1. Introduction
Dimorphic individuals, in which the bodies consist of both female and male parts, were observed before the term
gynandromorphism was coined and described by Rudolfi for the moth species,
Gastropacha quercifolia, in 1825. Though this phenomenon is more commonly reported in insects than in other animal classes, the estimated occurrence of insect gynandromorphs is about 0.01 to 0.05%. Gynandromorphism has been observed in several insect orders including Lepidoptera, Hymenoptera, Coleoptera, Diptera, and Orthoptera, but its occurrence across orders is not even [
1]. Several thousands have been reported in Lepidoptera in comparison with 90 in Hymenoptera; however, this fact could be due to easily recognizable sexual dimorphic features in the former order [
2].
Lepidoptera (moths and butterflies) have a female heterogametic sex chromosome system, with most females having a WZ constitution while males are ZZ [
3]. The cause of bilateral gynandromorphism arises from an event in mitosis during a very early stage in the development of the organism. When the juvenile organism grows through a series of cell divisions, a genetic error in the chromosomes leads to one of the dividing cells splitting its sex chromosomes aberrantly, resulting in two dissimilar daughter cells, one having the chromosomes that cause female development and the other the chromosomes that cause male development.
While external dimorphism appears as morphological features like size, form, and color, internal dimorphism is reflected by gender-specific reproductive organs and brain anatomy. In our lab, the Chemosensory lab at the Norwegian University of Science and Technology (NTNU), an emerged moth of the species, Helicoverpa armigera, proved to be a bilateral gynandromorph, with a clear external segregation of the male and female appearances on the left and right side, respectively. In addition, internally observed bilateral brain structures gave us a unique opportunity to uncover putative sexual dimorphic traits of the moth brain, making a comparison of the gynandromorphs male and female hemispheres.
In the moth brain, the antennal lobe (AL) stands out as a prominent sexually dimorphic neuropil. Functionally, the moth AL, also called the primary olfactory center of the insect brain, plays a role in processing chemo-sensory information. The AL consists of spherical units, glomeruli, which are segregated into several groups based on their morphological and functional specificities [
4]. In
H. armigera, two recent studies performed a global-wide comparison between female and male ALs to identify sexually dimorphic and isomorphic glomeruli [
5,
