
Comparative Analysis of Mitogenomes of Chironomus (Diptera: Chironomidae)
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Taxon Sampling and Sequencing
2.2. Genome Assembly and Annotation
2.3. Sequence Analyses
2.4. Phylogenetic Analyses
3. Results and Discussion
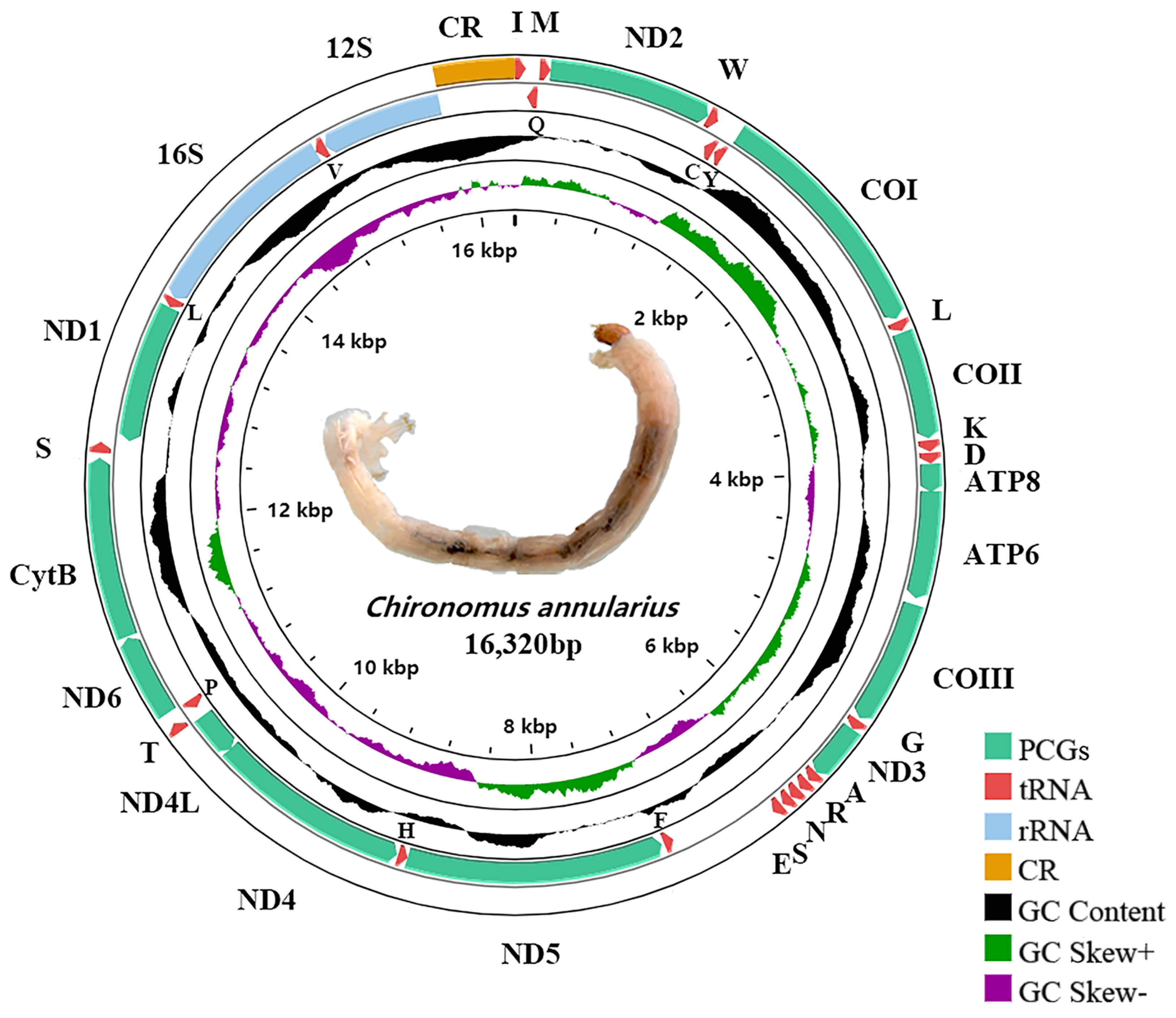
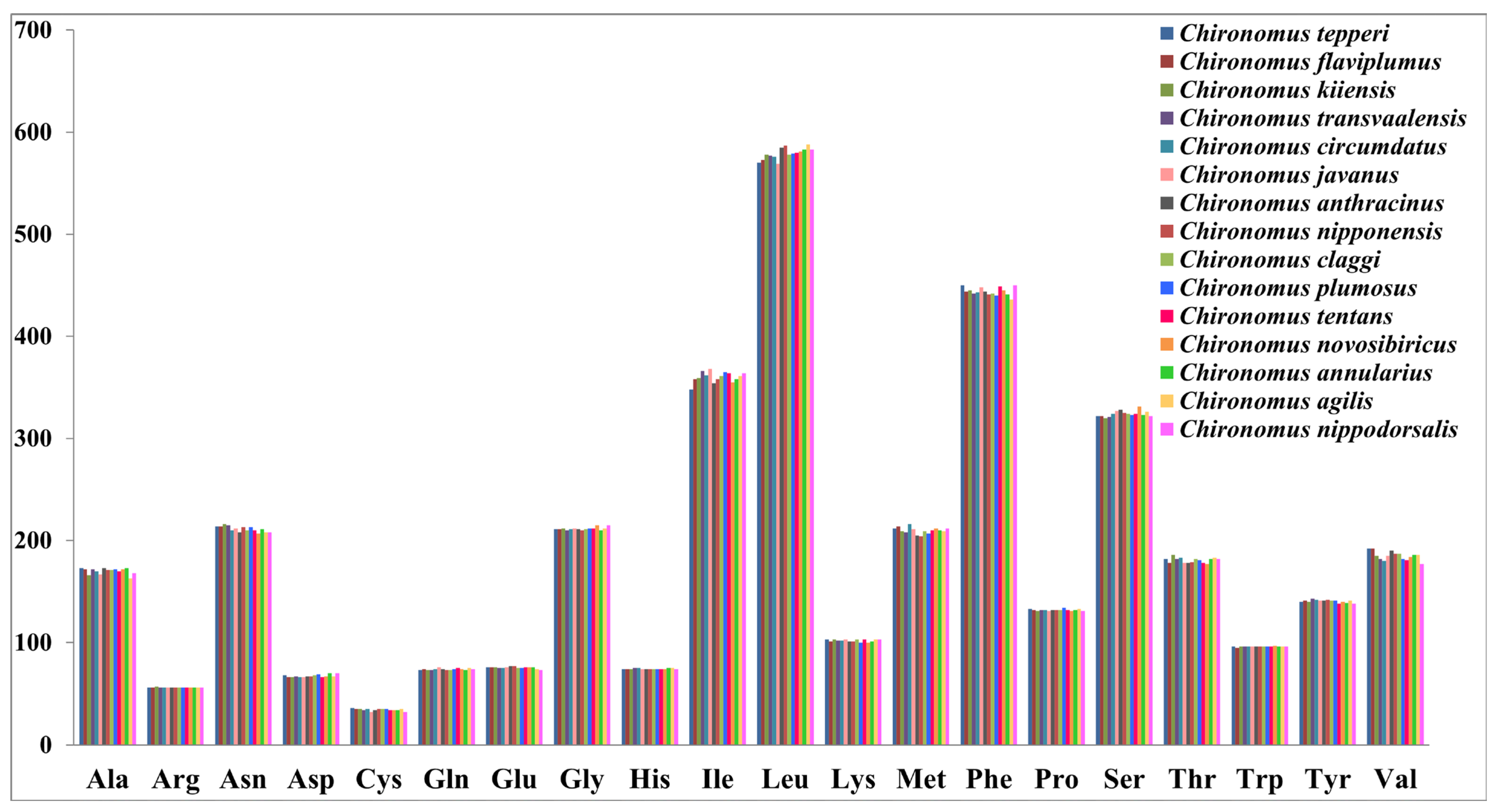
