
Genome-Wide Identification of the Long Noncoding RNAs of Tribolium castaneum in Response to Terpinen-4-ol Fumigation

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Culture and Treatment
2.2. RNA Extraction, Library Establishment, and Sequencing
2.3. lncRNA Bioinformatic Analysis
2.4. Coding Capacity and Express Analysis of lncRNA
2.5. Differential Gene Expression Analysis
2.6. Functional Annotation and Gene Enrichment Analysis
2.7. Prediction Targeted Genes and Families of lncRNAs
2.8. Real-Time Quantitative PCR Analysis
3. Results
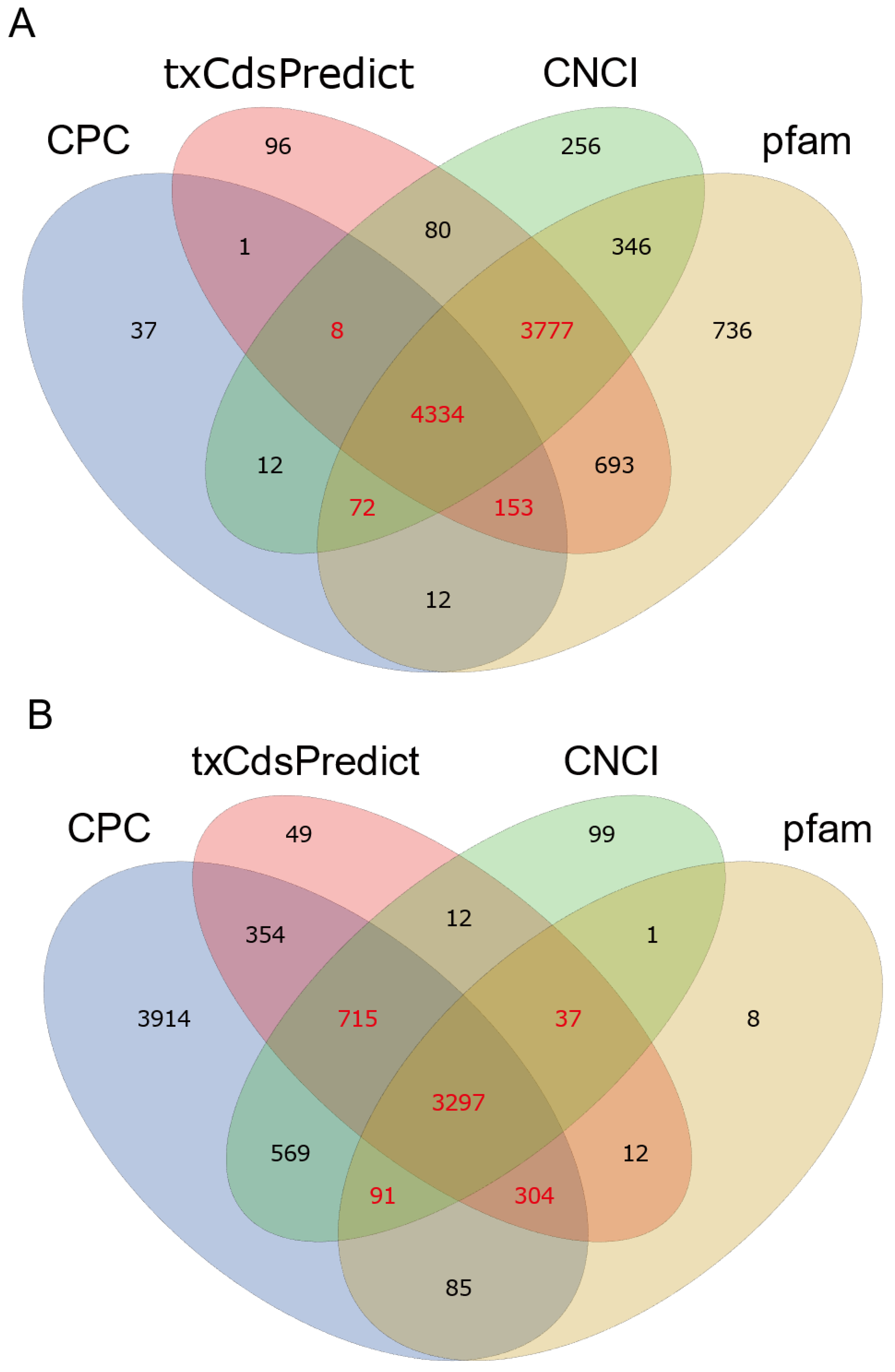
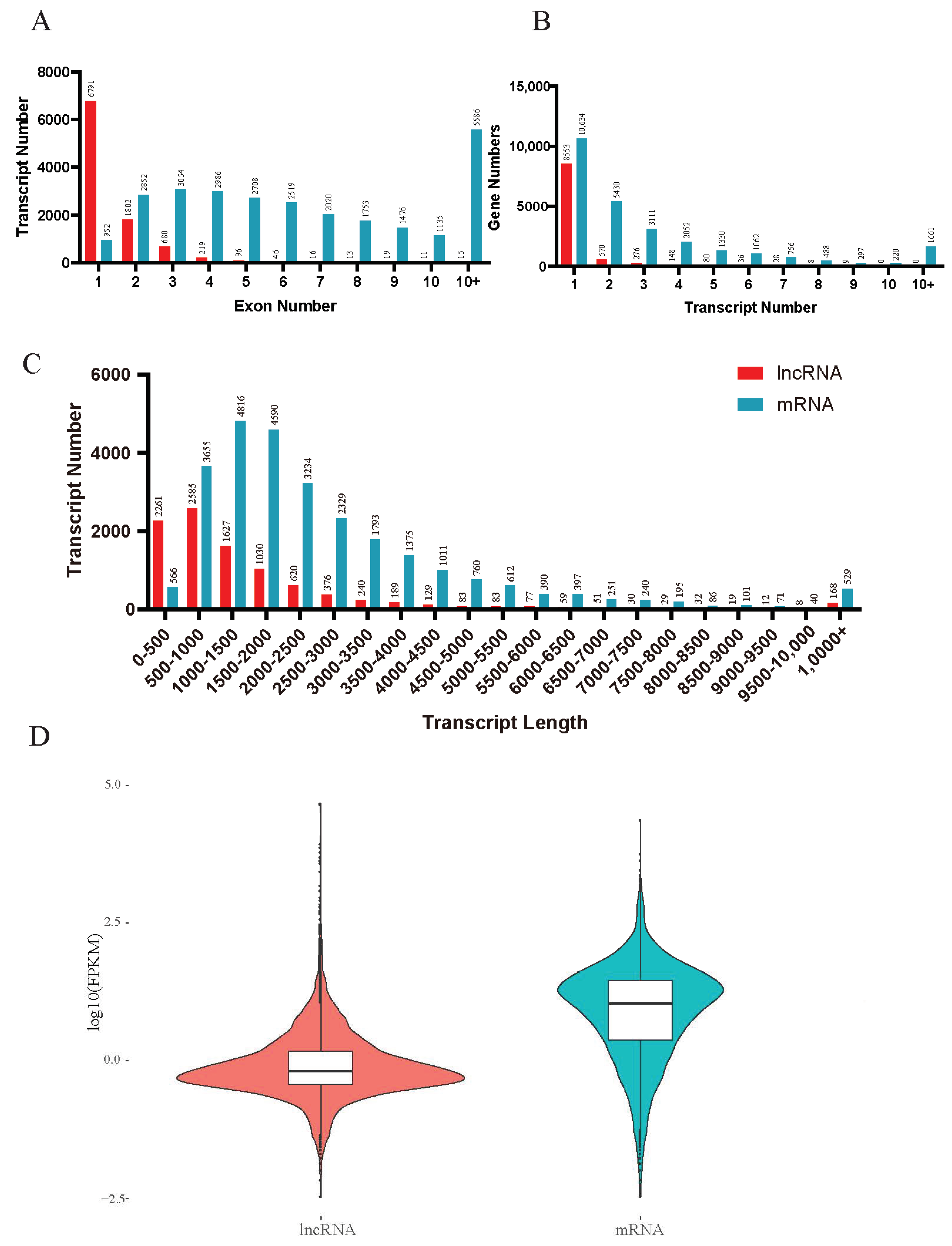
3.1. Identification and Characterization of the lncRNAs in Tribolium castaneum
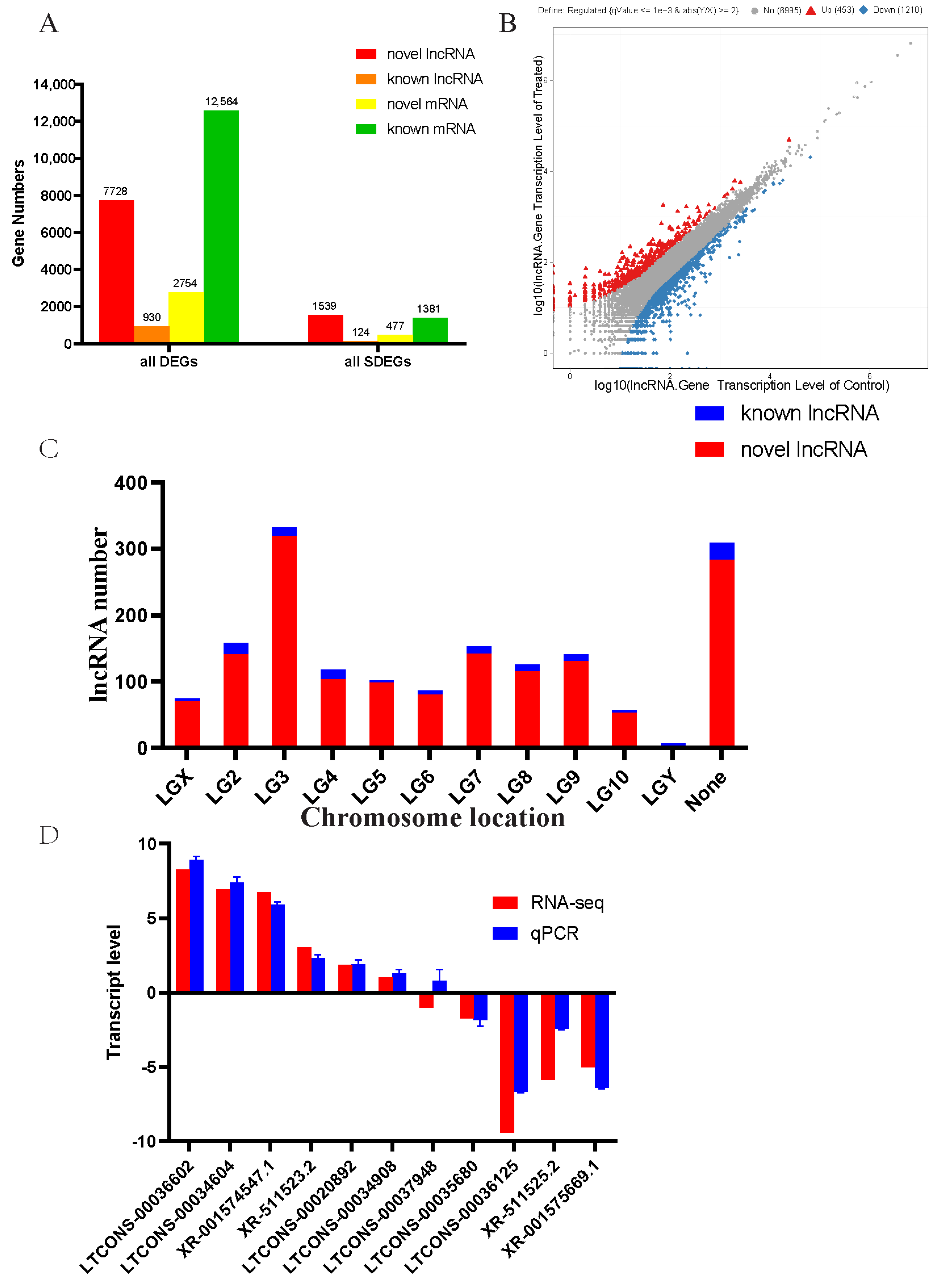
3.2. Analysis of Coding Capacity and Differentially Transcribed lncRNAs
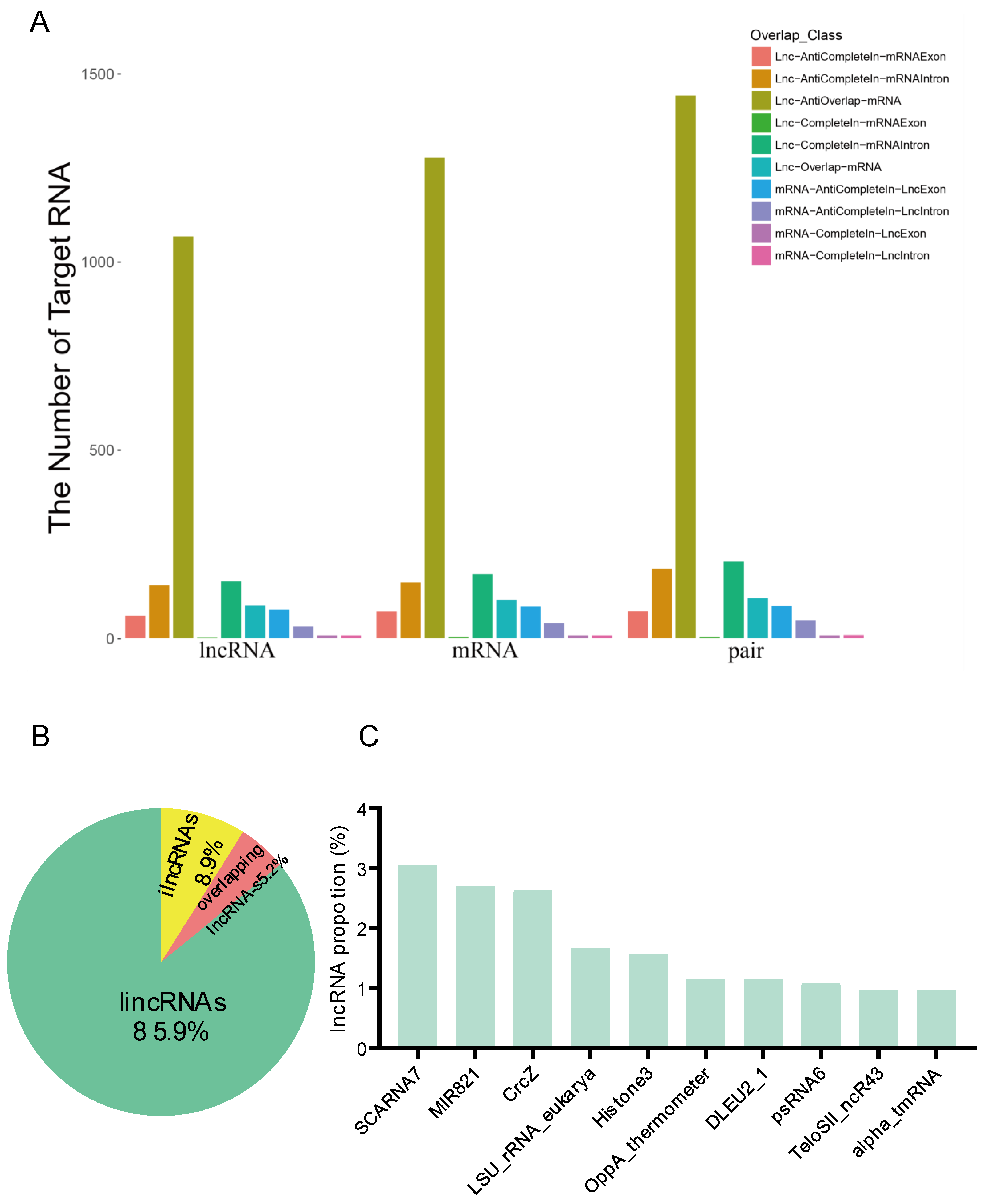
3.3. Targeted and Family Analysis of the Metabolism-Associated lncRNAs
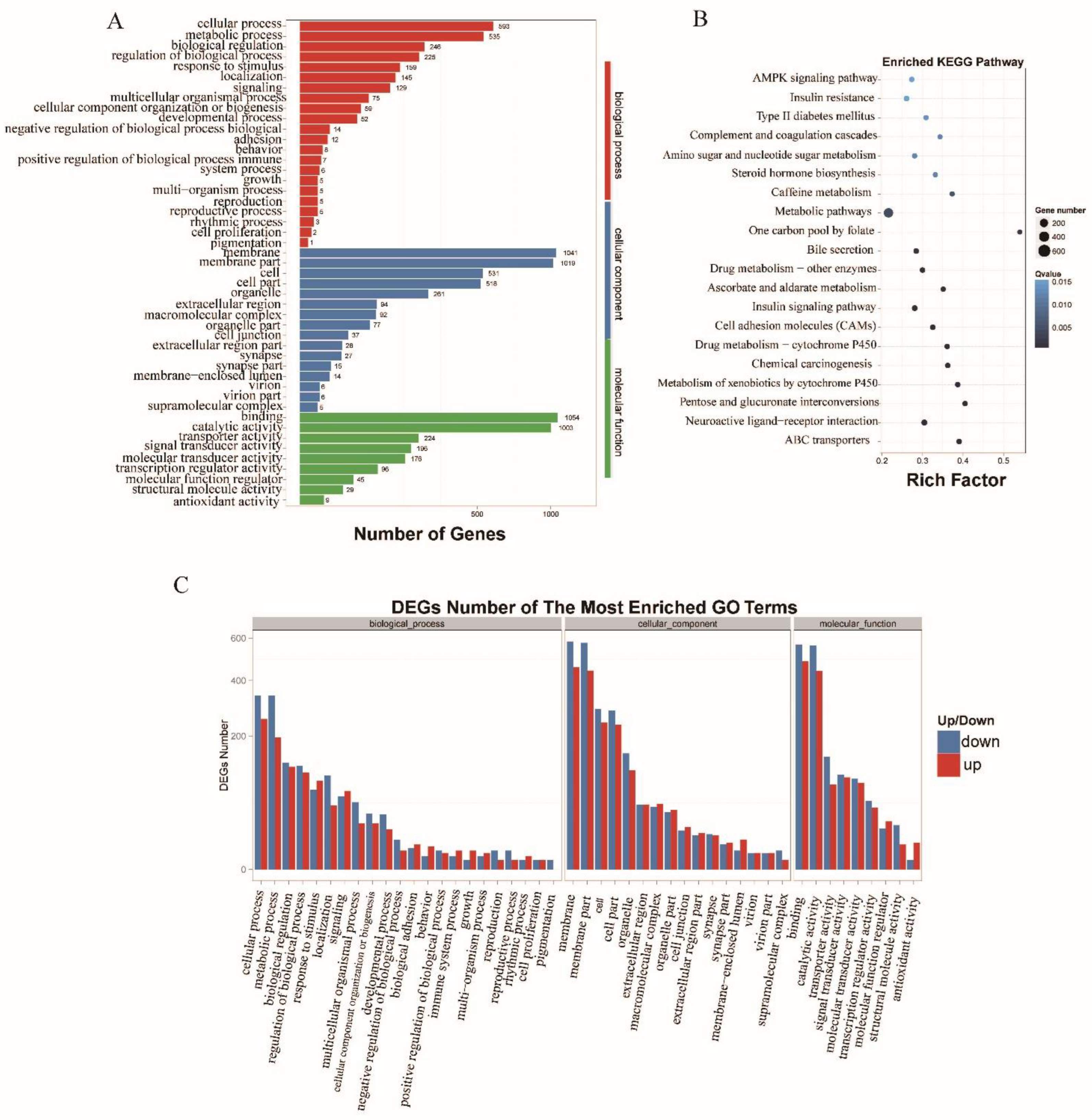
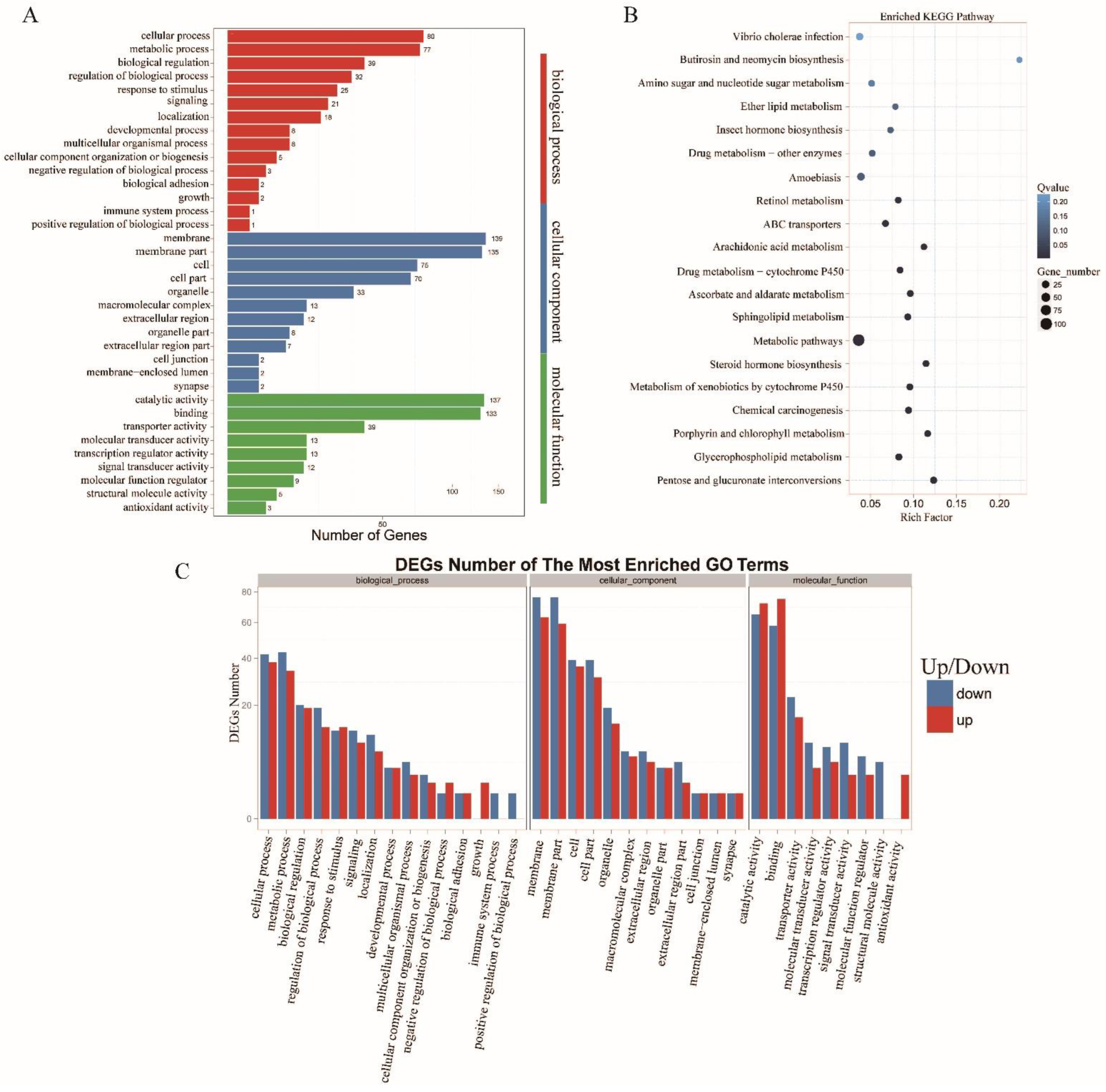
3.4. Functional Analysis and Enrichment of the Metabolism-Associated lncRNAs and Target Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grünwald, S.; Adam, I.V.; Gurmai, A.-M.; Bauer, L.; Boll, M.; Wenzel, U. The red flour beetle Tribolium castaneum as a model to monitor food safety and functionality. In Yellow Biotechnology I; Springer: Berlin/Heidelberg, Germany, 2013; pp. 111–122. [Google Scholar]
- Consortium, T.G.S. The Genome of the Model Beetle and Pest Tribolium castaneum. Nature 2008, 452, 949. [Google Scholar]
- Bergvinson, D.; García-Lara, S. Genetic Approaches to Reducing Losses of Stored Grain to Insects and Diseases. Curr. Opin. Plant Biol. 2004, 7, 480–485. [Google Scholar] [CrossRef]
- Zettler, L.J.; Cuperus, G.W. Pesticide Resistance in Tribolium castaneum (Coleoptera: Tenebrionidae) and Rhyzopertha dominica (Coleoptera: Bostrichidae) in Wheat. J. Econ. Entomol. 1990, 83, 1677–1681. [Google Scholar] [CrossRef]
- Collins, P.J. A New Resistance to Pyrethroids in Tribolium castaneum (Herbst). Pestic. Sci. 1990, 28, 101–115. [Google Scholar] [CrossRef]
- Hart, P.H.; Brand, C.; Carson, C.F.; Riley, T.V.; Prager, R.H.; Finlay-Jones, J.J. Terpinen-4-ol, the Main Component of the Essential Oil of Melaleuca alternifolia (Tea Tree Oil), Suppresses Inflammatory Mediator Production by Activated Human Monocytes. Inflamm. Res. 2000, 49, 619–626. [Google Scholar] [CrossRef]
