
Scanning Electron Microscopic Analysis of Antennal Sensilla and Tissue-Expression Profiles of Chemosensory Protein Genes in Ophraella communa (Coleoptera: Chrysomelidae)

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Scanning Electron Microscopy
2.3. CSP Cloning and Tissue-Expression Profiles
2.4. Data Analysis
3. Results
3.1. Antennae Description and Sensilla Identification
3.1.1. Sensilla Trichodea (ST)
Sensilla Trichodea 1 (ST1)
Sensilla Trichodea 2 (ST2)
3.1.2. Sensilla Chaetica (SCH)
3.1.3. Sensila Basiconinca (SB)
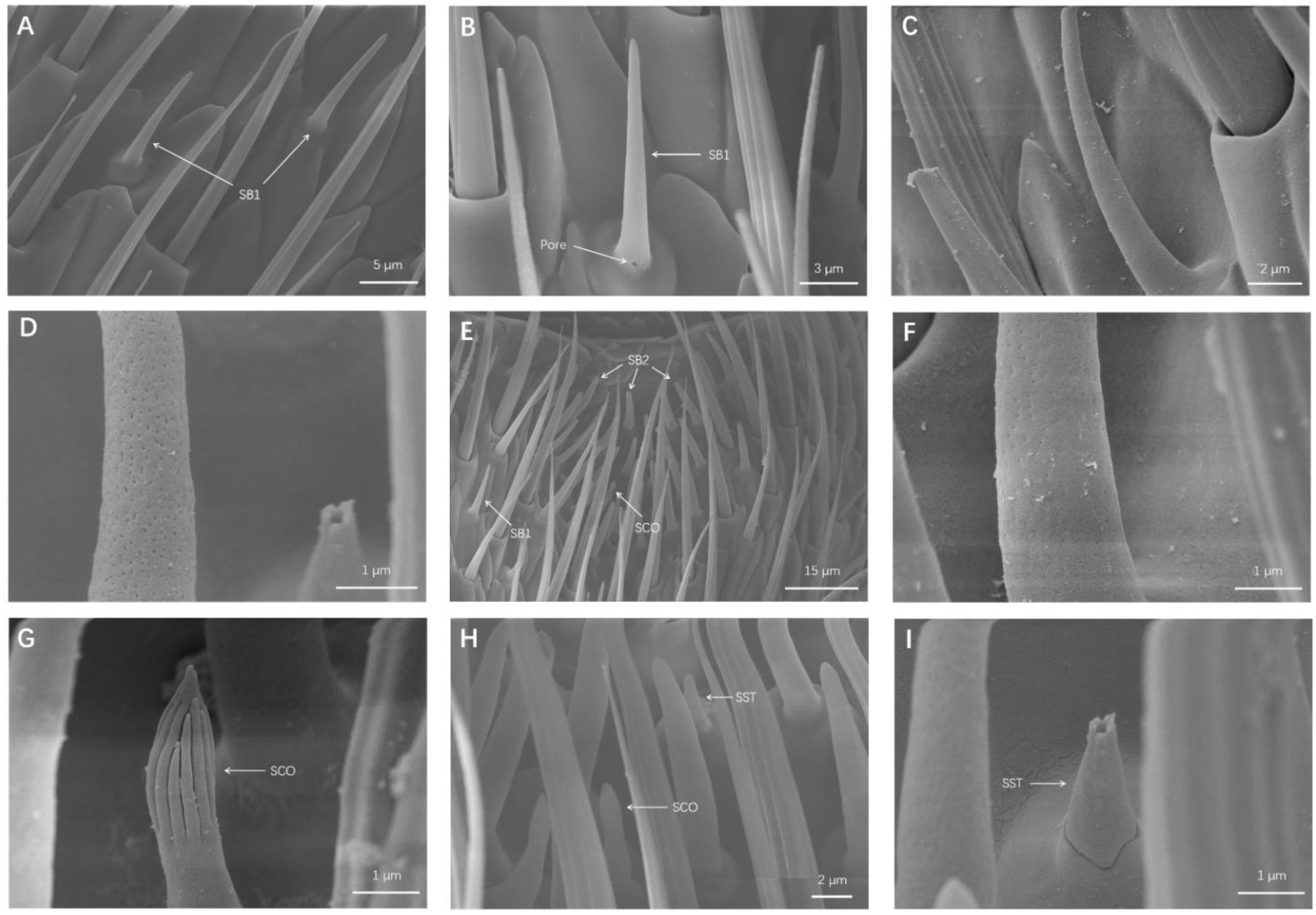
Sensila Basiconinca 1 (SB1)
Sensila Basiconinca 2 (SB2)
3.1.4. Sensilla Coeloconica (SCO)
3.1.5. Sensilla Styloconica (SST)
3.1.6. Böhm Bristles (BB)
3.2. Chemosensory Proteins (CSPs) Clone and Tissue-Expression Profiles
3.2.1. CSPs Cloning and Similarity Analysis
3.2.2. CSP Tissue-Expression Profiles
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leal, W.S. Odorant Reception in Insects: Roles of Receptors, Binding Proteins, and Degrading Enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Shields, V.D.C. Ultrastructure of the Uniporous Sensilla on the Galea of Larval Mamestra configurata (Walker) (Lepidoptera: Noctuidae). Can. J. Zool. 1994, 72, 2016–2031. [Google Scholar] [CrossRef]
- Li, Y.P.; Du, X.; Liu, F.F.; Li, Y.; Liu, T.X. Ultrastructure of the Sensilla on Antennae and Mouthparts of Larval and Adult Plutella xylostella (Lepidoptera: Plutellidae). J. Integr. Agric. 2018, 17, 1409–1420. [Google Scholar] [CrossRef] [Green Version]
- Zacharuk, R.Y. Antennae and Sensilla. In Comprehensive Insect Physiology, Biochemistry and Pharmacology; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon Press: Oxford, UK, 1985. [Google Scholar]
- Zacharuk, R.Y.; Shields, V.D. Sensilla of Immature Insects. Annu. Rev. Entomol. 1991, 36, 331–354. [Google Scholar] [CrossRef]
- Zhang, L.; Ren, L.L.; Luo, Y.Q.; Zong, S.X. Scanning Electron Microscopy Analysis of the Cephalic Sensilla of Chrysolina aeruginosa Fald. (Coleoptera, Chrysomelidae). Microsc. Res. Tech. 2013, 76, 423–431. [Google Scholar] [CrossRef]
- Waris, M.I.; Younas, A.; Adeel, M.M.; Duan, S.G.; Quershi, S.R.; Kaleem Ullah, R.M.; Wang, M.Q. The Role of Chemosensory protein 10 in the Detection of Behaviorally Active Compounds in Brown Planthopper, Nilaparvata lugens. Insect Sci. 2020, 27, 531–544. [Google Scholar] [CrossRef]
- Peng, X.; Liu, L.; Huang, Y.X.; Wang, S.J.; Li, D.X.; Chen, S.T.; Simon, J.C.; Qu, M.J.; Chen, M.H. Involvement of Chemosensory Proteins in Host Plant Searching in the Bird Cherry-Oat Aphid. Insect Sci. 2021, 28, 1338–1353. [Google Scholar] [CrossRef]
