
Marked Effects of Larval Salt Exposure on the Life History and Gut Microbiota of the Malaria Vector Anopheles merus (Diptera: Culicidae)

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Strain and Larval Treatment
2.2. The Effects of Variable Salt Concentration on Life History
2.2.1. Development Time
2.2.2. Adult Longevity
2.2.3. Fertility and Fecundity
2.2.4. Insecticide Tolerance
2.3. The Effect of Larval Salt Concentration on the Expression of Defensin-1
2.3.1. RNA Extraction and Quality Control
2.3.2. Analysis of Transcript Levels
2.4. The Effect of Larval Exposure to Variable Salt Concentration on An. merus Gut Microbiota
2.4.1. Midgut Resection and DNA Extraction
2.4.2. Amplification of the V3–V4 Hypervariable Region of the 16S rRNA Gene
2.4.3. Bioinformatic Analysis
3. Results
3.1. The Effect of Larval Salt Concentration on Life History
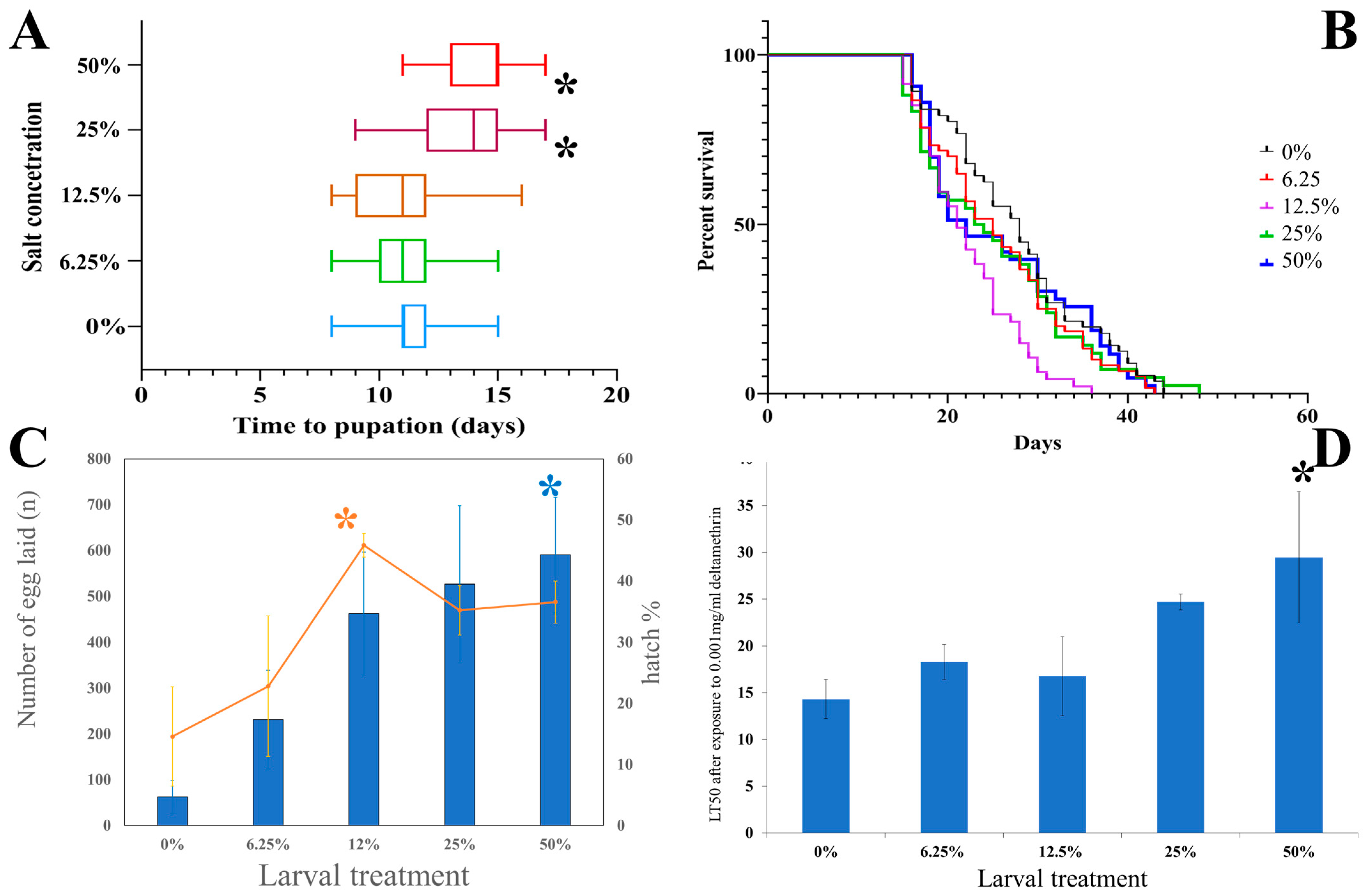
3.1.1. Development Time
3.1.2. Adult Longevity
3.1.3. Fertility and Fecundity
3.1.4. Insecticide Tolerance
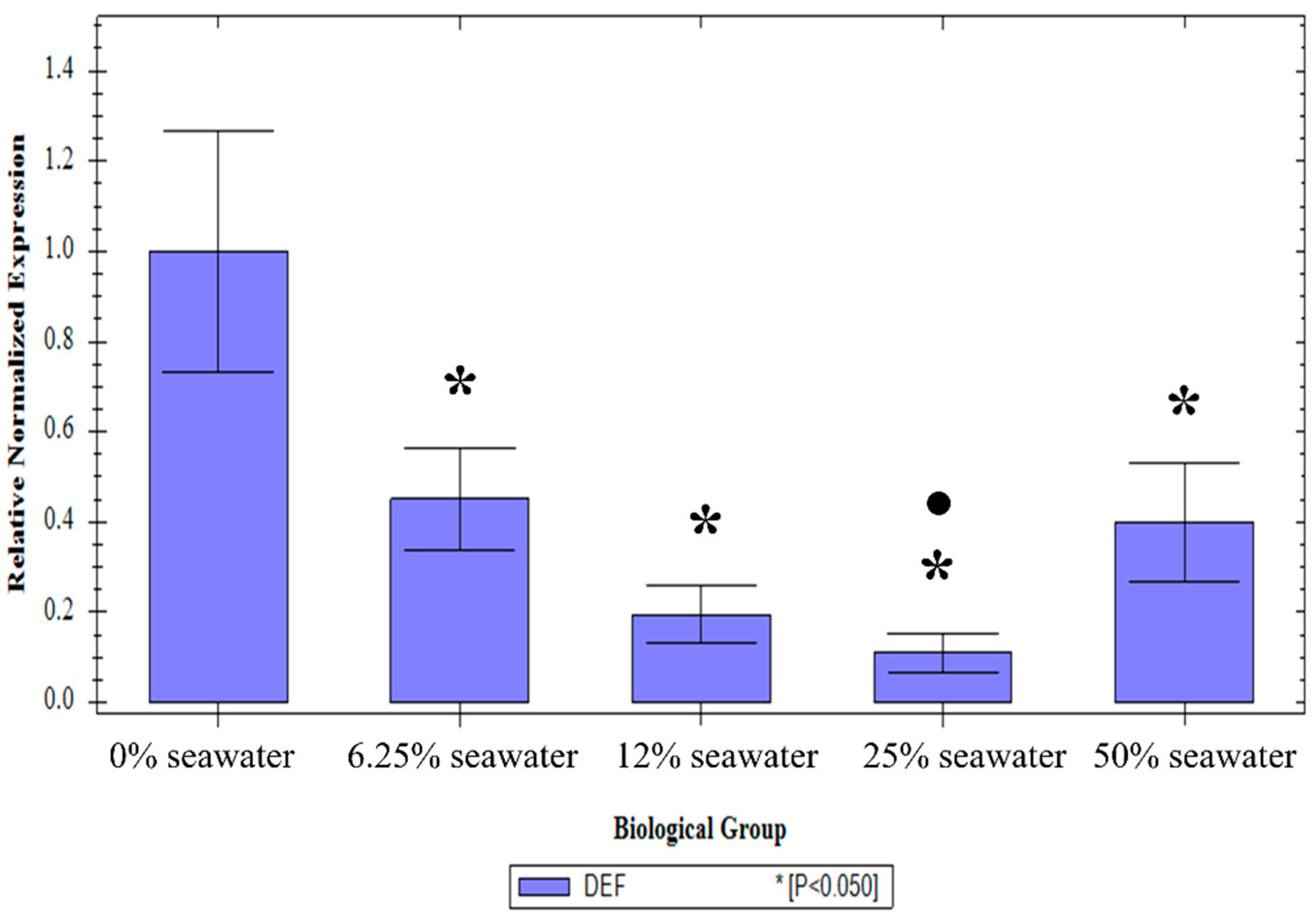
3.2. The Effect of the Larval Salt Concentration on Defensin-1 Expression
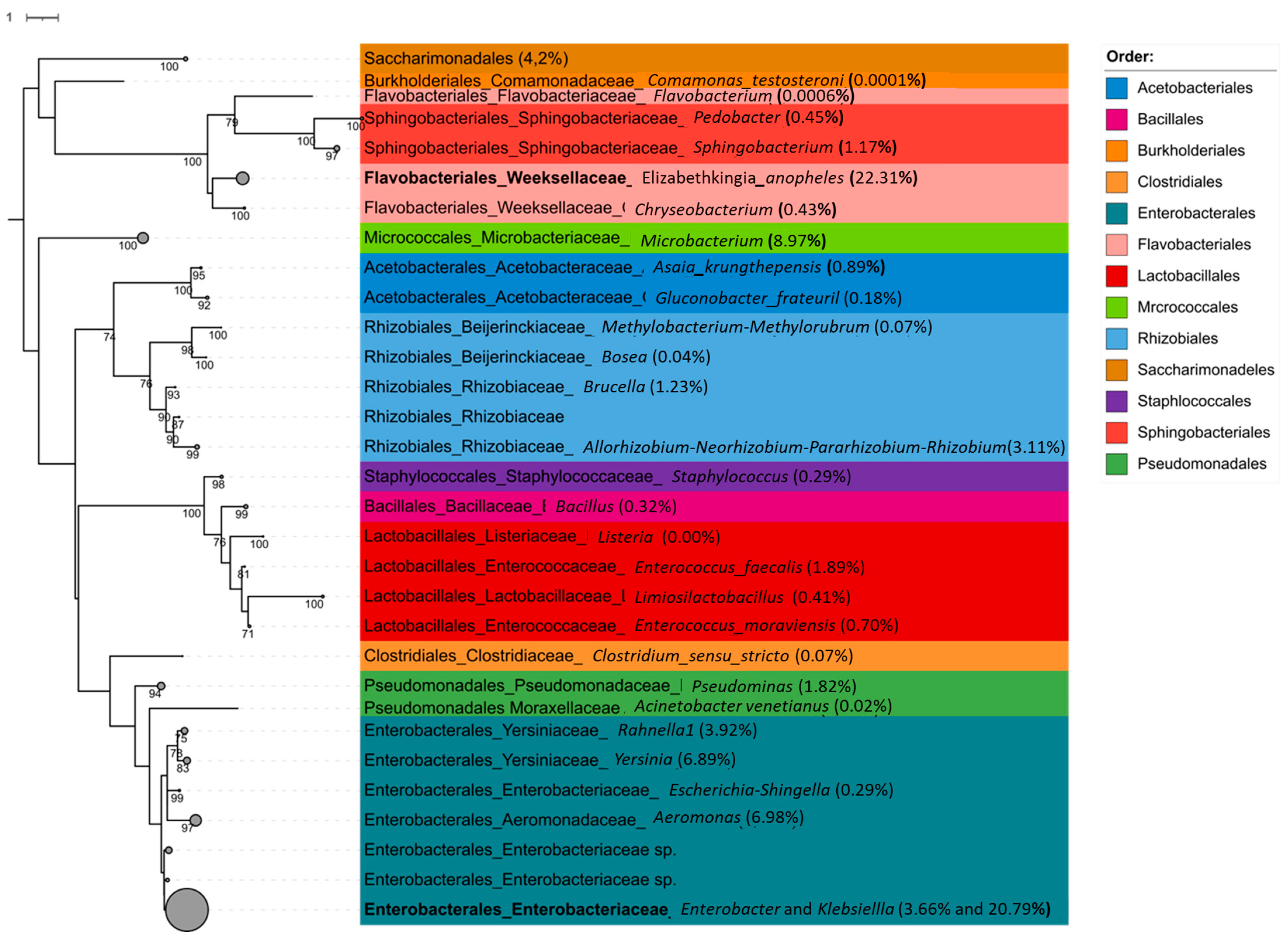
3.3. The Effect of Larval Salt Concentration on An. merus Microbiota
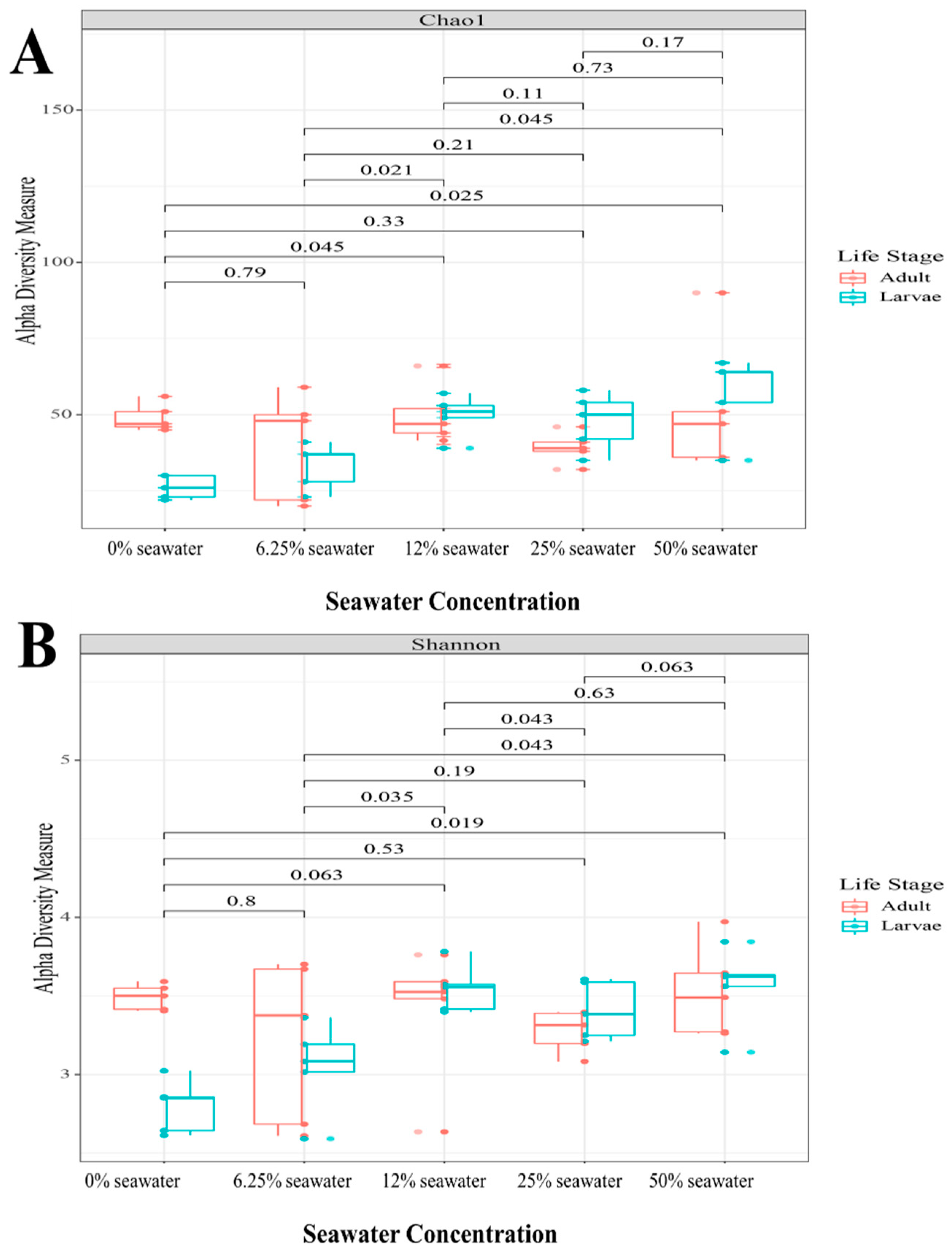
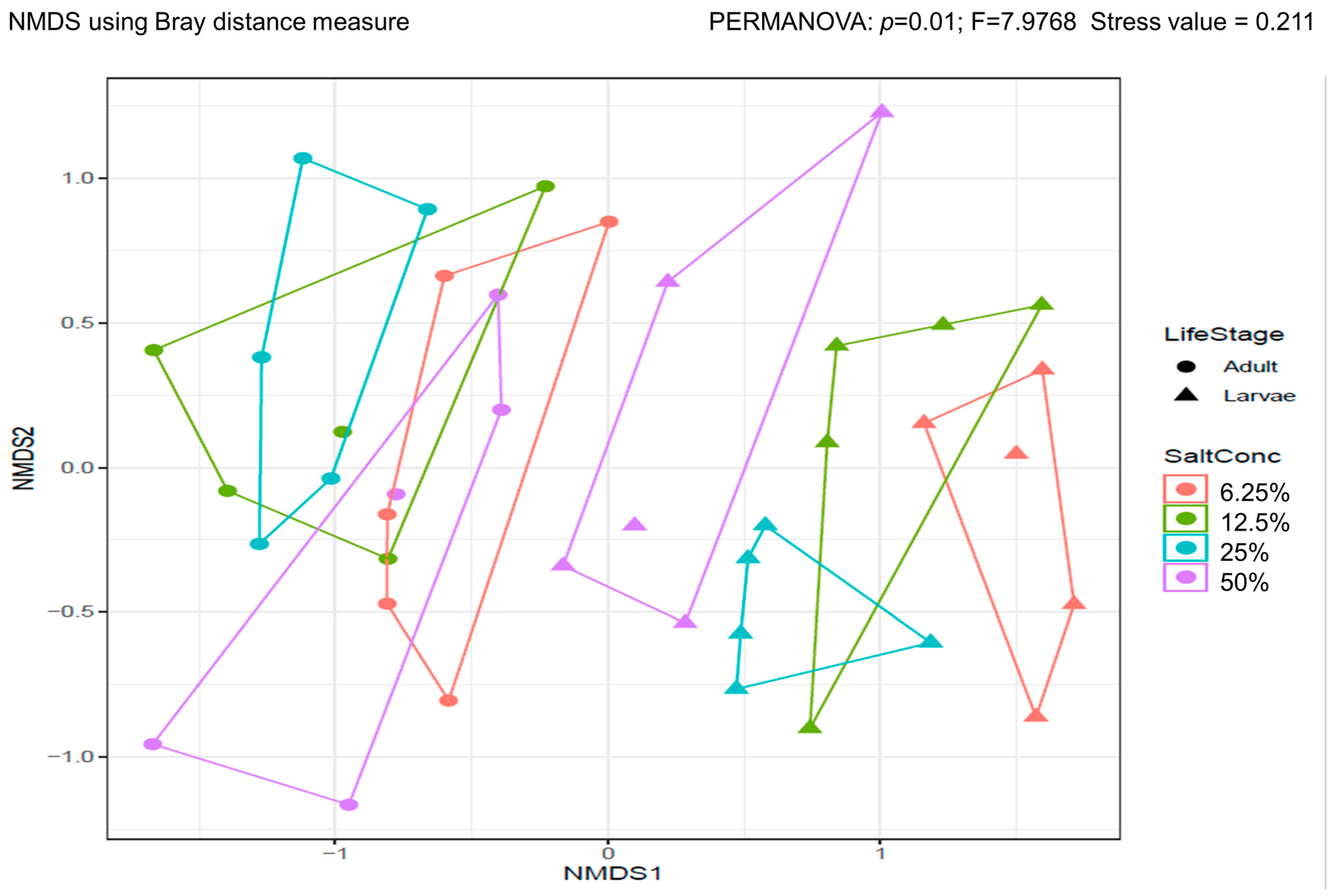
3.3.1. Alpha and Beta Diversity
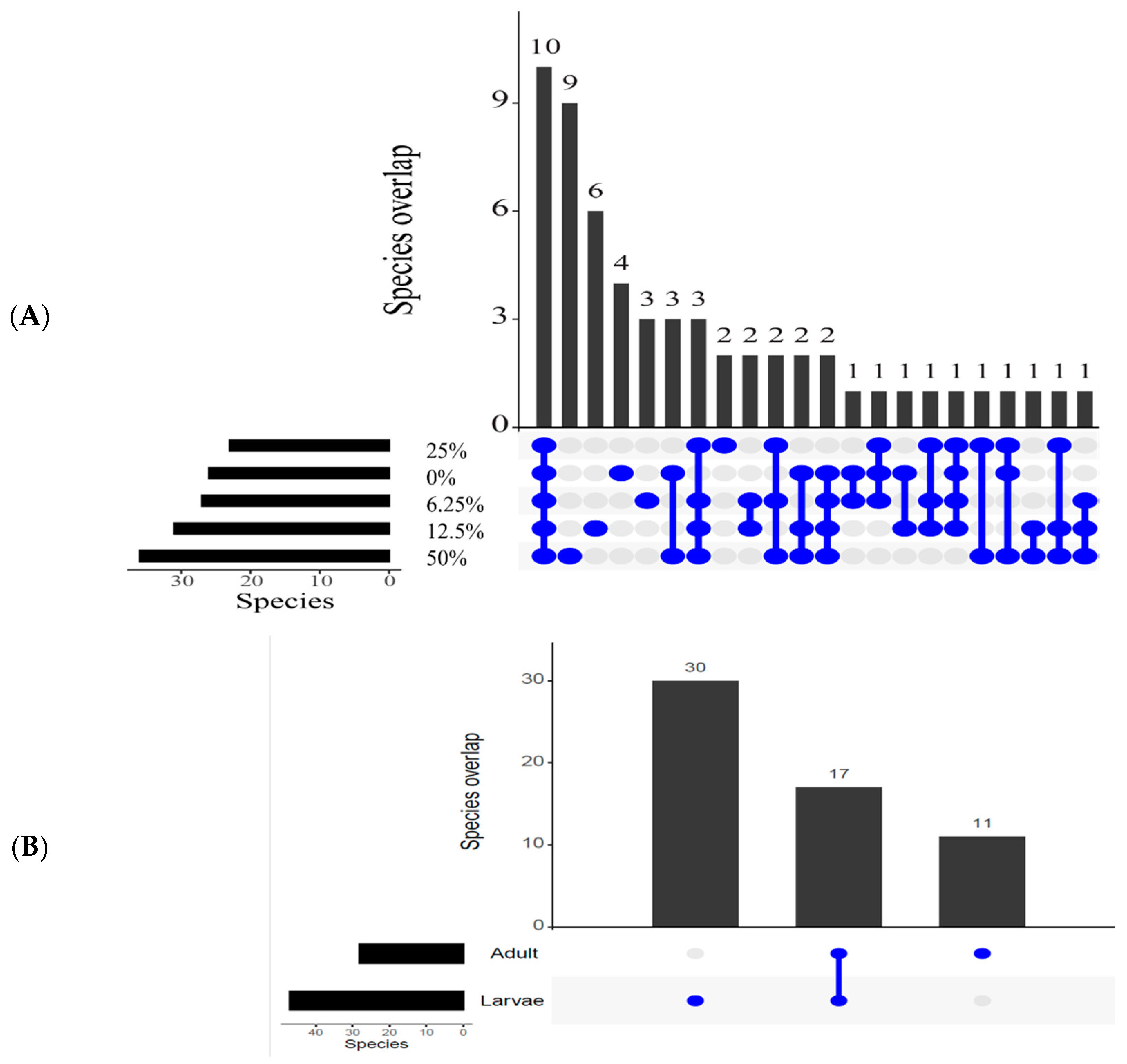
3.3.2. Overlapping Species
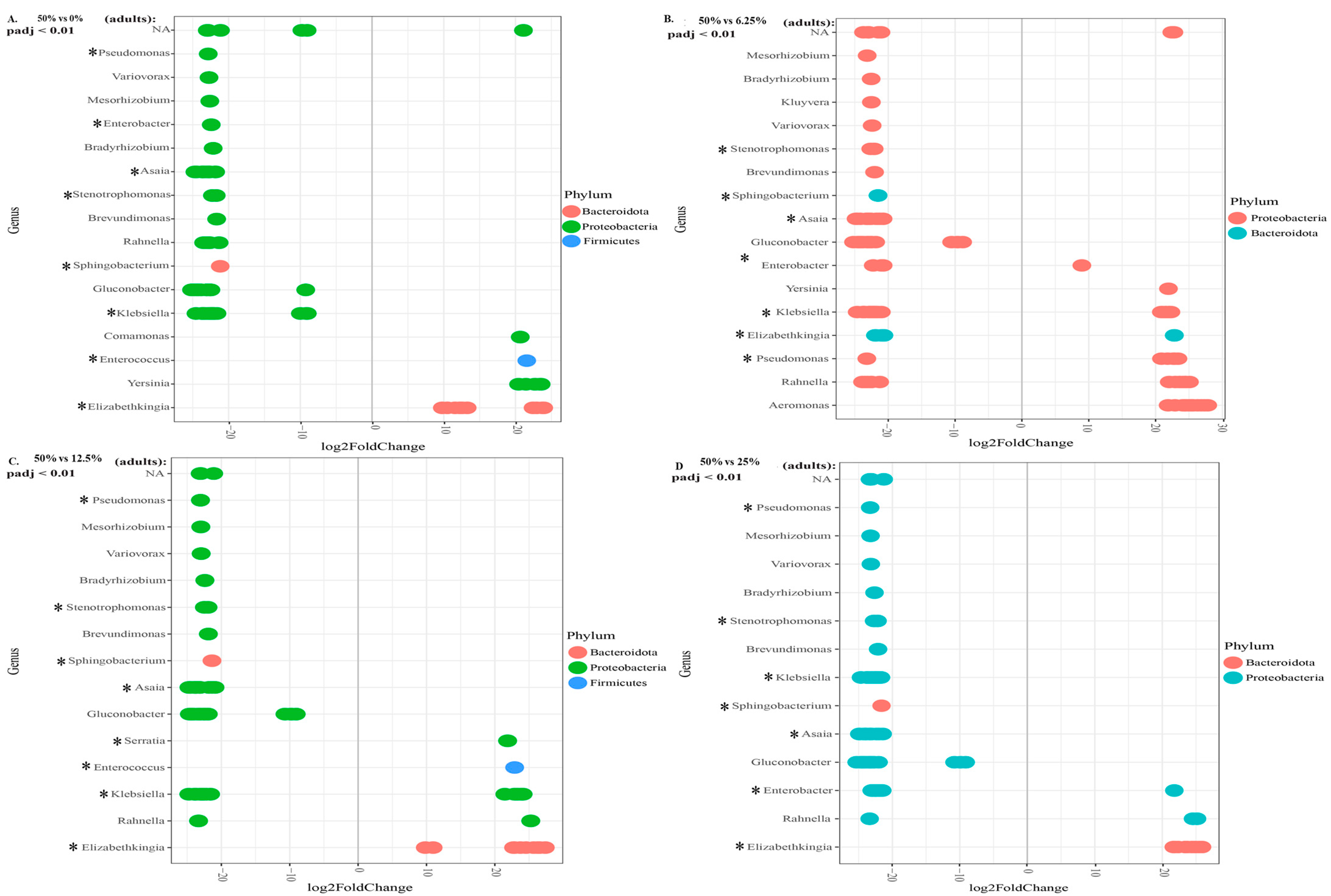
3.3.3. Relative Abundance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dobson, T. Seaweed flies (Diptera: Coleopidae). In Marine Insects; Cheng, L., Ed.; North Holland Publishers: Amsterdam, The Netherlands, 1976. [Google Scholar]
- Bradley, T.J. Physiology of osmoregulation in mosquitoes. Annu. Rev. Entomol. 1987, 32, 439–462. [Google Scholar] [CrossRef] [PubMed]
- O’Meara, G.F. Saltmarsh mosquitoes (Diptera: Culicidae). In Marine Insects; Cheng, L., Ed.; North Holland Publishing Company: Amsterdam, The Netherlands, 1976; pp. 300–330. [Google Scholar]
- Bradley, T.J. Saline-water insects: Ecology, physiology and evolution. In Aquatic Insects: Challenges to Populations; Lancaster, J., Briers, R.A., Eds.; CAB International: Wallingford, UK, 2008; pp. 20–35. [Google Scholar]
- Kengne, P.; Charmantier, G.; Blondeau-Bidet, E.; Costantini, C.; Ayala, D. Tolerance of disease-vector mosquitoes to brackish water and their osmoregulatory ability. Ecosphere 2019, 10, e02783. [Google Scholar] [CrossRef] [Green Version]