- Calcabrini, A.; Stringaro, A.; Toccacieli, L.; Meschini, S.; Marra, M.; Colone, M.; Arancia, G.; Molinari, A.; Salvatore, G.; Mondello, F. Terpinen-4-ol, the Main Component of Melaleuca alternifolia (Tea Tree) Oil Inhibits the in Vitro Growth of Human Melanoma Cells. J. Investig. Dermatol. 2004, 122, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Brilhante, R.S.N.; Caetano, É.P.; Lima, R.A.C.D.; Marques, F.J.D.F.; Castelo-Branco, D.D.S.C.M.; Melo, C.V.S.D.; Guedes, G.M.D.M.; Oliveira, J.S.D.; Camargo, Z.P.D.; Moreira, J.L.B.; et al. Terpinen-4-ol, Tyrosol, and β-Lapachone as Potential Antifungals against Dimorphic Fungi. Braz. J. Microbiol. 2016, 47, 917–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morcia, C.; Malnati, M.; Terzi, V. In Vitro Antifungal Activity of Terpinen-4-ol, Eugenol, Carvone, 1, 8-Cineole (Eucalyptol) and Thymol against Mycotoxigenic Plant Pathogens. Food Addit. Contam. Part A 2012, 29, 415–422. [Google Scholar]
- Walton, S.F.; McKinnon, M.; Pizzutto, S.; Dougall, A.; Williams, E.; Currie, B.J. Acaricidal Activity of Melaleuca alternifolia (Tea Tree) Oil: In Vitro Sensitivity of Sarcoptes scabiei Var Hominis to Terpinen-4-ol. Arch. Dermatol. 2004, 140, 563–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tighe, S.; Gao, Y.-Y.; Tseng, S.C.G. Terpinen-4-ol Is the Most Active Ingredient of Tea Tree Oil to Kill Demodex Mites. Transl. Vis. Sci. Technol. 2013, 2, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Liao, M.; Yang, Q.; Shi, S.; Xiao, J.; Cao, H. Knockdown of NADPH-Cytochrome P450 Reductase and CYP6MS1 Increases the Susceptibility of Sitophilus zeamais to Terpinen-4-ol. Pestic. Biochem. Physiol. 2020, 162, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Yeom, H.-J.; Kang, J.; Kim, S.-W.; Park, I.-K. Fumigant and Contact Toxicity of Myrtaceae Plant Essential Oils and Blends of Their Constituents against Adults of German Cockroach (Blattella germanica) and Their Acetylcholinesterase Inhibitory Activity. Pestic. Biochem. Physiol. 2013, 107, 200–206. [Google Scholar] [CrossRef]
