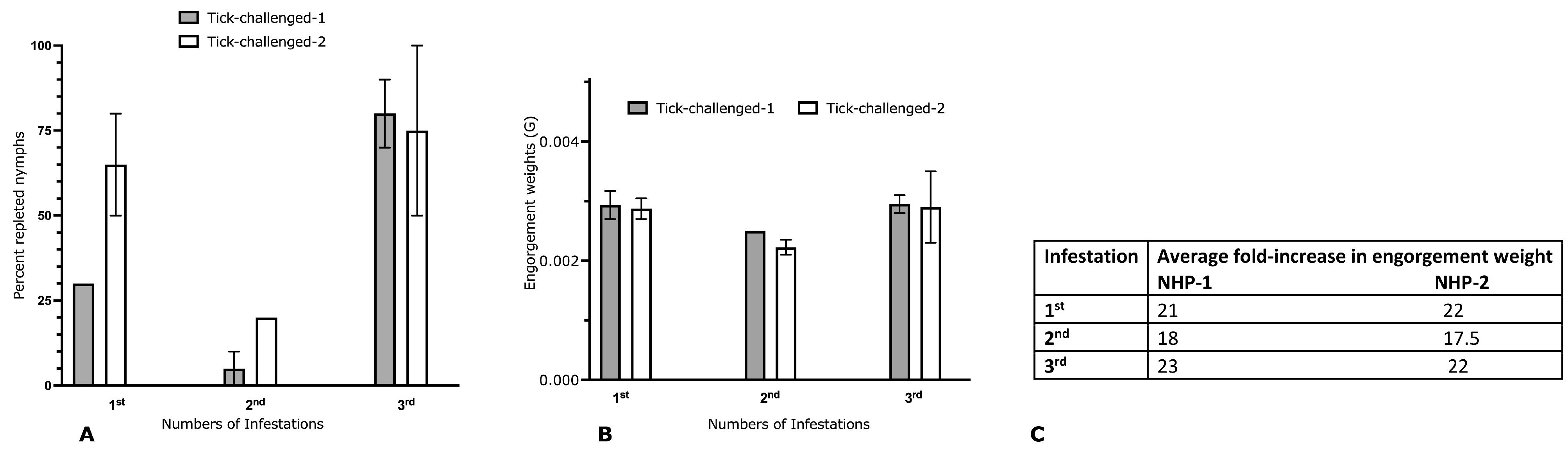
3.1. Repeated Infestation of Non-Human Primates with I. scapularis Does Not Elicit the Hallmarks of Acquired Tick Resistance
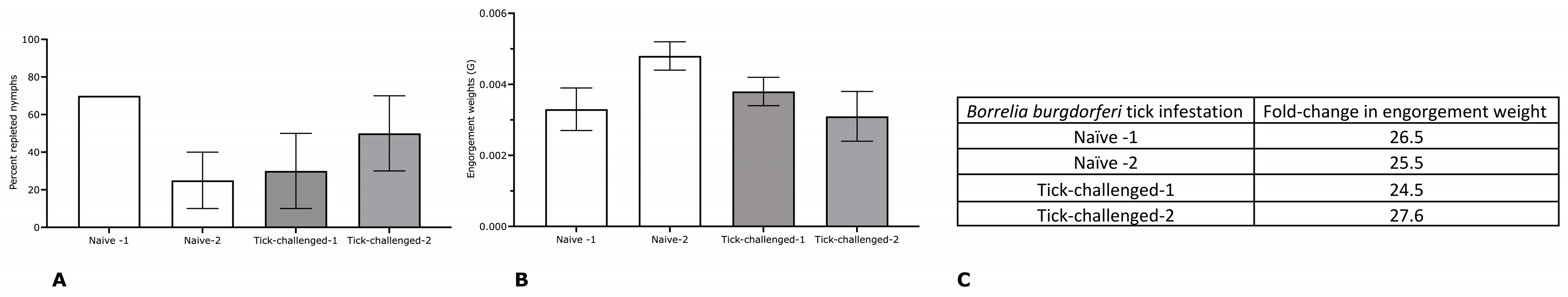
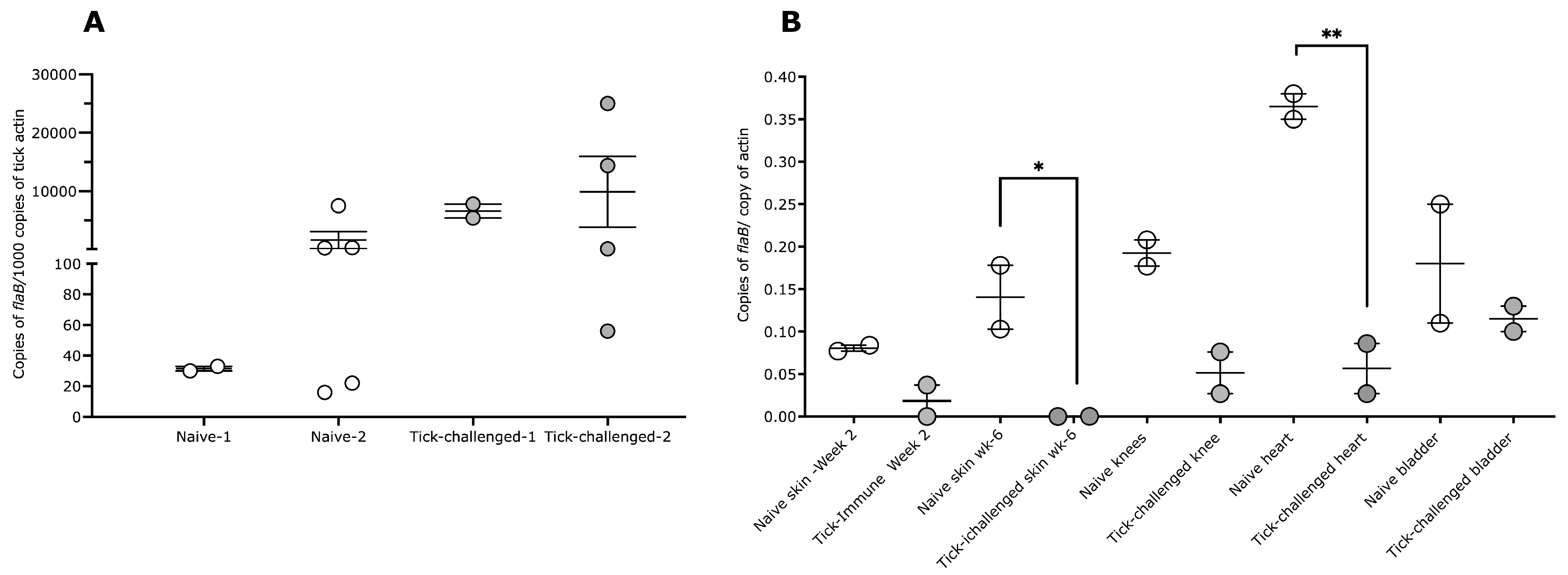
3.2. Tick Transmission of B. burgdorferi Is Impaired on Three-Time Tick-Infested Non-Human Primates
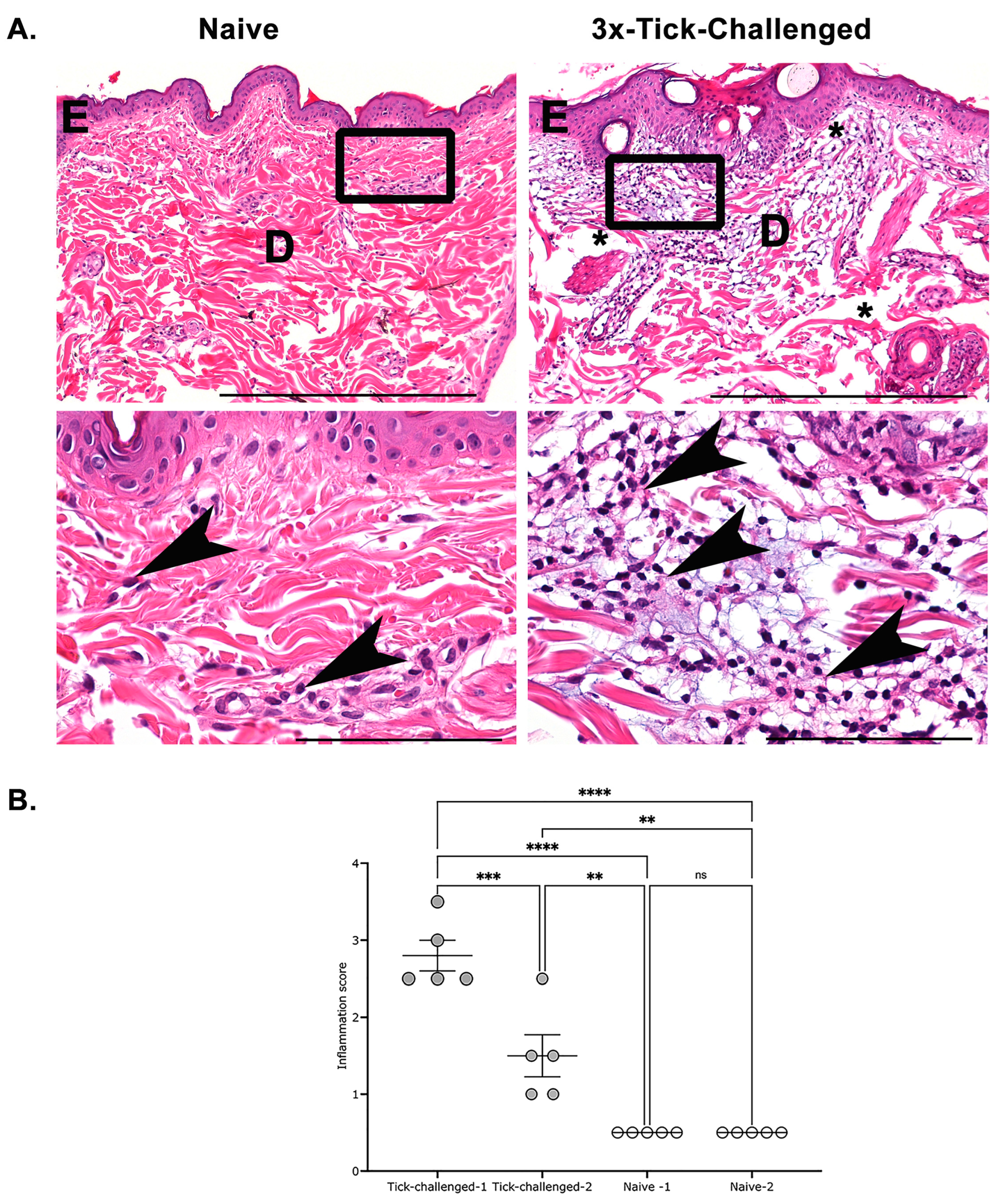
3.3. Histological Assessment of Skin Biopsies Demonstrates Increased Inflammation at the Tick Bite Site upon Repeated Tick Infestations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barbour, A.G.; Fish, D. The biological and social phenomenon of Lyme disease. Science 1993, 260, 1610–1616. [Google Scholar] [PubMed] [Green Version]
- Diuk-Wasser, M.A.; Vannier, E.; Krause, P.J. Coinfection by Ixodes Tick-Borne Pathogens: Ecological, Epidemiological, and Clinical Consequences. Trends Parasitol. 2016, 32, 30–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokarz, R.; Jain, K.; Bennett, A.; Briese, T.; Lipkin, W.I. Assessment of polymicrobial infections in ticks in New York state. Vector Borne Zoonotic Dis. 2010, 10, 217–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes-Solecki, M.; Arnaboldi, P.M.; Backenson, P.B.; Benach, J.L.; Cooper, C.L.; Dattwyler, R.J.; Doherty, T. Protective Immunity and New Vaccines for Lyme Disease. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2020, 70, 1768–1773. [Google Scholar]
- Sigal, L.H.; Zahradnik, J.M.; Lavin, P.; Patella, S.J.; Bryant, G.; Haselby, R.; Hilton, E.; Kunkel, M.; Adler-Klein, D. A vaccine consisting of recombinant Borrelia burgdorferi outer-surface protein A to prevent Lyme disease. Recombinant Outer-Surface Protein A Lyme Disease Vaccine Study Consortium. N. Engl. J. Med. 1998, 339, 216–222. [Google Scholar] [CrossRef]
- Steere, A.C.; Sikand, V.K.; Meurice, F.; Parenti, D.L.; Fikrig, E.; Schoen, R.T.; Nowakowski, J.; Schmid, C.H.; Laukamp, S.; Buscarino, C.; et al. Vaccination against Lyme disease with recombinant Borrelia burgdorferi outer-surface lipoprotein A with adjuvant. Lyme Disease Vaccine Study Group. N. Engl. J. Med. 1998, 339, 209–215. [Google Scholar] [CrossRef]