- Younas, A.; Waris, M.I.; Shaaban, M.; Tahir Ul Qamar, M.; Wang, M.Q. Appraisal of MsepCSP14 for Chemosensory Functions in Mythimna Separata. Insect Sci. 2021. [Google Scholar] [CrossRef]
- Palmer, W.A.; Goeden, R.D. The Host Range of Ophraella communa Lesage (Coleoptera: Chrysomelidae). Coleopt. Bull. 1991, 45, 115–120. [Google Scholar]
- Zhou, Z.S.; Chen, H.S.; Zheng, X.W.; Guo, J.Y.; Guo, W.; Li, M.; Luo, M.; Wan, F. Control of the Invasive Weed Ambrosia artemisiifolia With Ophraella communa and Epiblema Strenuana. Biocontrol. Sci. Technol. 2014, 24, 950–964. [Google Scholar] [CrossRef]
- Zhou, Z.S.; Guo, J.Y.; Guo, W.; Wan, F.H. Synergistic Effects of Olfactory and Tactile Cues in Short–Range Mate Finding of Ophraella communa. Entomol. Exp. Appl. 2011, 138, 48–54. [Google Scholar] [CrossRef]
- Ma, C.; Cui, S.; Bai, Q.; Tian, Z.; Zhang, Y.; Chen, G.; Gao, X.; Tian, Z.; Chen, H.; Guo, J.; et al. Olfactory Co-Receptor Is Involved in Host Recognition and Oviposition in Ophraella communa (Coleoptera: Chrysomelidae). Insect Mol. Biol. 2020, 29, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Zhao, C.C.; Cui, S.W.; Zhang, Y.; Chen, G.M.; Chen, H.S.; Wan, F.H.; Zhou, Z.S. Identification of candidate chemosensory genes of Ophraella communa LeSage (Coleoptera: Chrysomelidae) Based on antennal transcriptome analysis. Sci. Rep. 2019, 9, 15551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.H.; Li, B.L.; Li, C.R. Morphology and Distribution of Antennal Sensilla of Female Phyllotreta striolata (Fabricius) (Coleoptera: Chrysomelidae). Microsc. Res. Tech. 2016, 79, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Cui, S.W.; Tian, Z.Y.; Zhang, Y.; Chen, G.M.; Gao, X.Y.; Tian, Z.Q.; Chen, H.S.; Guo, J.Y.; Zhou, Z.S. OcomCSP12, Chemosensory Protein Expressed Specifically by Ovary, Mediates Reproduction in Ophraella communa (Coleoptera: Chrysomelidae). Front. Physiol. 2019, 10, 1290. [Google Scholar] [CrossRef] [Green Version]
- Seada, M.A.; Hamza, A.M. Differential Morphology of the Sensory Sensilla of Antennae, Palpi, Foretarsi and Ovipositor of Adult Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae). Ann. Agric. Sci. 2018, 63, 1–8. [Google Scholar] [CrossRef]
- Hao, Y.N.; Sun, Y.X.; Liu, C.Z. Functional Morphology of Antennae and Sensilla of Hippodamia variegata (Coleoptera: Coccinellidae). PLoS ONE 2020, 15, e0237452. [Google Scholar] [CrossRef]
- MacKay, C.A.; Sweeney, J.D.; Hillier, N.K. Morphology of Antennal Sensilla of the Brown Spruce Longhorn Beetle, Tetropium fuscum (Fabr.) (Coleoptera: Cerambycidae). Arthropod Struct. Dev. 2014, 43, 469–475. [Google Scholar] [CrossRef]