- Blagrove, M.S.; Sherlock, K.; Chapman, G.E.; Impoinvil, D.E.; McCall, P.J.; Medlock, J.M.; Lycett, G.; Solomon, T.; Baylis, M. Evaluation of the vector competence of a native UK mosquito Ochlerotatus detritus (Aedes detritus) for dengue, chikungunya and West Nile viruses. Parasites Vectors 2016, 9, 452. [Google Scholar] [CrossRef] [Green Version]
- Ramasamy, R.; Surendran, S.N. Global climate change and its potential impact on disease transmission by salinity-tolerant mosquito vectors in coastal zones. Front. Physiol. 2012, 3, 198. [Google Scholar] [CrossRef] [Green Version]
- Surendran, S.N.; Jude, P.J.; Ramasamy, R. Variations in salinity tolerance of malaria vectors of the Anopheles subpictus complex in Sri Lanka and the implications for malaria transmission. Parasites Vectors 2011, 4, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ould Lemrabott, M.A.; Le Goff, G.; Kengne, P.; Ndiaye, O.; Costantini, C.; Mint Lekweiry, K.; Ould Ahmedou Salem, M.S.; Robert, V.; Basco, L.; Simard, F.; et al. First report of Anopheles (Cellia) multicolor during a study of tolerance to salinity of Anopheles arabiensis larvae in Nouakchott, Mauritania. Parasites Vectors 2020, 13, 522. [Google Scholar] [CrossRef]
- Sinka, M.E.; Bangs, M.J.; Manguin, S.; Coetzee, M.; Mbogo, C.M.; Hemingway, J.; Patil, A.P.; Temperley, W.H.; Gething, P.W.; Kabaria, C.W.; et al. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: Occurrence data, distribution maps and bionomic précis. Parasites Vectors 2010, 3, 117. [Google Scholar] [CrossRef] [Green Version]
- Bartilol, B.; Omedo, I.; Mbogo, C.; Mwangangi, J.; Rono, M.K. Bionomics and ecology of Anopheles merus along the East and Southern Africa coast. Parasites Vectors 2021, 14, 84. [Google Scholar] [CrossRef]
- Braack, L.; Bornman, R.; Kruger, T.; Dahan-Moss, Y.; Gilbert, A.; Kaiser, M.; Oliver, S.V.; Cornel, A.J.; Lee, Y.; Norris, D.E.; et al. Malaria Vectors and Vector Surveillance in Limpopo Province (South Africa): 1927 to 2018. Int. J. Environ. Res. Public Health 2020, 17, 4125. [Google Scholar] [CrossRef]
- Mbokazi, F.; Coetzee, M.; Brooke, B.; Govere, J.; Reid, A.; Owiti, P.; Kosgei, R.; Zhou, S.; Magagula, R.; Kok, G.; et al. Changing distribution and abundance of the malaria vector Anopheles merus in Mpumalanga Province, South Africa. Public Health Action 2018, 8, S39–S43. [Google Scholar] [CrossRef]