6]. Three-dimensional reconstruction of the AL glomeruli revealed four distinct groups in both sexes. The most prominent sexual dimorphism in the moth species is demonstrated by the macroglomerular complex (MGC). In
H. armigera males, there are three glomeruli grouped into the MGC and processing input about female-produced components [
7]. In females, a small assembly of enlarged glomeruli, possibly being comparable with the MGC units in males, has been observed in several moth species:
Bombyx mori [
8],
Manduca sexta [
9], and
Heliothis virescens [
10]. In
H. armigera, five enlarged glomeruli were identified as female-specific units and belonging to the female complex (Fx; [
6]).
Generally, the largest group of glomeruli constitutes a category of so-called ordinary glomeruli (OG). In
H. armigera, the OGs demonstrate high homology, though the numbers of glomeruli in males and females are different; in the
H. armigera male there are 64–65 OGs and in female 66 [
6]. The main task of these glomeruli is to process plant odor information. Additionally, there are also some reports about their involvement in thermal and mechanical stimuli processing as well [
5,
11].
Another category of glomeruli is the so-called posterior complex (PCx), a cluster of dorso-posteriorly located units. The function of this group is not yet defined, though it was shown in several male heliothine species that one of the PCx glomeruli receives input from two co-located distinct types of pheromone sensory neurons [
7]. In addition, calcium imaging results also demonstrated pheromone-evoked responses in the PCx glomeruli (personal observations). The previous anatomical investigation of the AL in
H. armigera males and females showed sexual dimorphism in PCx, including a different number of glomeruli: ten in males and nine in females [
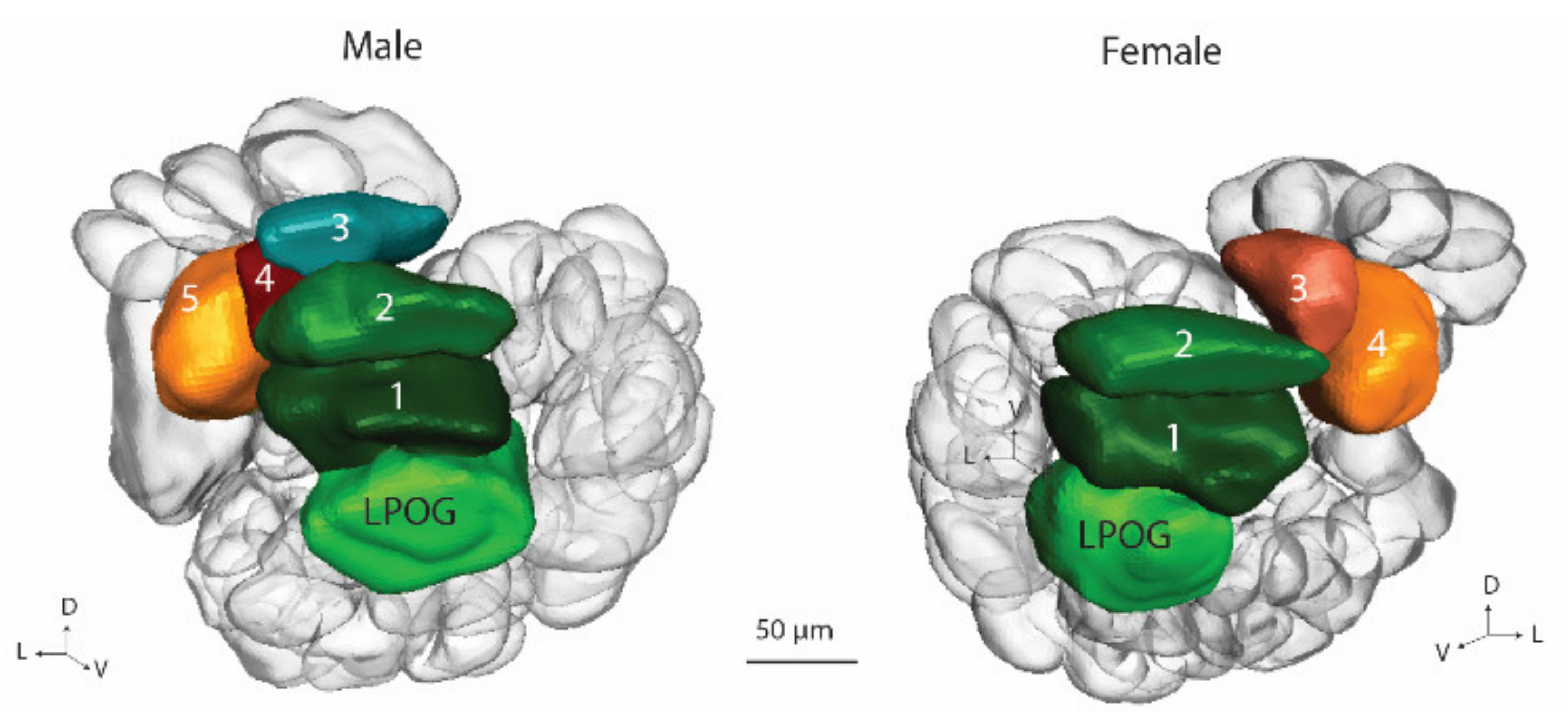
6]. The fourth glomerular category consists of a single large unit, the labial pit organ glomerulus (LPOG), receiving input from sensory neurons tuned to carbon dioxide [
12]. The former investigations of
H. armigera, showed that the LPOG volume is larger in females than in males [
6].
Whether other neuropils than the primary olfactory center may also have a sex difference is still an open question. Thus, in the present study, we examined the
H. armigera gynandromorph with respect to putative anatomical differences in the central brain areas of the two hemispheres. Brain regions including those for olfactory and visual processing were selected, as both olfactory and visual inputs are fundamental for moth navigation [
13,
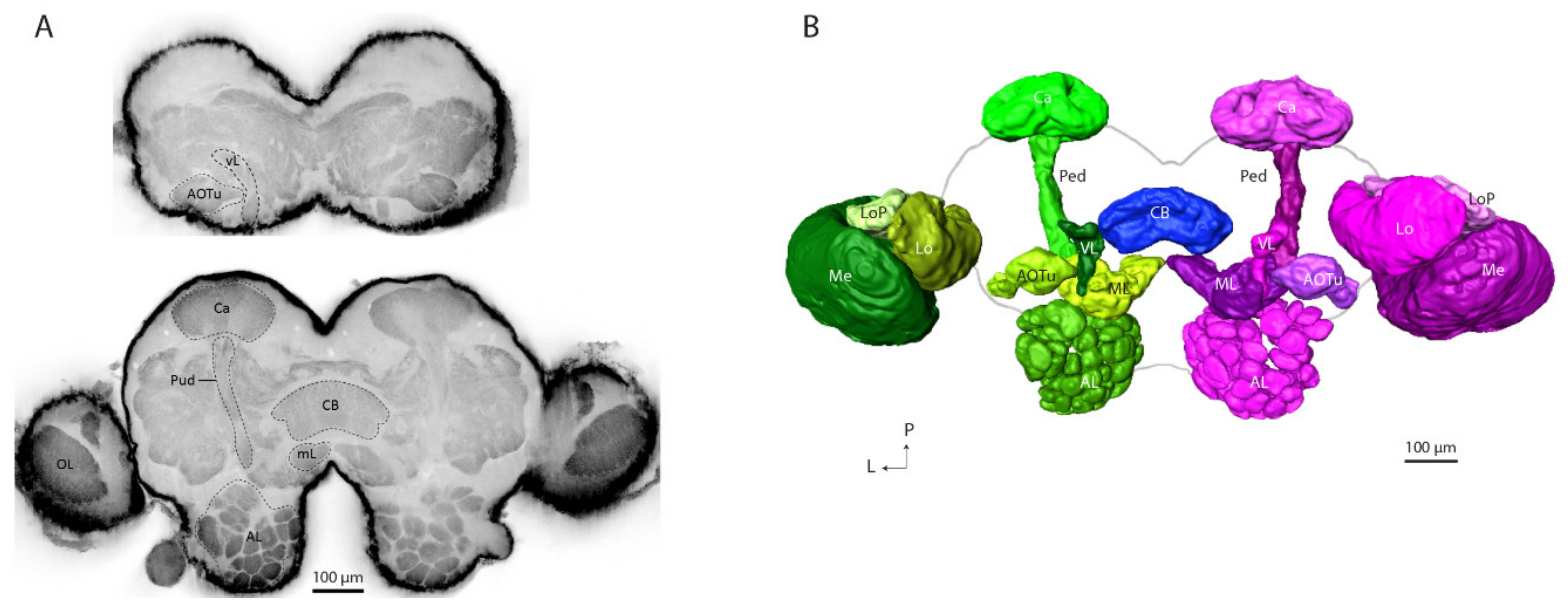
14]. Comparative quantitative analysis of the digital neuropil reconstructions was performed on the optical lobes (OL), the anterior optic tubercle (AOTu), and the mushroom bodies (MB) including the calyces, lobes, and the pedunculus. The results demonstrate that the sex differences are not only present in the AL, but also in some of the protocerebral neuropils in the higher brain centers.