- Kordali, S.; Aslan, I.; Çalmaşur, O.; Cakir, A. Toxicity of Essential Oils Isolated from Three Artemisia Species and Some of Their Major Components to Granary Weevil, Sitophilus granarius (L.) (Coleoptera: Curculionidae). Ind. Crops Prod. 2006, 23, 162–170. [Google Scholar] [CrossRef]
- Erler, F. Fumigant Activity of Six Monoterpenoids from Aromatic Plants in Turkey against the Two Stored-Product Pests Confused Flour Beetle, Tribolium confusum, and Mediterranean Flour Moth, Ephestia kuehniella. J. Plant Dis. Prot. 2005, 112, 602–611. [Google Scholar] [CrossRef]
- Suthisut, D.; Fields, P.G.; Chandrapatya, A. Fumigant Toxicity of Essential Oils from Three Thai Plants (Zingiberaceae) and Their Major Compounds against Sitophilus zeamais, Tribolium castaneum and Two Parasitoids. J. Stored Prod. Res. 2011, 47, 222–230. [Google Scholar] [CrossRef]
- Reidy, G.F.; Rose, H.A.; Visetson, S.; Murray, M. Increased Glutathione S-Transferase Activity and Glutathione Content in an Insecticide-Resistant Strain of Tribolium castaneum (Herbst). Pestic. Biochem. Physiol. 1990, 36, 269–276. [Google Scholar] [CrossRef]
- Zhu, F.; Parthasarathy, R.; Bai, H.; Woithe, K.; Kaussmann, M.; Nauen, R.; Harrison, D.A.; Palli, S.R. A Brain-Specific Cytochrome P450 Responsible for the Majority of Deltamethrin Resistance in the QTC279 Strain of Tribolium castaneum. Proc. Natl. Acad. Sci. USA 2010, 107, 8557–8562. [Google Scholar] [CrossRef] [Green Version]
- Andreev, D.; Kreitman, M.; Phillips, T.W.; Beeman, R.W. Multiple Origins of Cyclodiene Insecticide Resistance in Tribolium castaneum (Coleoptera: Tenebrionidae). J. Mol. Evol. 1999, 48, 615–624. [Google Scholar] [CrossRef]
- Huang, Y.; Liao, M.; Yang, Q.; Xiao, J.; Hu, Z.; Zhou, L.; Cao, H. Transcriptome Profiling Reveals Differential Gene Expression of Detoxification Enzymes in Sitophilus zeamais Responding to Terpinen-4-ol Fumigation. Pestic. Biochem. Physiol. 2018, 149, 44–53. [Google Scholar] [CrossRef]
- Liao, M.; Xiao, J.-J.; Zhou, L.-J.; Liu, Y.; Wu, X.-W.; Hua, R.-M.; Wang, G.-R.; Cao, H.-Q. Insecticidal Activity of Melaleuca alternifolia Essential Oil and RNA-Seq Analysis of Sitophilus zeamais Transcriptome in Response to Oil Fumigation. PLoS ONE 2016, 11, e0167748. [Google Scholar] [CrossRef] [Green Version]
- Jathar, S.; Kumar, V.; Srivastava, J.; Tripathi, V. Technological developments in lncRNA biology. In Long Non Coding RNA Biology; Springer: Berlin, Germany, 2017; pp. 283–323. [Google Scholar]