- White, B.J.; Kundert, P.N.; Turissini, D.A.; Van Ekeris, L.; Linser, P.J.; Besansky, N.J. Dose and developmental responses of Anopheles merus larvae to salinity. J. Exp. Biol. 2013, 216, 3433–3441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uyhelji, H.A.; Cheng, C.; Besansky, N.J. Transcriptomic differences between euryhaline and stenohaline malaria vector sibling species in response to salinity stress. Mol. Ecol. 2016, 25, 2210–2225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vizioli, J.; Richman, A.M.; Uttenweiler-Joseph, S.; Blass, C.; Bulet, P. The defensin peptide of the malaria vector mosquito Anopheles gambiae: Antimicrobial activities and expression in adult mosquitoes. Insect Biochem. Mol. Biol. 2001, 31, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Richman, A.M.; Dimopoulos, G.; Seeley, D.; Kafatos, F.C. Plasmodium activates the innate immune response of Anopheles gambiae mosquitoes. EMBO J. 1997, 16, 6114–6119. [Google Scholar] [CrossRef] [Green Version]
- Kido, E.A.; Pandolfi, V.; Houllou-Kido, L.M.; Andrade, P.P.; Marcelino, F.C.; Nepomuceno, A.L.; Abdelnoor, R.V.; Burnquist, W.L.; Benko-Iseppon, A.M. Plant antimicrobial peptides: An overview of SuperSAGE transcriptional profile and a functional review. Curr. Protein Pept. Sci. 2010, 11, 220–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreaux, A.M.G.; Stone, C.M.; Barreaux, P.; Koella, J.C. The relationship between size and longevity of the malaria vector Anopheles gambiae (s.s.) depends on the larval environment. Parasites Vectors 2018, 11, 485. [Google Scholar] [CrossRef] [Green Version]
- Jannat, K.N.; Roitberg, B.D. Effects of larval density and feeding rates on larval life history traits in Anopheles gambiae s.s. (Diptera: Culicidae). J. Vector Ecol. J. Soc. Vector Ecol. 2013, 38, 120–126. [Google Scholar] [CrossRef]
- Moller-Jacobs, L.L.; Murdock, C.C.; Thomas, M.B. Capacity of mosquitoes to transmit malaria depends on larval environment. Parasites Vectors 2014, 7, 593. [Google Scholar] [CrossRef]
- Okech, B.A.; Gouagna, L.C.; Yan, G.; Githure, J.I.; Beier, J.C. Larval habitats of Anopheles gambiae s.s. (Diptera: Culicidae) influences vector competence to Plasmodium falciparum parasites. Malar. J. 2007, 6, 50. [Google Scholar] [CrossRef] [Green Version]
- Jeanrenaud, A.; Brooke, B.D.; Oliver, S.V. The effects of larval organic fertiliser exposure on the larval development, adult longevity and insecticide tolerance of zoophilic members of the Anopheles gambiae complex (Diptera: Culicidae). PLoS ONE 2019, 14, e0215552. [Google Scholar] [CrossRef]