- Poland, G.A. Vaccines against Lyme disease: What happened and what lessons can we learn? Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2011, 52 (Suppl. 3), s253–s258. [Google Scholar] [CrossRef]
- Lathrop, S.L.; Ball, R.; Haber, P.; Mootrey, G.T.; Braun, M.; Shadomy, S.V.; Ellenberg, S.S.; Chen, R.T.; Hayes, E.B. Adverse event reports following vaccination for Lyme disease: December 1998-July 2000. Vaccine 2002, 20, 1603–1608. [Google Scholar] [CrossRef]
- Embers, M.E.; Narasimhan, S. Vaccination against Lyme disease: Past, present, and future. Front. Cell. Infect. Microbiol. 2013, 3, 6. [Google Scholar] [CrossRef] [Green Version]
- Wormser, G.P. A brief history of OspA vaccines including their impact on diagnostic testing for Lyme disease. Diagn. Microbiol. Infect. Dis. 2022, 102, 115572. [Google Scholar] [CrossRef]
- Dattwyler, R.J.; Gomes-Solecki, M. The year that shaped the outcome of the OspA vaccine for human Lyme disease. NPJ Vaccines 2022, 7, 10. [Google Scholar] [PubMed]
- Nigrovic, L.E.; Thompson, K.M. The Lyme vaccine: A cautionary tale. Epidemiol. Infect. 2007, 135, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nayak, A.; Schüler, W.; Seidel, S.; Gomez, I.; Meinke, A.; Comstedt, P.; Lundberg, U. Broadly Protective Multivalent OspA Vaccine against Lyme Borreliosis, Developed Based on Surface Shaping of the C-Terminal Fragment. Infect. Immun. 2020, 88, 4. [Google Scholar] [CrossRef]
- Izac, J.R.; O’Bier, N.S.; Oliver, L.D.; Camire, A.C.; Earnhart, C.G.; Rhodes, D.V.L.; Young, B.F.; Parnham, S.R.; Davies, C.; Marconi, R.T. Development and optimization of OspC chimeritope vaccinogens for Lyme disease. Vaccine 2020, 38, 1915–1924. [Google Scholar] [CrossRef] [PubMed]
- Nuttall, P.A. Wonders of tick saliva. Ticks Tick-Borne Dis. 2019, 10, 470–481. [Google Scholar] [PubMed] [Green Version]
- Narasimhan, S.; Kurokawa, C.; DeBlasio, M.; Matias, J.; Sajid, A.; Pal, U.; Lynn, G.; Fikrig, E. Acquired tick resistance: The trail is hot. Parasite Immunol. 2020, 43, e12808. [Google Scholar]
- Ndawula, C., Jr.; Tabor, A.E. Cocktail Anti-Tick Vaccines: The Unforeseen Constraints and Approaches toward Enhanced Efficacies. Vaccines 2020, 8, 457. [Google Scholar] [CrossRef]