- Khorkova, O.; Hsiao, J.; Wahlestedt, C. Basic Biology and Therapeutic Implications of LncRNA. Adv. Drug Deliv. Rev. 2015, 87, 15–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, R.; Li, H.; Zhang, H.; An, S. Comparative Analysis of DsRNA-induced LncRNAs in Three Kinds of Insect Species. Arch. Insect Biochem. Physiol. 2020, 103, e21640. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Yuan, Z.; Guo, D.; Hou, B.; Yin, C.; Zhang, W.; Li, F. Genome-Wide Identification of Long Noncoding RNA Genes and Their Potential Association with Fecundity and Virulence in Rice Brown Planthopper, Nilaparvata lugens. BMC Genom. 2015, 16, 749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Cheng, T.; Liu, C.; Liu, D.; Zhang, Q.; Long, R.; Zhao, P.; Xia, Q. Systematic Identification and Characterization of Long Non-Coding RNAs in the Silkworm, Bombyx mori. PLoS ONE 2016, 11, e0147147. [Google Scholar] [CrossRef] [Green Version]
- Etebari, K.; Asad, S.; Zhang, G.; Asgari, S. Identification of Aedes aegypti Long Intergenic Non-Coding RNAs and Their Association with Wolbachia and Dengue Virus Infection. PLoS Negl. Trop. Dis. 2016, 10, e0005069. [Google Scholar] [CrossRef]
- Azlan, A.; Obeidat, S.M.; Yunus, M.A.; Azzam, G. Systematic Identification and Characterization of Aedes aegypti Long Noncoding RNAs (LncRNAs). Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, Y.; Zhang, X.; Jia, S.; Chen, S.; Kang, L. Genome-Wide Identification and Developmental Expression Profiling of Long Noncoding RNAs during Drosophila metamorphosis. Sci. Rep. 2016, 6, 1–8. [Google Scholar]
- Zhang, J.; Yang, Z.; Feng, P.; Zhong, X.; Ma, Q.; Su, Q.; Wang, X.; Li, C.; Yang, Y. Identification and the Potential Roles of Long Non-Coding RNAs in Cotton Leaves Damaged by Aphis gossypii. Plant Growth Regul. 2019, 88, 215–225. [Google Scholar] [CrossRef]
- Li, W.-J.; Song, Y.-J.; Han, H.-L.; Xu, H.-Q.; Wei, D.; Smagghe, G.; Wang, J.-J. Genome-Wide Analysis of Long Non-Coding RNAs in Adult Tissues of the Melon Fly, Zeugodacus cucurbitae (Coquillett). BMC Genom. 2020, 21, 1–12. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, T.; He, W.; Shen, X.; Zhao, Q.; Bai, J.; You, M. Genome-Wide Identification and Characterization of Putative LncRNAs in the Diamondback Moth, Plutella xylostella (L.). Genomics 2018, 110, 35–42. [Google Scholar] [CrossRef]