- Lei, G.; Fu, Y.; Wu, W.J. Type and Distribution of Sensilla in the Antennae of Euplatypus parallelus (F.) (Coleoptera: Curculionidea, Platypodinae). PLoS ONE 2020, 15, e0241275. [Google Scholar]
- Shi, X.; Zhang, S.F.; Liu, F.; Xu, F.Y.; Zhang, F.B.; Guo, X.B.; Zhang, Z.; Kong, X.B. SEM Analysis of Sensilla on the Mouthparts and Antennae of Asian Larch Bark Beetle Ips Subelongatus. Micron 2021, 140, 102976. [Google Scholar] [CrossRef]
- Merivee, E.; Ploomi, A.; Rahi, M.; Bresciani, J.; Ravn, H.P.; Luik, A.; Sammelselg, V. Antennal Sensilla of the Ground Beetle Bembidionpro perans Steph. (Coleoptera, Carabidae). Micron 2002, 33, 429–440. [Google Scholar] [CrossRef]
- Saïd, I.; Tauban, D.; Renou, M.; Mori, K.; Rochat, D. Structure and Function of the Antennal Sensilla of the Palm Weevil Rhynchophorus palmarum (Coleoptera, Curculionidae). J. Insect Physiol. 2003, 49, 857–872. [Google Scholar] [CrossRef]
- Jourdan, H.; Barbier, R.; Bernard, J.; Ferran, A. Antennal Sensilla and Sexual Dimorphism of the Adult Ladybird Beetle Semiadalia undecimnotata Schn. (Coleoptera: Coccinellidae). Int. J. Insect Morphol. Embryol. 1995, 24, 307–322. [Google Scholar] [CrossRef]
- Marques-Silva, S.; Matiello-Guss, C.P.; Delabie, J.; Mariano, C.; Serrão, J.E. Sensilla and Secretory Glands in the Antennae of a Primitive Ant: Dinoponera lucida (Formicidae: Ponerinae). Microsc. Res. Tech. 2006, 69, 885–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weis, A.; Schönitzer, K.; Melzer, R.R. Exocrine Glands in the Antennae of the Carabid Beetle, Platynus assimilis (Paykull) 1790 (Coleoptera, Carabidae, Pterostichinae). Int. J. Insect Morphol. Embryol. 1999, 28, 331–335. [Google Scholar] [CrossRef]
- Medvedev, L.N.; Pavlov, S.I. Mating Behavior of the Chrysomelidae (Coleoptera). Entomol. Rev. 1988, 67, 100–109. [Google Scholar]
- Peng, L.F.; Chen, L.M.; Li, J.L.; Hou, Y.M.; Chen, Y.G. Mate recognition and antennal morphology of Octodonta nipae (Coleoptera: Chrysomelidae) adults. J. Asia-Pac. Entomol. 2018, 21, 268–278. [Google Scholar] [CrossRef]
- Guo, X.J.; Yu, Q.Q.; Chen, D.F.; Wei, J.N.; Yang, P.C.; Yu, J.; Wang, X.H.; Kang, L. 4-Vinylanisole Is an Aggregation Pheromone in Locusts. Nature 2020, 584, 584–588. [Google Scholar] [CrossRef]
- Shi, X.; Zhang, S.F.; Liu, F.; Zhang, Z.; Xu, F.Y.; Yin, S.Y.; Kong, X.B. Sensilla on Antennae and Mouthparts of Adult Spruce Bark Beetle Ips typographus (Coleoptera: Curculionidae). Microsc. Res. Tech. 2021, 84, 1484–1497. [Google Scholar] [CrossRef] [PubMed]