- Ribeiro, J.M.; Francischetti, I.M. Role of arthropod saliva in blood feeding: Sialome and post-sialome perspectives. Annu. Rev. Entomol. 2003, 48, 73–88. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, J.M.; Alarcon-Chaidez, F.; Francischetti, I.M.B.; Mans, B.J.; Mather, T.N.; Valenzuela, J.G.; Wikel, S.K. An annotated catalog of salivary gland transcripts from Ixodes scapularis ticks. Insect Biochem. Mol. Biol. 2006, 36, 111–129. [Google Scholar]
- Perner, J.; Kropackova, S.; Kopacek, P.; Ribeiro, J.M.C. Sialome diversity of ticks revealed by RNAseq of single tick salivary glands. PLoS Negl. Trop. Dis. 2018, 12, e0006410. [Google Scholar] [CrossRef] [Green Version]
- Trager, W. Accquired immunity to ticks. J. Parasitol. 1939, 25, 57–81. [Google Scholar] [CrossRef]
- Nazario, S.; DePonte, K.; De Silva, A.M.; Kantor, F.S.; Fikrig, E.; Fish, D.; Anderson, J.F.; Das, S.; Marcantonio, N. Prevention of Borrelia burgdorferi transmission in guinea pigs by tick immunity. Am. J. Trop. Med. Hyg. 1998, 58, 780–785. [Google Scholar] [CrossRef]
- Narasimhan, S.; Kurokawa, C.; Diktas, H.; Strank, N.O.; Černý, J.; Murfin, K.; Cao, Y.; Lynn, G.; Trentleman, J.; Wu, M.-J.; et al. Ixodes scapularis saliva components that elicit responses associated with acquired tick-resistance. Ticks Tick-Borne Dis. 2020, 11, 101369. [Google Scholar] [CrossRef] [PubMed]
- Sonnesyn, S.W.; Manivel, J.C.; Johnson, R.C.; Goodman, J.L. A guinea pig model for Lyme disease. Infect. Immun. 1993, 61, 4777–4784. [Google Scholar] [CrossRef] [Green Version]
- Burke, G.; Wikel, S.K.; Spielman, A.; Telford, S.R.; McKay, K.; Krause, P.J. Hypersensitivity to ticks and Lyme disease risk. Emerg. Infect. Dis. 2005, 11, 36–41. [Google Scholar] [PubMed] [Green Version]
- Pachner, A.R.; Gelderblom, H.; Cadavid, D. The rhesus model of Lyme neuroborreliosis. Immunol. Rev. 2001, 183, 186–204. [Google Scholar] [CrossRef]
- Philipp, M.T.; Aydintug, M.K.; Bohm, R.P.; Cogswell, F.B.; Dennis, V.; Lanners, H.N.; Lowrie, R.C.; Roberts, E.D.; Conway, M.D.; Karaçorlu, M.; et al. Early and early disseminated phases of Lyme disease in the rhesus monkey: A model for infection in humans. Infect. Immun. 1993, 61, 3047–3059. [Google Scholar] [CrossRef] [Green Version]