- Liu, F.; Guo, D.; Yuan, Z.; Chen, C.; Xiao, H. Genome-Wide Identification of Long Non-Coding RNA Genes and Their Association with Insecticide Resistance and Metamorphosis in Diamondback Moth, Plutella xylostella. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etebari, K.; Furlong, M.J.; Asgari, S. Genome Wide Discovery of Long Intergenic Non-Coding RNAs in Diamondback Moth (Plutella xylostella) and Their Expression in Insecticide Resistant Strains. Sci. Rep. 2015, 5, 14642. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Xu, M.; Shi, H.; Gao, X.; Liang, P. Genome-Wide Identification of LncRNAs Associated with Chlorantraniliprole Resistance in Diamondback Moth Plutella xylostella (L.). BMC Genom. 2017, 18, 1–11. [Google Scholar] [CrossRef]
- Yang, L.; Wang, Y.; Lu, Y.; Li, B.; Chen, K.; Li, C. Genome-wide Identification and Characterization of Long Non-coding RNAs in Tribolium castaneum. Insect Sci. 2020, 28, 1262–1276. [Google Scholar] [CrossRef] [PubMed]
- Satyavathi, V.; Ghosh, R.; Subramanian, S. Long Non-Coding RNAs Regulating Immunity in Insects. Non-Coding RNA 2017, 3, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Hussain, F.; Unnithan, G.C.; Dong, S.; UlAbdin, Z.; Gu, S.; Mathew, L.G.; Fabrick, J.A.; Ni, X.; Carrière, Y. A Long Non-Coding RNA Regulates Cadherin Transcription and Susceptibility to Bt Toxin Cry1Ac in Pink Bollworm, Pectinophora gossypiella. Pestic. Biochem. Physiol. 2019, 158, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short Oligonucleotide Alignment Program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie Enables Improved Reconstruction of a Transcriptome from RNA-Seq Reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript Assembly and Quantification by RNA-Seq Reveals Unannotated Transcripts and Isoform Switching during Cell Differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Zhang, Y.; Ye, Z.-Q.; Liu, X.-Q.; Zhao, S.-Q.; Wei, L.; Gao, G. CPC: Assess the Protein-Coding Potential of Transcripts Using Sequence Features and Support Vector Machine. Nucleic Acids Res. 2007, 35, W345–W349. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Luo, H.; Bu, D.; Zhao, G.; Yu, K.; Zhang, C.; Liu, Y.; Chen, R.; Zhao, Y. Utilizing Sequence Intrinsic Composition to Classify Protein-Coding and Long Non-Coding Transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A. The Pfam Protein Families Database: Towards a More Sustainable Future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R Package for Identifying Differentially Expressed Genes from RNA-Seq Data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A Universal Tool for Annotation, Visualization and Analysis in Functional Genomics Research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Tafer, H.; Hofacker, I.L. RNAplex: A Fast Tool for RNA–RNA Interaction Search. Bioinformatics 2008, 24, 2657–2663. [Google Scholar] [CrossRef]
- Burge, S.W.; Daub, J.; Eberhardt, R.; Tate, J.; Barquist, L.; Nawrocki, E.P.; Eddy, S.R.; Gardner, P.P.; Bateman, A. Rfam 11.0: 10 Years of RNA Families. Nucleic Acids Res. 2013, 41, D226–D232. [Google Scholar] [CrossRef] [Green Version]
- Nawrocki, E.P.; Kolbe, D.L.; Eddy, S.R. Infernal 1.0: Inference of RNA Alignments. Bioinformatics 2009, 25, 1335–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Ajayi, O.E.; Guo, D.; Wu, Q. Genome-wide Characterization and Developmental Expression Profiling of Long Non-coding RNAs in Sogatella furcifera. Insect Sci. 2020, 27, 987–997. [Google Scholar] [CrossRef]
- Ma, Z.; Luan, Z.; Zhang, X. Effects of Terpinen-4-ol on Culex Pipiens pallens and Its Na+, K+-ATPase. Chin. J. Pestic. Sci. 2009, 11, 230–234. [Google Scholar]
- Guo, Z.-B.; Feng, J.; Zhang, X. Inhibition of Na+, K+-ATPase in Housefly (Musca domestica L.) by Terpinen-4-ol and Its Ester Derivatives. Agric. Sci. China 2009, 8, 1492–1497. [Google Scholar] [CrossRef]
- Koch, R.B.; Cutkomp, L.K.; Do, F.M. Chlorinated Hydrocarbon Insecticide Inhibition of Cockroach and Honey Bee ATPases. Life Sci. 1969, 8, 289–297. [Google Scholar] [CrossRef]
- Coats, J.R.; Karr, L.L.; Drewes, C.D. Toxicity and Neurotoxic Effects of Monoterpenoids: In Insects and Earthworms; ACS Publications: Washington, DC, USA, 1991; ISBN 1947-5918. [Google Scholar]
- Priestley, C.M.; Williamson, E.M.; Wafford, K.A.; Sattelle, D.B. Thymol, a Constituent of Thyme Essential Oil, Is a Positive Allosteric Modulator of Human GABAA Receptors and a Homo-oligomeric GABA Receptor from Drosophila melanogaster. Br. J. Pharmacol. 2003, 140, 1363–1372. [Google Scholar] [CrossRef] [Green Version]