2. Materials and Methods
2.1. Insects
Helicoverpa armigera pupae (Lepidoptera; Noctuidae, Heliothinae), delivered by Keyun Bio-pesticides (Henan, China), were reared in climate chambers (Refritherm 200 and 6E, Struers-Kebolab, Albertsund, Denmark, or Binder KBF 720, Tuttlingen, Germany) at 24 °C and 70% air humidity on a 14:10 h light/dark cycle. The individual used in this study was one day old. According to Norwegian law of animal welfare, there are no restrictions regarding the experimental use of Lepidoptera.
2.2. Immunostaining
The preparation procedure was described in detail elsewhere [
14,
15]. In short, the brain was dissected in Ringer’s solution (in mM: 150 NaCl, 3 CaCl
2, 3 KCl, 25 sucrose, and 10 N-tris (hydroxymethyl)-methyl-2-amino-ethanesulfonic acid, pH 6.9) and immediately transferred into a Zinc–Formaldehyde fixative [
16] at room temperature overnight. The brain was then washed in HEPES-buffered saline (HBS, 8 × 30 min), and subjected to a permeabilization step (60 min incubation with a fresh mixture of 20% DMSO and 80% methanol) before being washed 3 × 10 min in a Tris-HCL buffer (0.1 M, pH 7.4). After pre-incubation in 5% normal goat serum (NGS, Sigma–Aldrich, St. Louis, MO, USA) in 0.1 M phosphate-buffered saline (PBS, pH 7.2) containing 0.3% Triton X-100 (PBT), the brain was incubated for 6 days at 4 °C in the primary antibody, SYNORF1 (dilution 1:25 in PBT containing 1% NGS). Following rinsing in PBT 8 × 30 min, the brain was incubated for 5 days at 4 °C with Alexa Flour 647 conjugated goat-anti-mouse secondary antibody solution (Invitrogen, Eugene, OR; dilution 1:300 in PBT with 1% NGS). After washing 4 × 30 min in PBT and 2 × 30 min in PBS, the brain was dehydrated in an increasing ethanol series (50%, 70%, 90%, 95%, and 100% (2×), 10 min each). Then, the brain was transferred to a mixture of methyl salicylate and ethanol (1:1) for 10 min and after that cleared completely in methyl salicylate for at least 1 h. Finally, the brain was mounted in a Permount mounting medium (Electron Microscopy Sciences) between two coverslips, separated by spacers.
2.3. Confocal Image Acquisition
The immunostained brain was imaged dorso-frontally by using a confocal laser scanning microscope (LSM 800 Zeiss, Jena, Germany) equipped with a Plan-Neofluar 20×/0.5 objective. The sample was excited with a HeNe laser at 653 nm and the fluorescent emission passed through a 650 nm long-pass filter at a voxel size of 0.62 × 0.62 × 2 μm. The confocal images shown in this study were edited in ZEN 2.3 (blue edition, Carl Zeiss Microscopy GmbH, Jana, Germany).