- Narasimhan, S.; Rajeevan, N.; Liu, L.; Zhao, Y.O.; Heisig, J.; Pan, J.; Eppler-Epstein, R.; DePonte, K.; Fish, D.; Fikrig, E. Gut microbiota of the tick vector Ixodes scapularis modulate colonization of the Lyme disease spirochete. Cell Host Microbe 2014, 15, 58–71. [Google Scholar] [CrossRef] [Green Version]
- Narasimhan, S.; Schuijt, T.J.; Abraham, N.M.; Rajeevan, N.; Coumou, J.; Graham, M.; Robson, A.; Wu, M.-J.; Daffre, S.; Hovius, J.W.; et al. Modulation of the tick gut milieu by a secreted tick protein favors Borrelia burgdorferi colonization. Nat. Commun. 2017, 8, 184. [Google Scholar] [CrossRef] [Green Version]
- Narasimhan, S.; DePonte, K.; Marcantonio, N.; Liang, X.; Royce, T.E.; Nelson, K.F.; Booth, C.J.; Koski, B.; Anderson, J.F.; Kantor, F.; et al. Immunity against Ixodes scapularis salivary proteins expressed within 24 hours of attachment thwarts tick feeding and impairs Borrelia transmission. PLoS ONE 2007, 2, e451. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, R.R.; Booth, C.J.; Wang, X.; Blaho, V.A.; Malawista, S.E.; Brown, C.R. Recruitment of macrophages and polymorphonuclear leukocytes in Lyme carditis. Infect. Immun. 2007, 75, 613–620. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.J.; Askenase, P.W. Immune rejection of ectoparasites (ticks) by T cell and IgG1 antibody recruitment of basophils and eosinophils. Fed. Proc. 1983, 42, 1744–1749. [Google Scholar]
- Brown, S.J.; Worms, M.J.; Askenase, P.W. Rhipicephalus appendiculatus: Larval feeding sites in guinea pigs actively sensitized and receiving immune serum. Exp. Parasitol. 1983, 55, 111–120. [Google Scholar] [CrossRef]
- Brown, S.J.; Askenase, P.W. Cutaneous basophil responses and immune resistance of guinea pigs to ticks: Passive transfer with peritoneal exudate cells or serum. J. Immunol. 1981, 127, 2163–2167. [Google Scholar] [CrossRef]
- Wikel, S.K.; Allen, J.R. Acquired resistance to ticks. I. Passive transfer of resistance. Immunology 1976, 30, 311–316. [Google Scholar]