- Oliver, S.V.; Brooke, B.D. The effect of metal pollution on the life history and insecticide resistance phenotype of the major malaria vector Anopheles arabiensis (Diptera: Culicidae). PLoS ONE 2018, 13, e0192551. [Google Scholar] [CrossRef] [Green Version]
- Oliver, S.V.; Brooke, B.D. The effect of commercial herbicide exposure on the life history and insecticide resistance phenotypes of the major malaria vector Anopheles arabiensis (Diptera: Culicidae). Acta Trop. 2018, 188, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Coon, K.L.; Valzania, L.; Brown, M.R.; Strand, M.R. Predaceous Toxorhynchites mosquitoes require a living gut microbiota to develop. Proc. Biol. Sci. 2020, 287, 20192705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coon, K.L.; Vogel, K.J.; Brown, M.R.; Strand, M.R. Mosquitoes rely on their gut microbiota for development. Mol. Ecol. 2014, 23, 2727–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coon, K.L.; Brown, M.R.; Strand, M.R. Gut bacteria differentially affect egg production in the anautogenous mosquito Aedes aegypti and facultatively autogenous mosquito Aedes atropalpus (Diptera: Culicidae). Parasites Vectors 2016, 9, 375. [Google Scholar] [CrossRef] [Green Version]
- Coon, K.L.; Hegde, S.; Hughes, G.L. Interspecies microbiome transplantation recapitulates microbial acquisition in mosquitoes. Microbiome 2022, 10, 58. [Google Scholar] [CrossRef]
- Bai, L.; Wang, L.; Vega-Rodríguez, J.; Wang, G.; Wang, S. A Gut Symbiotic Bacterium Serratia marcescens Renders Mosquito Resistance to Plasmodium Infection through Activation of Mosquito Immune Responses. Front. Microbiol. 2019, 10, 1580. [Google Scholar] [CrossRef] [Green Version]
- Boissière, A.; Tchioffo, M.T.; Bachar, D.; Abate, L.; Marie, A.; Nsango, S.E.; Shahbazkia, H.R.; Awono-Ambene, P.H.; Levashina, E.A.; Christen, R.; et al. Midgut microbiota of the malaria mosquito vector Anopheles gambiae and interactions with Plasmodium falciparum infection. PLoS Pathog. 2012, 8, e1002742. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, B.; Portugal, S.; Tran, T.M.; Gozzelino, R.; Ramos, S.; Gomes, J.; Regalado, A.; Cowan, P.J.; d’Apice, A.J.; Chong, A.S.; et al. Gut microbiota elicits a protective immune response against malaria transmission. Cell 2014, 159, 1277–1289. [Google Scholar] [CrossRef] [Green Version]
- Tchioffo, M.T.; Boissière, A.; Churcher, T.S.; Abate, L.; Gimonneau, G.; Nsango, S.E.; Awono-Ambéné, P.H.; Christen, R.; Berry, A.; Morlais, I. Modulation of malaria infection in Anopheles gambiae mosquitoes exposed to natural midgut bacteria. PLoS ONE 2013, 8, e81663. [Google Scholar] [CrossRef]