- Enan, E.E. Molecular Response of Drosophila melanogaster Tyramine Receptor Cascade to Plant Essential Oils. Insect Biochem. Mol. Biol. 2005, 35, 309–321. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, M.; Kim, H.; Kim, K. Effect of Lavender Oil on Motor Function and Dopamine Receptor Expression in the Olfactory Bulb of Mice. J. Ethnopharmacol. 2009, 125, 31–35. [Google Scholar] [CrossRef]
- Kostyukovsky, M.; Rafaeli, A.; Gileadi, C.; Demchenko, N.; Shaaya, E. Activation of Octopaminergic Receptors by Essential Oil Constituents Isolated from Aromatic Plants: Possible Mode of Action against Insect Pests. Pest Manag. Sci. 2002, 58, 1101–1106. [Google Scholar] [CrossRef]
- Rombolà, L.; Scuteri, D.; Watanabe, C.; Sakurada, S.; Hamamura, K.; Sakurada, T.; Tonin, P.; Corasaniti, M.T.; Bagetta, G.; Morrone, L.A. Role of 5-HT1A Receptor in the Anxiolytic-Relaxant Effects of Bergamot Essential Oil in Rodent. Int. J. Mol. Sci. 2020, 21, 2597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Himmel, N.J.; Letcher, J.M.; Sakurai, A.; Gray, T.R.; Benson, M.N.; Cox, D.N. Drosophila Menthol Sensitivity and the Precambrian Origins of Transient Receptor Potential-Dependent Chemosensation. Philos. Trans. R. Soc. B 2019, 374, 20190369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankowska, M.; Rogalska, J.; Wyszkowska, J.; Stankiewicz, M. Molecular Targets for Components of Essential Oils in the Insect Nervous System—A Review. Molecules 2018, 23, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-H.; Ha, K.B.; Park, D.H.; Fang, Y.; Kim, J.H.; Park, M.G.; Woo, R.M.; Kim, W.J.; Park, I.-K.; Choi, J.Y. Plant-Derived Compounds Regulate Formation of the Insect Juvenile Hormone Receptor Complex. Pestic. Biochem. Physiol. 2018, 150, 27–32. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specific Primer Names | Sequence (5′-3′) | Amplification Size (bp) |
|---|---|---|
| RPL18-F | CGACCAAAGGATATGGGATG | 198 |
| RPL18-R | GGACCAAAATGTTTCACTGCT | |
| qLTCONS_00036602-F | GTTCGGACATTTGGTTCAC | 80 |
| qLTCONS_00036602-R | AGGCGTTCAGGCATAATC | |
| qLTCONS_00034604-F | CACCATAGGACTCCAGTT | 111 |
| qLTCONS_00034604-R | CAGGTAGGTCAGTTGTCA | |
| qXR_001574547.1-F | GGTCTTGAAGTGTCTTGATG | 81 |
| qXR_001574547.1-R | TGAATATAACGGCGGAGAG | |