- Das, S.; Banerjee, G.; DePonte, K.; Marcantonio, N.; Kantor, F.S.; Fikrig, E. Salp25D, an Ixodes scapularis antioxidant, is 1 of 14 immunodominant antigens in engorged tick salivary glands. J. Infect. Dis. 2001, 184, 1056–1064. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.R.; Kemp, D.H. Observations on the behaviour of Dermacentor andersoni larvae infesting normal and tick resistant guinea-pigs. Parasitology 1982, 84 Pt 2, 195–204. [Google Scholar] [CrossRef]
- Wada, T.; Ishiwata, K.; Koseki, H.; Ishikura, T.; Ugajin, T.; Ohnuma, N.; Obata-Ninomiya, K.; Ishikawa, R.; Yoshikawa, S.; Mukai, K.; et al. Selective ablation of basophils in mice reveals their nonredundant role in acquired immunity against ticks. J. Clin. Investig. 2010, 120, 2867–2875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, S.J.; Barker, R.W.; Askenase, P.W. Bovine resistance to Amblyomma americanum ticks: An acquired immune response characterized by cutaneous basophil infiltrates. Vet. Parasitol. 1984, 16, 147–165. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.J.; Askenase, P.W. Amblyomma americanum: Requirement for host Fc receptors in antibody-mediated acquired immune resistance to ticks. Exp. Parasitol. 1985, 59, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Ohta, T.; Yoshikawa, S.; Tabakawa, Y.; Yamaji, K.; Ishiwata, K.; Shitara, H.; Taya, C.; Oh-Hora, M.; Kawano, Y.; Miyake, K.; et al. Skin CD4(+) Memory T Cells Play an Essential Role in Acquired Anti-Tick Immunity through Interleukin-3-Mediated Basophil Recruitment to Tick-Feeding Sites. Front. Immunol. 2017, 8, 1348. [Google Scholar] [CrossRef] [Green Version]
- Tabakawa, Y.; Ohta, T.; Yoshikawa, S.; Robinson, E.J.; Yamaji, K.; Ishiwata, K.; Kawano, Y.; Miyake, K.; Yamanishi, Y.; Ohtsu, H.; et al. Histamine Released from Skin-Infiltrating Basophils but Not Mast Cells Is Crucial for Acquired Tick Resistance in Mice. Front. Immunol. 2018, 9, 1540. [Google Scholar] [CrossRef] [PubMed]