- Altner, H.; Prillinger, L. Ultrastructure of Invertebrate Chemo-, Thermo-, and Hygroreceptors and Its Functional Significance. Int. Rev. Cytol. 1980, 67, 69–139. [Google Scholar]
- Bartlet, E.; Romani, R.; Williams, I.H.; Isidoro, N. Functional Anatomy of Sensory Structures on the Antennae of Psylliodes chrysocephala L. (Coleoptera: Chrysomelidae). Int. J. Insect Morphol. Embryol. 1999, 28, 291–300. [Google Scholar] [CrossRef]
- Tominaga, Y.; Yokohari., F. External Structure of the Sensillum Capitulum, A Hygro- And Thermoreceptive Sensillum of the Cockroach, Periplaneta americana. Cell Tissue Res. 1982, 226, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Altner, H.; Sass, H.; Altner, I. Relationship Between Structure and Function of Antennal Chemo-, Hygro-, and Thermoreceptive Sensilla in Periplaneta americana. Cell Tissue Res. 1977, 176, 389–405. [Google Scholar] [CrossRef] [PubMed]
- Onagbola, E.O.; Fadamiro, H.Y. Scanning Electron Microscopy Studies of Antennal Sensilla of Pteromalus Cerealellae (Hymenoptera: Pteromalidae). Micron 2008, 39, 526–535. [Google Scholar] [CrossRef]
- Wang, H.M.; Zheng, H.X.; Zhang, Y.W.; Zhang, X.H. Morphology and Distribution of Antennal, Maxillary Palp and Labial Palp Sensilla of the Adult Bruchid Beetles, Callosobruchus chinensis (L.) (Coleoptera: Bruchidae). Entomol. Res. 2018, 48, 466–479. [Google Scholar] [CrossRef]
- Chi, D.F.; Wang, G.L.; Liu, J.W.; Wu, Q.Y.; Zhu, Y.P. Antennal Morphology and Sensilla of Asian Multicolored Ladybird Beetles, Harmonia axyridis Pallas (Coleoptera: Coccinellidae). Entomol. News 2009, 120, 137–152. [Google Scholar] [CrossRef]
- Picimbon, J.F.; Dietrich, K.; Krieger, J.; Breer, H. Identity and Expression Pattern of Chemosensory Proteins in Heliothis virescens (Lepidoptera. Noctuidae. Insect Biochem. Mol. Biol. 2001, 31, 1173–1181. [Google Scholar] [CrossRef]
- Pelletier, J.; Leal, W.S. Characterization of Olfactory Genes in the Antennae of the Southern House Mosquito, Culex quinquefasciatus. J. Insect Physiol. 2011, 57, 915–929. [Google Scholar] [CrossRef]
- Kitabayashi, A.N.; Arai, T.; Kubo, T.; Natori, S. Molecular Cloning of cDNA for p10, A Novel Protein That Increases in the Regenerating Legs of Periplaneta americana (American cockroach). Insect Biochem. Mol. Biol. 1998, 28, 785–790. [Google Scholar] [CrossRef]
- Ban, L.; Scaloni, A.; Brandazza, A.; Angeli, S.; Zhang, L.; Yan, Y.; Pelosi, P. Chemosensory Proteins of Locusta migratoria. Insect Mol. Biol. 2003, 12, 125–134. [Google Scholar] [CrossRef]