- Alfano, N.; Tagliapietra, V.; Rosso, F.; Manica, M.; Arnoldi, D.; Pindo, M.; Rizzoli, A. Changes in Microbiota Across Developmental Stages of Aedes koreicus, an Invasive Mosquito Vector in Europe: Indications for Microbiota-Based Control Strategies. Front. Microbiol. 2019, 10, 2832. [Google Scholar] [CrossRef] [PubMed]
- Dinparast Djadid, N.; Jazayeri, H.; Raz, A.; Favia, G.; Ricci, I.; Zakeri, S. Identification of the midgut microbiota of An. stephensi and An. maculipennis for their application as a paratransgenic tool against malaria. PLoS ONE 2011, 6, e28484. [Google Scholar] [CrossRef] [PubMed]
- Duguma, D.; Hall, M.W.; Smartt, C.T.; Neufeld, J.D. Effects of Organic Amendments on Microbiota Associated with the Culex nigripalpus Mosquito Vector of the Saint Louis Encephalitis and West Nile Viruses. Msphere 2017, 2, e00387-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krajacich, B.J.; Huestis, D.L.; Dao, A.; Yaro, A.S.; Diallo, M.; Krishna, A.; Xu, J.; Lehmann, T. Investigation of the seasonal microbiome of Anopheles coluzzii mosquitoes in Mali. PLoS ONE 2018, 13, e0194899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louie, W.; Coffey, L.L. Microbial Composition in Larval Water Enhances Aedes aegypti Development but Reduces Transmissibility of Zika Virus. Msphere 2021, 6, e0068721. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhou, S.; Wu, J.; Zhang, X.; Wang, Y.; Li, Z.; Chen, X.G.; Zhou, X. An Experimental Evaluation of Toxicity Effects of Sodium Chloride on Oviposition, Hatching and Larval Development of Aedes albopictus. Pathogens 2022, 11, 262. [Google Scholar] [CrossRef]
- Hunt, R.H.; Brooke, B.D.; Pillay, C.; Koekemoer, L.L.; Coetzee, M. Laboratory selection for and characteristics of pyrethroid resistance in the malaria vector Anopheles funestus. Med. Vet. Entomol. 2005, 19, 271–275. [Google Scholar] [CrossRef]
- Kaplan, E.L.; Meier, P. Nonparametric Estimation from Incomplete Observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Harrington, D.P.; Fleming, T.R. A class of rank test procedures for censored survival data. Biometrika 1982, 69, 553–566. [Google Scholar] [CrossRef]