- .Kim, T.K.; Tirloni, L.; Pinto, A.F.; Moresco, J.; Yates, J.R., 3rd; da Silva Vaz, I., Jr. Ixodes scapularis Tick Saliva Proteins Sequentially Secreted Every 24 h during Blood Feeding. PLoS Negl. Trop. Dis. 2016, 10, e0004323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynn, G.E.; Černý, J.; Kurokawa, C.; Diktaş, H.; Matias, J.; Sajid, A.; Arora, G.; DePonte, K.; Narasimhan, S.; Fikrig, E. Immunization of guinea pigs with cement extract induces resistance against Ixodes scapularis ticks. Ticks Tick-Borne Dis. 2022, 13, 102017. [Google Scholar] [CrossRef] [PubMed]
- Kotál, J.; Langhansová, H.; Lieskovská, J.; Andersen, J.F.; Francischetti, I.M.; Chavakis, T.; Kopecký, J.; Pedra, J.H.; Kotsyfakis, M.; Chmelař, J. Modulation of host immunity by tick saliva. J. Proteom. 2015, 128, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Sajid, A.; Matias, J.; Arora, G.; Kurokawa, C.; DePonte, K.; Tang, X.; Lynn, G.; Wu, M.-J.; Pal, U.; Strank, N.O.; et al. mRNA vaccination induces tick resistance and prevents transmission of the Lyme disease agent. Sci. Transl. Med. 2021, 13, eabj9827. [Google Scholar] [CrossRef]
- Narasimhan, S.; Booth, C.J.; DePonte, K.; Wu, M.-J.; Liang, X.; Mohanty, S.; Kantor, F.; Fikrig, E. Host-specific expression of Ixodes scapularis salivary genes. Ticks Tick-Borne Dis. 2019, 10, 386–397. [Google Scholar] [CrossRef]
- Anderson, J.M.; Moore, I.N.; Nagata, B.M.; Ribeiro, J.M.C.; Valenzuela, J.G.; Sonenshine, D.E. Ticks, Ixodes scapularis, Feed Repeatedly on White-Footed Mice despite Strong Inflammatory Response: An Expanding Paradigm for Understanding Tick-Host Interactions. Front. Immunol. 2017, 8, 1784. [Google Scholar] [CrossRef] [Green Version]
- Kurokawa, C.; Narasimhan, S.; Vidyarthi, A.; Booth, C.J.; Mehta, S.; Meister, L.; Diktas, H.; Strank, N.; Lynn, G.E.; DePonte, K.; et al. Repeat tick exposure elicits distinct immune responses in guinea pigs and mice. Ticks Tick-Borne Dis. 2020, 11, 101529. [Google Scholar]
- Matsuda, H.; Ptak, W.; Askenase, P.W. Role of mast cells versus basophils in IgE-dependent local ear skin release of the serotonin required to initiate contact sensitivity in mice. Int. Arch. Allergy Immunol. 1995, 107, 364. [Google Scholar] [CrossRef]