2.4. Digital Reconstruction and Volumetric Analysis
The high-quality confocal stack allowed for making 3D visualizations based on the auto-fluorescence signals without an immunostaining application. A complete image of the raw confocal stack of the gynandromorph moth was used for making digital reconstructions of the relevant brain structures by means of the AMIRA software (AMIRA 6.0, Visage Imaging, Fürth, Germany). The brain areas were manually demarcated by using a segmentation editor for all three spatial planes with a consequent wrap-tool to obtain full neuropil volumes. Surface models of the segmented areas were constructed, and their volumes were measured by the material statistics function in the segmentation editor.
2.5. Glomerular Naming
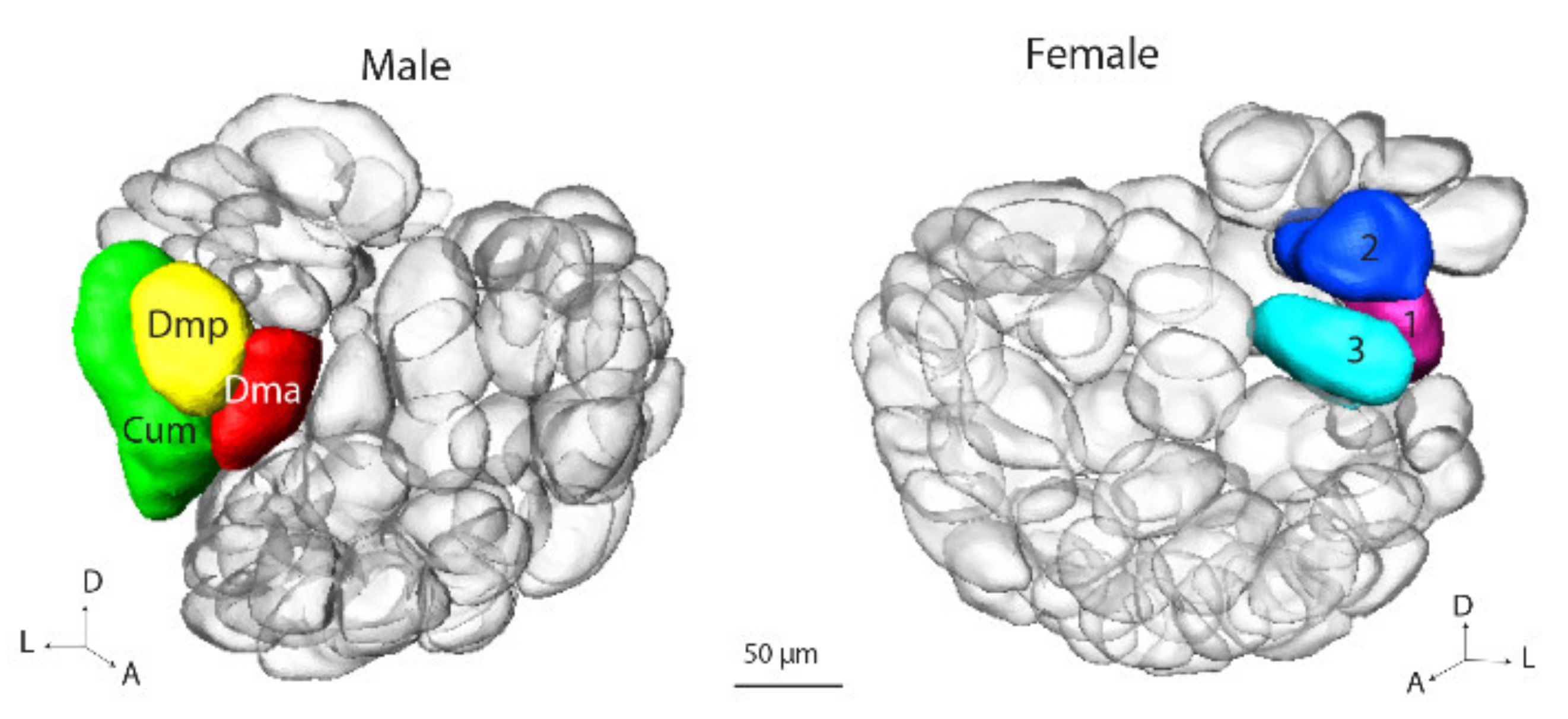
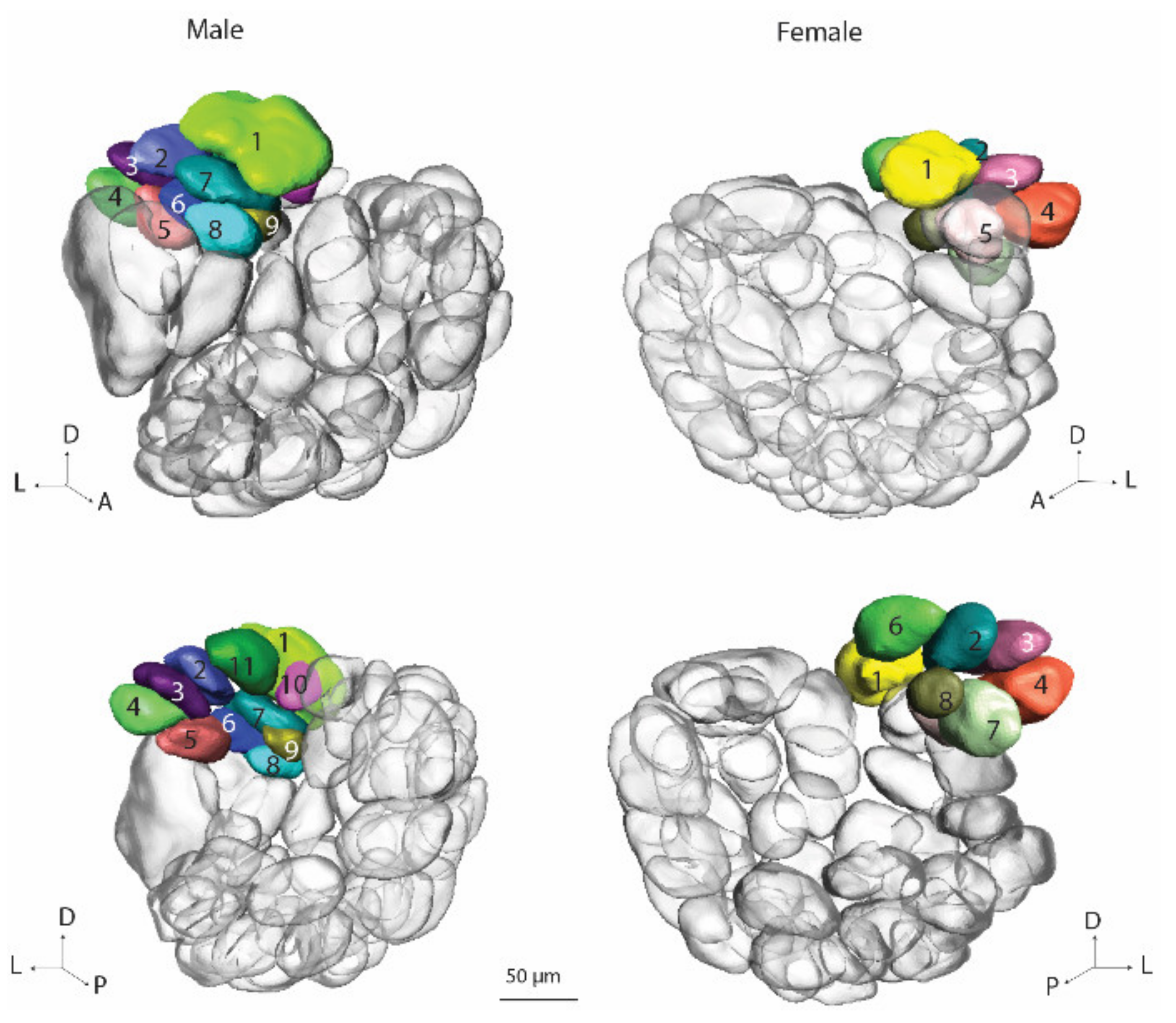
In correspondence with the previous classification of antennal-lobe glomeruli in the male and female heliothine moth, we grouped the units in four assemblies: (1) the male/female specific complex, (2) the ordinary glomeruli (OG), (3) the posterior complex (PCx), and (4) the labial pit organ glomerulus (LPOG) [
5,
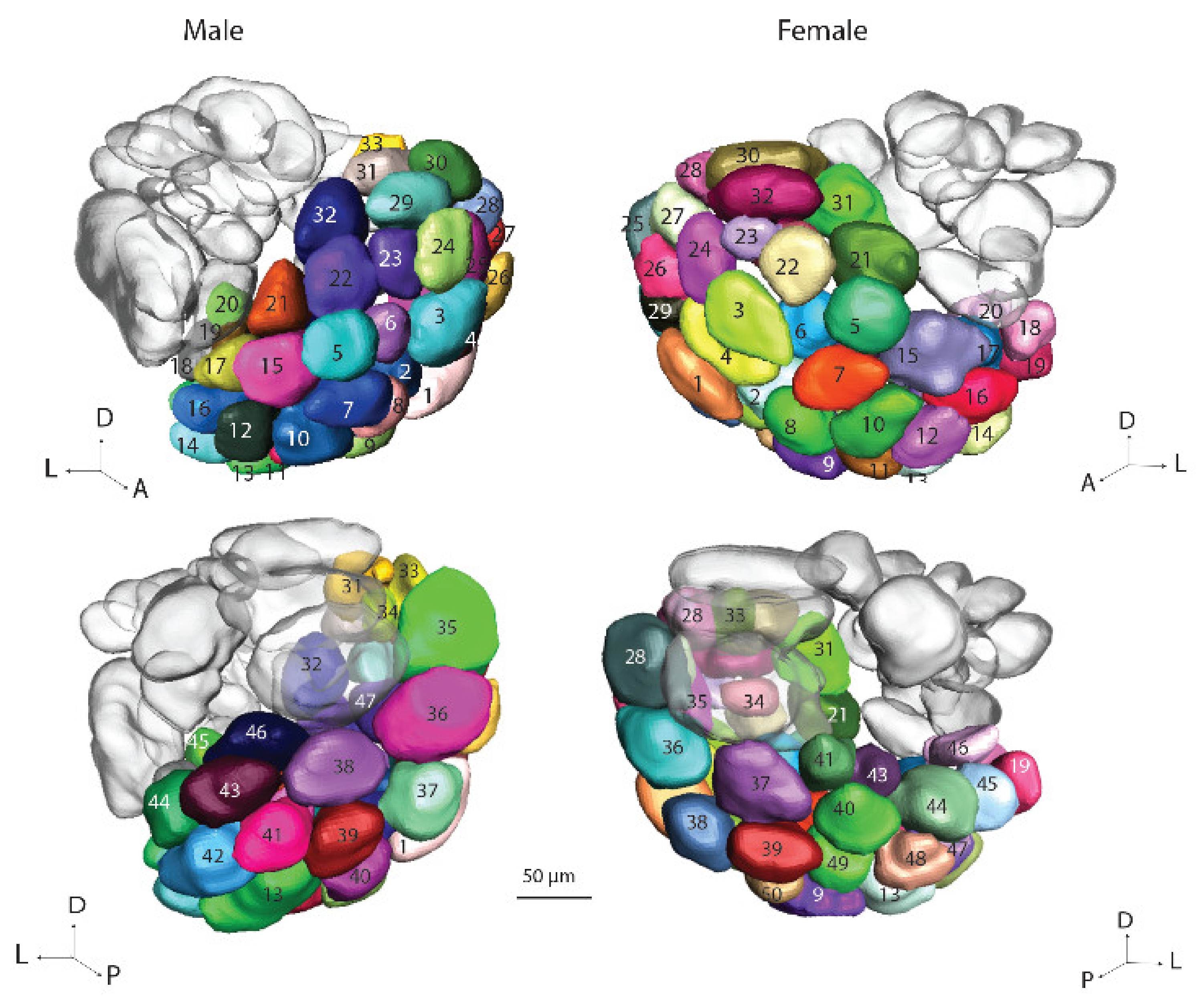
6]. For the male specific MGC, the previously established names, Cu, Dmp and Dma were used. For the female counterpart, namely, the female complex, Fx, the three relevant glomeruli were named, Fx1–Fx3. Since the OGs in the male and female hemispheres could not be identified with respect to homologies, they were numbered without any correspondence. The OGs in the male part were named O
m1–O
m66 and in the female part O
f1–O
f66. For the PCx units, the names P
m1–P
m9 were used for the glomeruli in the male hemisphere and P
f1–P
f9 for the glomeruli in the female hemisphere. The LPOG unit was named L
m and L
f for male and female halves, respectively. In addition to the four above-mentioned glomerular assemblies, we created a new one, the ventroposterior complex (VPx), which was selected out of the OGs and named V
m1–V
m5 and V
f1–V
f5 in males and females, respectively.