- Samuel, M.; Brooke, B.D.; Oliver, S.V. Effects of inorganic fertilizer on larval development, adult longevity and insecticide susceptibility in the malaria vector Anopheles arabiensis (Diptera: Culicidae). Pest Manag. Sci. 2020, 76, 1560–1568. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis, 2nd ed.; Cambridge University Press: New York, NY, USA, 1952. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An Analysis of Variance Test for Normality (Complete Samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Tukey, J.W. Comparing individual means in the analysis of variance. Biometrics 1949, 5, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef] [Green Version]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Andersen, K.S.; Kirkegaard, R.H.; Karst, S.M.; Albertsen, M.J.B. ampvis2: An R package to analyse and visualise 16S rRNA amplicon data. BioRxiv 2018, 299537. [Google Scholar]
- Conway, J.R.; Lex, A.; Gehlenborg, N. UpSetR: An R package for the visualization of intersecting sets and their properties. Bioinformatics 2017, 33, 2938–2940. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Albuquerque, F.M.; Freitas, E.C.B.F.; do Nascimento, D.V.; de Almeida, W.A.; Nova, I.C.V.; Paiva, P.M.G.; Napoleão, T.H.; Barros, R.; Pontual, E.V.J.R. Sodium chloride reduces the viability of eggs, larvae, and pupae of Spodoptera frugiperda, as well as delays its development. Res. Soc. Dev. 2022, 11, e29311427232. [Google Scholar] [CrossRef]
- Ibrahim, S.S.; Riveron, J.M.; Stott, R.; Irving, H.; Wondji, C.S. The cytochrome P450 CYP6P4 is responsible for the high pyrethroid resistance in knockdown resistance-free Anopheles arabiensis. Insect Biochem. Mol. Biol. 2016, 68, 23–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moyes, C.L.; Lees, R.S.; Yunta, C.; Walker, K.J.; Hemmings, K.; Oladepo, F.; Hancock, P.A.; Weetman, D.; Paine, M.J.I.; Ismail, H.M. Assessing cross-resistance within the pyrethroids in terms of their interactions with key cytochrome P450 enzymes and resistance in vector populations. Parasites Vectors 2021, 14, 115. [Google Scholar] [CrossRef] [PubMed]
- Weedall, G.D.; Mugenzi, L.M.J.; Menze, B.D.; Tchouakui, M.; Ibrahim, S.S.; Amvongo-Adjia, N.; Irving, H.; Wondji, M.J.; Tchoupo, M.; Djouaka, R.; et al. A cytochrome P450 allele confers pyrethroid resistance on a major African malaria vector, reducing insecticide-treated bednet efficacy. Sci. Transl. Med. 2019, 11, eaat7386. [Google Scholar] [CrossRef] [Green Version]
- Wondji, C.S.; Hearn, J.; Irving, H.; Wondji, M.J.; Weedall, G. RNAseq-based gene expression profiling of the Anopheles funestus pyrethroid-resistant strain FUMOZ highlights the predominant role of the duplicated CYP6P9a/b cytochrome P450s. G3 2022, 12, jkab352. [Google Scholar] [CrossRef] [PubMed]