- McLaren, D.J.; Worms, M.J.; Askenase, P.W. Cutaneous basophil associated resistance to ectoparasites (ticks). Electron microscopy of Rhipicephalus appendiculatus larval feeding sites in actively sensitised guinea pigs and recipients of immune serum. J. Pathol. 1983, 139, 291–308. [Google Scholar] [CrossRef] [PubMed]
- McLaren, D.J.; Worms, M.J.; Brown, S.J.; Askenase, P.W. Ornithodorus tartakovskyi: Quantitation and ultrastructure of cutaneous basophil responses in the guinea pig. Exp. Parasitol. 1983, 56, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Karasuyama, H.; Miyake, K.; Yoshikawa, S. Immunobiology of Acquired Resistance to Ticks. Front. Immunol. 2020, 11, 601504. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, S.; Sukumaran, B.; Bozdogan, U.; Thomas, V.; Liang, X.; DePonte, K. A tick antioxidant facilitates the Lyme disease agent’s successful migration from the mammalian host to the arthropod vector. Cell Host Microbe 2007, 2, 7–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Q.; Seemanapalli, S.V.; Reif, K.E.; Brown, C.R.; Liang, F.T. Increasing the recruitment of neutrophils to the site of infection dramatically attenuates Borrelia burgdorferi infectivity. J. Immunol. 2007, 178, 5109–5115. [Google Scholar] [CrossRef] [Green Version]
- Menten-Dedoyart, C.; Faccinetto, C.; Golovchenko, M.; Dupiereux, I.; Van Lerberghe, P.B.; Dubois, S.; Piesman, M. Neutrophil extracellular traps entrap and kill Borrelia burgdorferi sensu stricto spirochetes and are not affected by Ixodes ricinus tick saliva. J. Immunol. 2012, 189, 5393–5401. [Google Scholar] [CrossRef]
- Ribeiro, J.M.; Weis, J.J.; Telford, S.R. Saliva of the tick Ixodes dammini inhibits neutrophil function. Exp. Parasitol. 1990, 70, 382–388. [Google Scholar] [CrossRef]
- Wikel, S.K.; Ramachandra, R.N.; Bergman, D.K.; Burkot, T.R.; Piesman, J. Infestation with pathogen-free nymphs of the tick Ixodes scapularis induces host resistance to transmission of Borrelia burgdorferi by ticks. Infect. Immun. 1997, 65, 335–338. [Google Scholar] [CrossRef] [Green Version]
- Ramamoorthi, N.; Narasimhan, S.; Pal, U.; Bao, F.; Yang, X.F.; Fish, D.; Anguita, J.; Norgard, M.V.; Kantor, F.S.; Anderson, J.F.; et al. The Lyme disease agent exploits a tick protein to infect the mammalian host. Nature 2005, 436, 573–577. [Google Scholar] [CrossRef] [Green Version]