| qXR_511523.2-F | CAAAGGCGGAGAGTTTATG | 96 |
| qXR_511523.2-R | TAAGCGACTGTGGGAAATC | |
| qLTCONS_00020892-F | GGGTCAAGACTCACTTTTG | 83 |
| qLTCONS_00020892-R | GTGTCAGTGTCCTAACCT | |
| LTCONS_00034908-F | GTCTGTCATTCCTTCCAT | 96 |
| LTCONS_00034908-R | CTTACCCGTTTCACTTTC | |
| qLTCONS_00037948-F | GCCTGGAAAGAACAAGAAG | 88 |
| qLTCONS_00037948-R | TACTCTCACCTCATCTCACT | |
| qLTCONS_00035680-F | GTTTGAGGGCAGTAATGTC | 133 |
| qLTCONS_00035680-R | TTCGGTAGTCTTCCTTGTC | |
| qLTCONS_00036125-F | GACCTGTCCTGTTGATTC | 101 |
| qLTCONS_00036125-R | CAGCATCTCCTCTTTCAC | |
| qXR_511525.2-F | CGTTCCGAATGTATGATGAC | 96 |
| qXR_511525.2-R | GGCTGCGATGAGATAGTT | |
| qXR_001575669.1-F | TACGACAGCATCATCTACAG | 87 |
| qXR_001575669.1-R | CACGGCGATATTCCTTGA |
| Sample | Total Clean Reads | Total Mapping Ratio | Uniquely Mapping Ratio |
|---|---|---|---|
| Control_1 | 79,106,542 | 75.93% | 74.14% |
| Control_2 | 97,915,032 | 71.73% | 70.04% |
| Control_3 | 76,002,752 | 74.04% | 72.29% |
| Treated_1 | 82,392,022 | 71.54% | 69.97% |
| Treated_2 | 105,768,982 | 67.30% | 65.79% |
| Treated_3 | 75,888,294 | 72.29% | 70.60% |
| Sample | Novel_lncRNA Transcripts | Novel_mRNA Transcripts | Known_lncRNA Transcripts | Known_mRNA Transcripts |
|---|---|---|---|---|
| Control_1 | 7329 | 2648 | 862 | 12,346 |
| Control_2 | 7215 | 2621 | 832 | 12,308 |
| Control_3 | 7281 | 2622 | 843 | 12,318 |
| Treated_1 | 7132 | 2599 | 804 | 12,242 |
| Treated_2 | 7065 | 2605 | 811 | 12,266 |
| Treated_3 | 7194 | 2611 | 827 | 12,322 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Yue, S.; Huang, Y.; Zhao, X.; Cao, H.; Liao, M. Genome-Wide Identification of the Long Noncoding RNAs of Tribolium castaneum in Response to Terpinen-4-ol Fumigation. Insects 2022, 13, 283. https://doi.org/10.3390/insects13030283
Wu H, Yue S, Huang Y, Zhao X, Cao H, Liao M. Genome-Wide Identification of the Long Noncoding RNAs of Tribolium castaneum in Response to Terpinen-4-ol Fumigation. Insects. 2022; 13(3):283. https://doi.org/10.3390/insects13030283
Chicago/Turabian StyleWu, Hailong, Shuaili Yue, Yong Huang, Xinping Zhao, Haiqun Cao, and Min Liao. 2022. "Genome-Wide Identification of the Long Noncoding RNAs of Tribolium castaneum in Response to Terpinen-4-ol Fumigation" Insects 13, no. 3: 283. https://doi.org/10.3390/insects13030283