- Mancini, M.V.; Damiani, C.; Accoti, A.; Tallarita, M.; Nunzi, E.; Cappelli, A.; Bozic, J.; Catanzani, R.; Rossi, P.; Valzano, M.; et al. Estimating bacteria diversity in different organs of nine species of mosquito by next generation sequencing. BMC Microbiol. 2018, 18, 126. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Allam, M.; Kwenda, S.; Khumalo, Z.T.H.; Ismail, A.; Oliver, S.V. The dynamic gut microbiota of zoophilic members of the Anopheles gambiae complex (Diptera: Culicidae). Sci. Rep. 2022, 12, 1495. [Google Scholar] [CrossRef]
- Xiang, M.A.; Linser, P.J.; Price, D.A.; Harvey, W.R. Localization of two Na+- or K+-H+ antiporters, AgNHA1 and AgNHA2, in Anopheles gambiae larval Malpighian tubules and the functional expression of AgNHA2 in yeast. J. Insect Physiol. 2012, 58, 570–579. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.; Bellingrath-Kimura, S.D.; Mishra, J.; Arora, N.K. Salt-Tolerant Plant Growth Promoting Rhizobacteria for Enhancing Crop Productivity of Saline Soils. Front. Microbiol. 2019, 10, 2791. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Xu, Z.; Liu, H.; Liu, Y.; Zhou, Y.; Meng, C.; Ma, S.; Xie, Z.; Li, Y.; Zhang, C.S. Patterns in the Microbial Community of Salt-Tolerant Plants and the Functional Genes Associated with Salt Stress Alleviation. Microbiol. Spectr. 2021, 9, e0076721. [Google Scholar] [CrossRef] [PubMed]
- Dada, N.; Lol, J.C.; Benedict, A.C.; López, F.; Sheth, M.; Dzuris, N.; Padilla, N.; Lenhart, A. Pyrethroid exposure alters internal and cuticle surface bacterial communities in Anopheles albimanus. ISME J. 2019, 13, 2447–2464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickson, L.B.; Jiolle, D.; Minard, G.; Moltini-Conclois, I.; Volant, S.; Ghozlane, A.; Bouchier, C.; Ayala, D.; Paupy, C.; Moro, C.V.; et al. Carryover effects of larval exposure to different environmental bacteria drive adult trait variation in a mosquito vector. Sci. Adv. 2017, 3, e1700585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Gilbreath, T.M., 3rd; Kukutla, P.; Yan, G.; Xu, J. Dynamic gut microbiome across life history of the malaria mosquito Anopheles gambiae in Kenya. PLoS ONE 2011, 6, e24767. [Google Scholar] [CrossRef] [Green Version]
- Terenius, O.; Lindh, J.M.; Eriksson-Gonzales, K.; Bussière, L.; Laugen, A.T.; Bergquist, H.; Titanji, K.; Faye, I. Midgut bacterial dynamics in Aedes aegypti. FEMS Microbiol. Ecol. 2012, 80, 556–565. [Google Scholar] [CrossRef] [Green Version]