- Zeng, Y.; Merchant, A.; Wu, Q.J.; Wang, S.L.; Kong, L.; Zhou, X.G.; Xie, W.; Zhang, Y.J. A Chemosensory Protein BtabCSP11 Mediates Reproduction in Bemisia tabaci. Front. Physiol. 2020, 11, 709. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence-F (5′-3′) | Primer Sequence-R (5′-3′) |
|---|---|---|
| Clone primers | ||
| CSP1 | ATGGTACCGTTAATCTGTG | CTATGTTCTGTTAAACCGGA |
| CSP2 | ATGAATCGTTGTGGTTTGTC | TTAAATGGAGGAGTCGTTGA |
| CSP3 | ATGAATAGTTTTTATTTATCG | TTAAATGCTCGTATCTTTAAG |
| CSP4 | ATGGTGTCGTTCCTATTAGT | CTAAGCTTTGGTAATGGGCT |
| CSP5 | ATGAAAGTAACTATTGCAAT | TTACAAAACAATTCCTTCTT |
| CSP6 | ATGGAAATGTGTTTTATTTTG | TTATAATTCGGGAAATCCTT |
| CSP7 | ATGAAGCCGATTTTTCTGGTG | TTATAAAACAATTCCTTCT |
| CSP8 | ATGAACAAATCAGTATTGTTTG | TTACAATGTGTTTTTGTATTC |
| CSP9 | ATGGGGTTTGCAAGATTAA | TTAAGGACTGTTTAAGAATG |
| CSP10 | ATGAAGACGTTTGTCGTTTG | TTATGTACTGATTTCTTTTTCA |
| CSP11 | ATGCAAACATTTCAGCCT | TTAACCGGAATACTGTTT |
| RT-qPCR primers | ||
| CSP1-q | CCTGATGATATATACGTGA | TTGTACTATTTGTGGAAGC |
| CSP2-q | TTGTGGTTTGTCTGTCTT | ACGTAGCTCTTGATGATT |
| CSP3-q | TGGCGCTCCGAAAACCTT | ATCGCAATCACCACAACCT |
| CSP4-q | ATGGTGTCGTTCCTATTAGTTTT | TTTCTTCGAGGTTGATGTTGTC |
| CSP5-q | GGTTACATCGACTGCCTT | TTTTTGCTCCTTTTTTCT |
| CSP6-q | CCTAAAAAAAGTCATCCCT | TATAATTTCTCGTCCAGCT |
| CSP7-q | GAAAAACAAAGAACAGGA | TATTTAGCAGACAACTCG |
| CSP8-q | CAGACACCTGTACCAAAG | TCATCAAATAACCAATCA |
| CSP9-q | TAGTAAGCGATTATTGGA | TATGGATAGCCTCAGGTA |
| CSP10-q | AAGTCCTTCCTGACGCTC | AGTTCCCCATACCAATCG |
| CSP11-q | ACTCCGCCAACTAAAATG | ACTGACTGCACGAACCCT |
| RPL19 | AAGGAAGGCATTGTGGAT | GACGCAAATCTCGCATAC |
| Antennal Segment | Length (μm) | |
|---|---|---|
| Male | Female | |
| Scape | 283.4 ± 12.96 | 273.6 ± 10.17 |
| Pedicel | 158.8 ± 8.31 | 179.2 ± 7.51 |
| Flagellomere 1 | 273 ± 4.93 | 294.6 ± 2.82 |
| Flagellomere 2 | 230 ± 5.43 | 229.4 ± 8.66 |
| Flagellomere 3 | 184.4 ± 2.79 | 190.8 ± 2.80 |
| Flagellomere 4 | 202.6 ± 4.75 | 215.2 ± 9.16 |
| Flagellomere 5 | 194.6 ± 7.31 | 198.4 ± 4.41 |
| Flagellomere 6 | 186.6 ± 11.57 | 173.0 ± 5.02 |
| Flagellomere 7 | 183.4 ± 7.05 | 183.8 ± 7.59 |
| Flagellomere 8 | 186.8 ± 3.51 | 178 ± 8.93 |
| Flagellomere 9 | 276.2 ± 9.84 | 265.6 ± 6.49 |
| Total length | 2359.8 ± 42.87 | 2381.6 ± 40.57 |
| Sensillum Type | Length (μm) | Width (μm) | Density | ||
|---|---|---|---|---|---|
| Male | Female | Male | Female | ||
| Sensilla trichodea 1 | 43.45 ± 0.88 | 42.63 ± 1.09 | 2.35 ± 0.09 | 2.50 ± 0.08 | ++++ |
| Sensilla trichodea 2 | 22.28 ± 0.56 | 22.65 ± 0.48 | 1.88 ± 0.04 | 2.11 ± 0.04 | +++ |
| Sensilla chaetica | 47.07 ± 1.48 | 48.47 ± 1.85 | 2.74 ± 0.08 | 3.02 ± 0.07 | +++ |
| Sensilla basiconica 1 | 11.83 ± 0.21 | 13.27 ± 1.40 | 1.45 ± 0.03 | 1.60 ± 0.03 | ++ |