2.6. Neuropil Volume Analyses
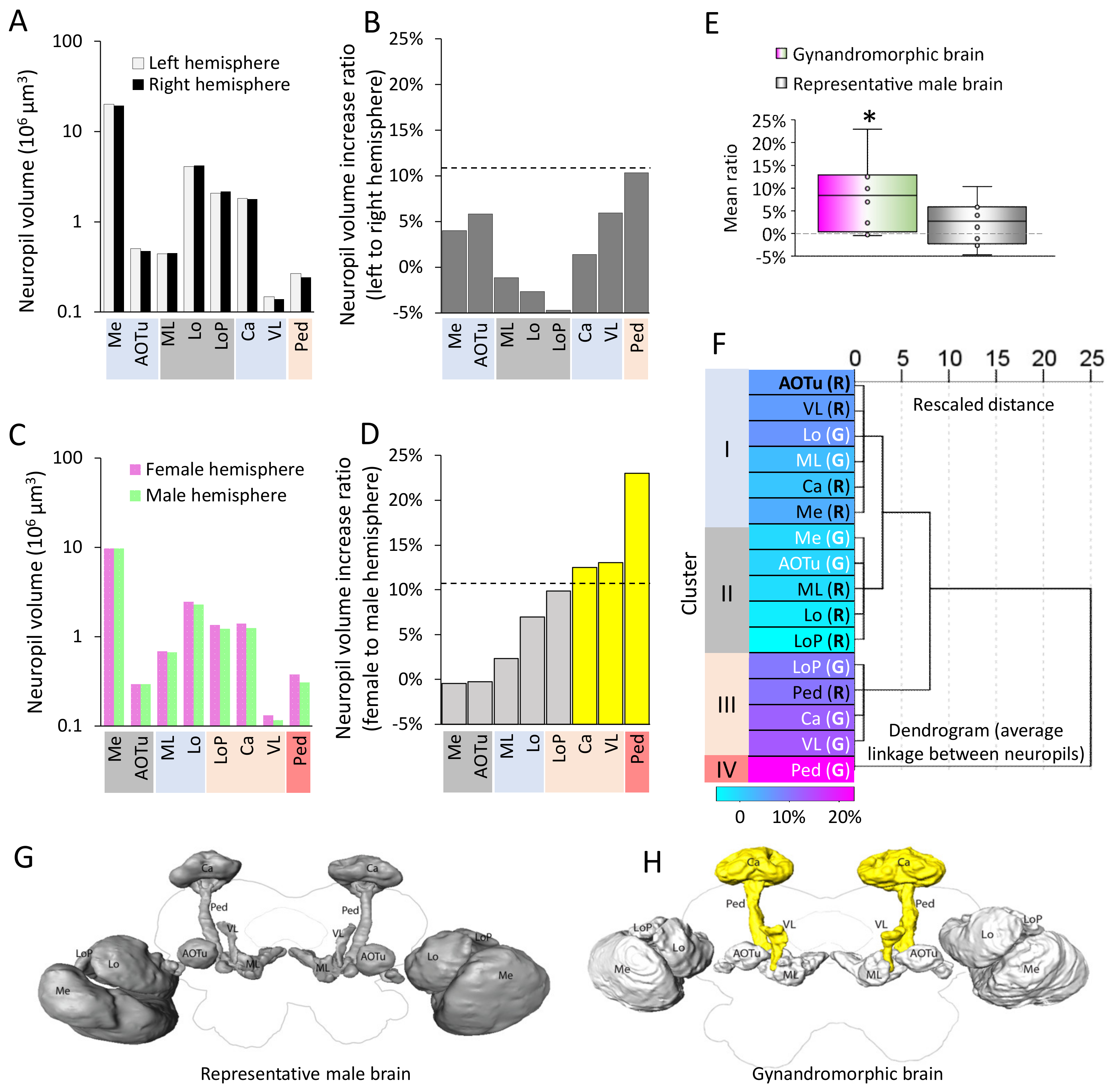
As gynandromorphic moths are rare, the analyses of the neuropile volume were conducted based on the comparison of data from one gynandromorphic brain and a male genotype representative brain. We used the hemispheric ratio to clarify the volumetric difference of corresponding neuropils between the two hemispheres. Thus, in the representative brain, the hemispheric ratio of a neuropil was computed by comparing the volume of corresponding neuropils in the left and right hemisphere. For the gynandromorphic brain, this ratio was calculated by comparing the neuropil volumes of corresponding structures in the male and female halves. For example, the hemispheric ratio of the vertical lobe in the gynandromorphic brain was calculated as the increase rate of vertical lobe volume in the female hemisphere relative to the male hemisphere. To determine whether the hemispheric ratios of neuropils in the gynandromorphic brain were different from those in the representative brain, we defined a threshold (T) based upon the hemispheric ratio of eight corresponding neuropils in the representative brain at a 10% significance level. We also compared the mean hemispheric ratios (across 8 neuropils) to a theoretically symmetrical brain, in which the hemispheric ratio should be 0, by using a one-sample t test. In addition, to investigate whether the hemispheric ratio was related with the neuropil function, we generated a hierarchical cluster analysis by using the centroid clustering method to obtain an overview of the linkage of all hemispheric ratios from, totally, 16 neuropils, including 8 in the representative brain and 8 in the gynandromorphic brain. All probabilities given are two-tailed. SPSS, version 25, was used for the statistical analysis.
4. Discussion
In the present study, we investigated the brain of a gynandromorphic
H. armigera moth to uncover putative sex-specific neuropil regions. Comparison of the primary olfactory center, the AL, in normal males and females of this species has been conducted previously [
5,
6]; however, since neuropil volumes in different moth brains may depend on age, and pre- and post-eclosure experience, such comparisons across different individuals could reflect inter-individual variations rather than sex-specific distinctions. Thus, the gynandromorphic sample used in this study provided a remarkable opportunity to explore the putative sex differences in the same brain, where both the male and female hemispheres developed under the same conditions.