- Foley, J.E.; Lerche, N.W.; Dumler, J.S.; Madigan, J.E. A simian model of human granulocytic ehrlichiosis. Am. J. Trop. Med. Hyg. 1999, 60, 987–993. [Google Scholar] [CrossRef] [Green Version]
- Gumber, S.; Nascimento, F.S.; Rogers, K.A.; Bishop, H.S.; Rivera, H.N.; Xayavong, M.V.; Fikrig, E. Experimental transfusion-induced Babesia microti infection: Dynamics of parasitemia and immune responses in a rhesus macaque model. Transfusion 2016, 56 Pt 2, 1508–1519. [Google Scholar] [CrossRef] [Green Version]
- Ruebush, T.K., 2nd; Piesman, J.; Collins, W.E.; Spielman, A.; Warren, M. Tick transmission of Babesia microti to rhesus monkeys (Macaca mulatta). Am. J. Trop. Med. Hyg. 1981, 30, 555–559. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue | Naïve 1 | Naïve 2 | 3x-Tick Challenged 1 | 3x-Tick Challenged 2 |
|---|---|---|---|---|
| Skin-2 weeks post tick detachment | Positive | Positive | Negative | Negative |
| Skin-6 weeks post tick detachment | Positive | Positive | Negative | Negative |
| Heart | Positive | Positive | Positive | Positive |
| Joint/Knee | Positive | Positive | Positive | Positive |
| Bladder | Positive | Positive | Positive | Positive |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Narasimhan, S.; Booth, C.J.; Philipp, M.T.; Fikrig, E.; Embers, M.E. Repeated Tick Infestations Impair Borrelia burgdorferi Transmission in a Non-Human Primate Model of Tick Feeding. Pathogens 2023, 12, 132. https://doi.org/10.3390/pathogens12010132
Narasimhan S, Booth CJ, Philipp MT, Fikrig E, Embers ME. Repeated Tick Infestations Impair Borrelia burgdorferi Transmission in a Non-Human Primate Model of Tick Feeding. Pathogens. 2023; 12(1):132. https://doi.org/10.3390/pathogens12010132
Chicago/Turabian StyleNarasimhan, Sukanya, Carmen J. Booth, Mario T. Philipp, Erol Fikrig, and Monica E. Embers. 2023. "Repeated Tick Infestations Impair Borrelia burgdorferi Transmission in a Non-Human Primate Model of Tick Feeding" Pathogens 12, no. 1: 132. https://doi.org/10.3390/pathogens12010132