- E Silva, B.; Matsena Zingoni, Z.; Koekemoer, L.L.; Dahan-Moss, Y.L. Microbiota identified from preserved Anopheles. Malar. J. 2021, 20, 230. [Google Scholar] [CrossRef]
- Feng, Y.; Peng, Y.; Song, X.; Wen, H.; An, Y.; Tang, H.; Wang, J. Anopheline mosquitoes are protected against parasite infection by tryptophan catabolism in gut microbiota. Nat. Microbiol. 2022, 7, 707–715. [Google Scholar] [CrossRef]
- Romoli, O.; Gendrin, M. The tripartite interactions between the mosquito, its microbiota and Plasmodium. Parasites Vectors 2018, 11, 200. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Rani, J.; Chauhan, C.; Kumari, S.; Tevatiya, S.; Das De, T.; Savargaonkar, D.; Pandey, K.C.; Dixit, R. Altered Gut Microbiota and Immunity Defines Plasmodium vivax Survival in Anopheles stephensi. Front. Immunol. 2020, 11, 609. [Google Scholar] [CrossRef]
- Zouache, K.; Martin, E.; Rahola, N.; Gangue, M.F.; Minard, G.; Dubost, A.; Van, V.T.; Dickson, L.; Ayala, D.; Lambrechts, L.; et al. Larval habitat determines the bacterial and fungal microbiota of the mosquito vector Aedes aegypti. FEMS Microbiol. Ecol. 2022, 98, fiac016. [Google Scholar] [CrossRef]
- Vézilier, J.; Nicot, A.; Lorgeril, J.; Gandon, S.; Rivero, A. The impact of insecticide resistance on Culex pipiens immunity. Evol. Appl. 2013, 6, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Minard, G.; Mavingui, P.; Moro, C.V. Diversity and function of bacterial microbiota in the mosquito holobiont. Parasites Vectors 2013, 6, 146. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, A.; Patel, N.F.; Allam, M.; Chan, W.-Y.; Mohale, T.; Ismail, A.; Oliver, S.V. Marked Effects of Larval Salt Exposure on the Life History and Gut Microbiota of the Malaria Vector Anopheles merus (Diptera: Culicidae). Insects 2022, 13, 1165. https://doi.org/10.3390/insects13121165
Singh A, Patel NF, Allam M, Chan W-Y, Mohale T, Ismail A, Oliver SV. Marked Effects of Larval Salt Exposure on the Life History and Gut Microbiota of the Malaria Vector Anopheles merus (Diptera: Culicidae). Insects. 2022; 13(12):1165. https://doi.org/10.3390/insects13121165
Chicago/Turabian StyleSingh, Ashmika, Nashrin F. Patel, Mushal Allam, Wai-Yin Chan, Thabo Mohale, Arshad Ismail, and Shüné V. Oliver. 2022. "Marked Effects of Larval Salt Exposure on the Life History and Gut Microbiota of the Malaria Vector Anopheles merus (Diptera: Culicidae)" Insects 13, no. 12: 1165. https://doi.org/10.3390/insects13121165