Confocal imaging and subsequent 3D reconstruction of individual brain regions in the gynandromorphic sample allowed us to make a precise volume comparison between several pairs of olfactory neuropils in the male and female hemispheres including the antennal lobe, calyces, pedunculus, and lobes of the mushroom bodies. In addition, we also examined the optical lobes regarding possible sex differences.
The most prominent sex-specific feature in the moth brain is the MGC of the male AL [
5,
18]. A previous study on a gynandromorphic moth of the species,
Agrotis ipsilon (Hufnagel), demonstrated morphological differences within the brain and reproductive system [
18]. Here, however, the ALs in both the male and female parts were incompletely developed in comparison with the typical male/female genotype. Thus, the AL of the
A. ipsilon’s gynandromorphic brain included ordinary glomeruli in both hemispheres but no female-specific glomeruli in the female hemisphere and only a partially developed MGC in the male hemisphere. In our study on the gynandromorphic
H. armigera, on the other hand, both ALs seemed to be completely developed. The anatomical organization in the two brain hemispheres was in full correspondence with the ‘normal’ male and female individuals, respectively. This included the presence of five glomerular assemblies in both hemispheres, the MGC/Fx, OGs, PCx, LPOG, and VCx.
As shown in the results, the arrangement of glomerular assemblies in the two brain halves of the gynandromorphic brain were comparable, with one exception—a sex-specific group formed by the MGC and Fx in the male and female hemispheres, respectively. This is in full agreement with previous data on this species reporting about a sex-specific group of glomeruli located at the antennal-nerve entrance into the AL [
5,
6,
17].
Concerning the largest group of glomeruli, the OGs, comparison between the ALs of the gynandromorph
H. armigera and a normal type showed a significant difference. In this study, there was a total number of 47 and 50 OGs in the male and female hemispheres, respectively; however, in the previous studies on
H. armigera, [
5,
6], the respective numbers were 66 and 61 [
5,
6]. The relatively large difference between the numbers of identified glomeruli could be due to the different staining procedures applied. While only synapse-associated protein immunostaining was used in the present study, the former investigations combined multiple staining methods including two distinct immunostaining techniques, anterograde staining of the sensory axons, and retrograde staining of AL output neurons. The previous reports therefore allowed a staining quality probably visualizing distinct glomeruli in the deep parts of the AL-glomeruli that might not be recognized in this study. Moreover, the definition of the VCx glomeruli as a separate group in the present study, which were previously included in the OGs, leads to a lower number as well.
In addition to the anatomical difference in one part of the male and female AL of the gynandromorph, we found volumetric distinctions between the protocerebral neuropils in the two hemispheres. Altogether, the high resolution of the confocal images made it possible to reconstruct three optical lobe neuropils (medulla, lobula, and lobula plate) and five protocerebral neuropils (MB calyces, pedunculus, vertical lobe, medial lobes, and the AOTu). This contrasts with the fact that none of the corresponding neuropil pairs labeled in the representative brain had hemispheric ratios above the threshold. The three MB neuropil pairs exceeding the hemispheric ratio threshold in the gynandromorphic brain were the calyces, vertical lobe, and pedunculus.
Interestingly, the female hemisphere in the gynandromorph demonstrated enlarged volumes of neuropils within the MBs, a higher order region for olfactory memory formation. Like in other insects, the MBs of the moth are formed by several morphological types of Kenyon cells (KC) having distinct anatomical traits within the Ca and the MB lobes [
19,
20]. Notably, the KCs are assumed to play a role in adjusting odor representation according to experience [
21]. The difference between the volumes of certain MB compartments as obtained in this study may indicate the involvement of distinct circuits for processing biologically relevant signals in males and females, respectively. While the male moth mostly orients for detecting and finding a suitable conspecific female, the female has to recognize suitable host plants for oviposition—a task involving appropriate behavior according to the ecological surroundings. This may be reflected in a bigger number of the KCs in the female, leading to the ability to identify, learn, and remember a large amount of potentially suitable plant odors. Therefore, the different volumes of the MB structures in the two hemispheres of the gynandromorph may be related to an increased number of KCs in the female part satisfying the special requirements during oviposition. While volumetric differences between the MB structures of male and female moths have not been previously reported, sex-specific projection patterns within the calyces are well documented. Thus, in males, projection neurons (PNs) originating from the MGC and the OGs are reported to terminate within distinct areas of the calyces whereas females seem to have no organized patterns of PNs within this neuropil structure [
14,
22,
23].
In conclusion, the data from the gynandromorph moth investigated here, not only confirmed the previous findings about sexual dimorphism in the primary olfactory center of the moth brain, the AL, but also in the higher centers like the calyces, pedunculus, and vertical lobe of the mushroom body. It is worth noting that there is a limitation in our conclusion, in particular that the statistical analyses were conducted in only two individual brains, due to the rareness of a gynandromorphic brain; however, the difference between the male and female brains we revealed in the gynandromorphic brain sample is representative.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}