| Sensilla basiconica 2 | 10.85 ± 0.28 | 10.70 ± 0.27 | 1.81 ± 0.06 | 1.80 ± 0.06 | ++ |
| Sensilla coeloconica | 5.03 ± 0.32 | 4.99 ± 0.32 | 1.28 ± 0.03 | 1.24 ± 0.03 | + |
| Sensilla styloconica | 3.62 ± 0.19 | 4.08 ± 0.23 | 1.13 ± 0.04 | 1.12 ± 0.05 | + |
| Böhm bristle | 7.16 ± 0.43 | 6.46 ± 0.34 | 1.30 ± 0.05 | 1.10 ± 0.09 | ++ |
| CSP1 | CSP2 | CSP3 | CSP4 | CSP5 | CSP6 | CSP7 | CSP8 | CSP9 | CSP10 | CSP11 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CSP1 | 10.69% | 10.73% | 15.33% | 15.33% | 15.71% | 15.33% | 11.11% | 11.11% | 16.86% | 6.04% | |
| CSP2 | 10.69% | 48.18% | 21.43% | 21.90% | 18.98% | 23.36% | 10.95% | 17.52% | 21.17% | 5.88% | |
| CSP3 | 10.73% | 48.18% | 18.12% | 26.67% | 16.30% | 23.70% | 11.85% | 22.22% | 29.63% | 5.38% | |
| CSP4 | 15.33% | 21.43% | 18.12% | 40.44% | 38.81% | 43.28% | 31.85% | 30.15% | 43.28% | 11.43% | |
| CSP5 | 15.33% | 21.90% | 26.67% | 40.44% | 32.58% | 41.22% | 30.08% | 25.56% | 44.27% | 17.52% | |
| CSP6 | 15.71% | 18.98% | 16.30% | 38.81% | 32.58% | 39.53% | 21.71% | 30.53% | 42.64% | 17.39% | |
| CSP7 | 15.33% | 23.36% | 23.70% | 43.28% | 41.22% | 39.53% | 19.38% | 31.54% | 42.19% | 7.69% | |
| CSP8 | 11.11% | 10.95% | 11.85% | 31.85% | 30.08% | 21.71% | 19.38% | 15.50% | 29.69% | 12.32% | |
| CSP9 | 11.11% | 17.52% | 22.22% | 30.15% | 25.56% | 30.53% | 31.54% | 15.50% | 37.90% | 11.11% | |
| CSP10 | 16.86% | 21.17% | 29.63% | 43.28% | 44.27% | 42.64% | 42.19% | 29.69% | 37.90% | 7.01% | |
| CSP11 | 6.04% | 5.88% | 5.38% | 11.43% | 17.52% | 17.39% | 7.69% | 12.32% | 11.11% | 7.01% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, C.; Yue, Y.; Zhang, Y.; Tian, Z.-Y.; Chen, H.-S.; Guo, J.-Y.; Zhou, Z.-S. Scanning Electron Microscopic Analysis of Antennal Sensilla and Tissue-Expression Profiles of Chemosensory Protein Genes in Ophraella communa (Coleoptera: Chrysomelidae). Insects 2022, 13, 183. https://doi.org/10.3390/insects13020183
Ma C, Yue Y, Zhang Y, Tian Z-Y, Chen H-S, Guo J-Y, Zhou Z-S. Scanning Electron Microscopic Analysis of Antennal Sensilla and Tissue-Expression Profiles of Chemosensory Protein Genes in Ophraella communa (Coleoptera: Chrysomelidae). Insects. 2022; 13(2):183. https://doi.org/10.3390/insects13020183
Chicago/Turabian StyleMa, Chao, Yang Yue, Yan Zhang, Zhen-Ya Tian, Hong-Song Chen, Jian-Ying Guo, and Zhong-Shi Zhou. 2022. "Scanning Electron Microscopic Analysis of Antennal Sensilla and Tissue-Expression Profiles of Chemosensory Protein Genes in Ophraella communa (Coleoptera: Chrysomelidae)" Insects 13, no. 2: 183. https://doi.org/10.3390/insects13020183