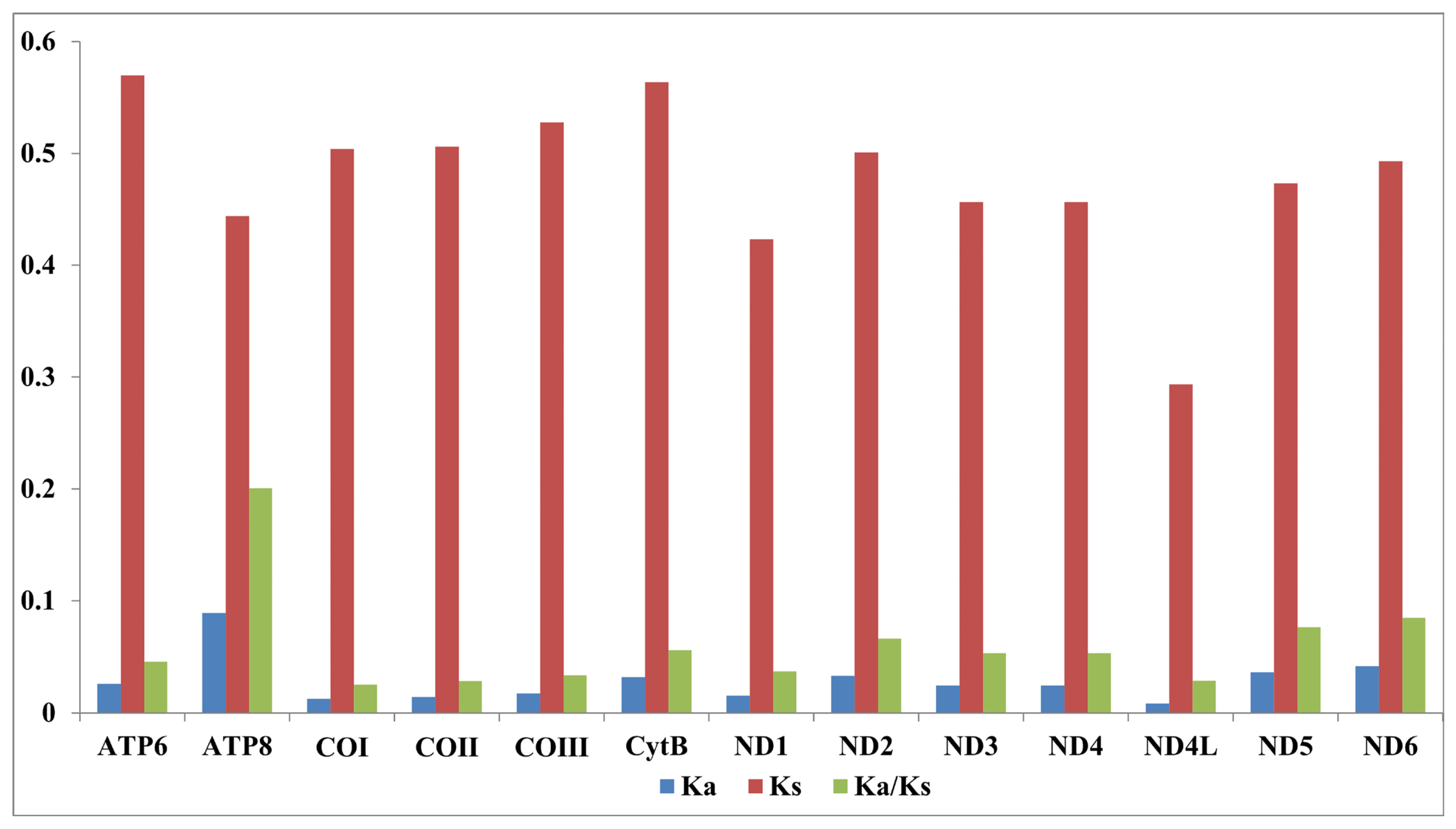
3.1. Basic Structure of Chironomus Mitogenomes
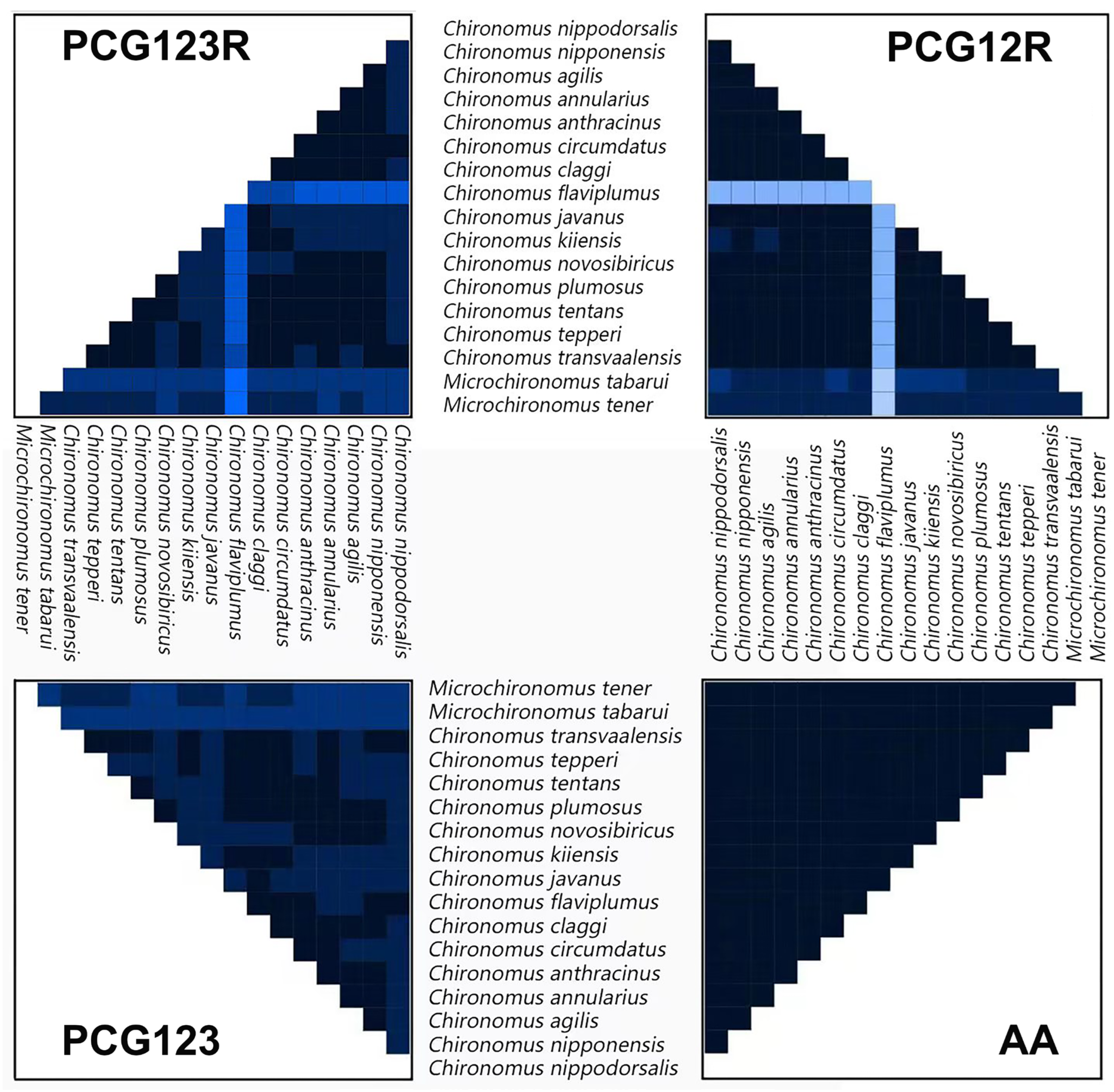
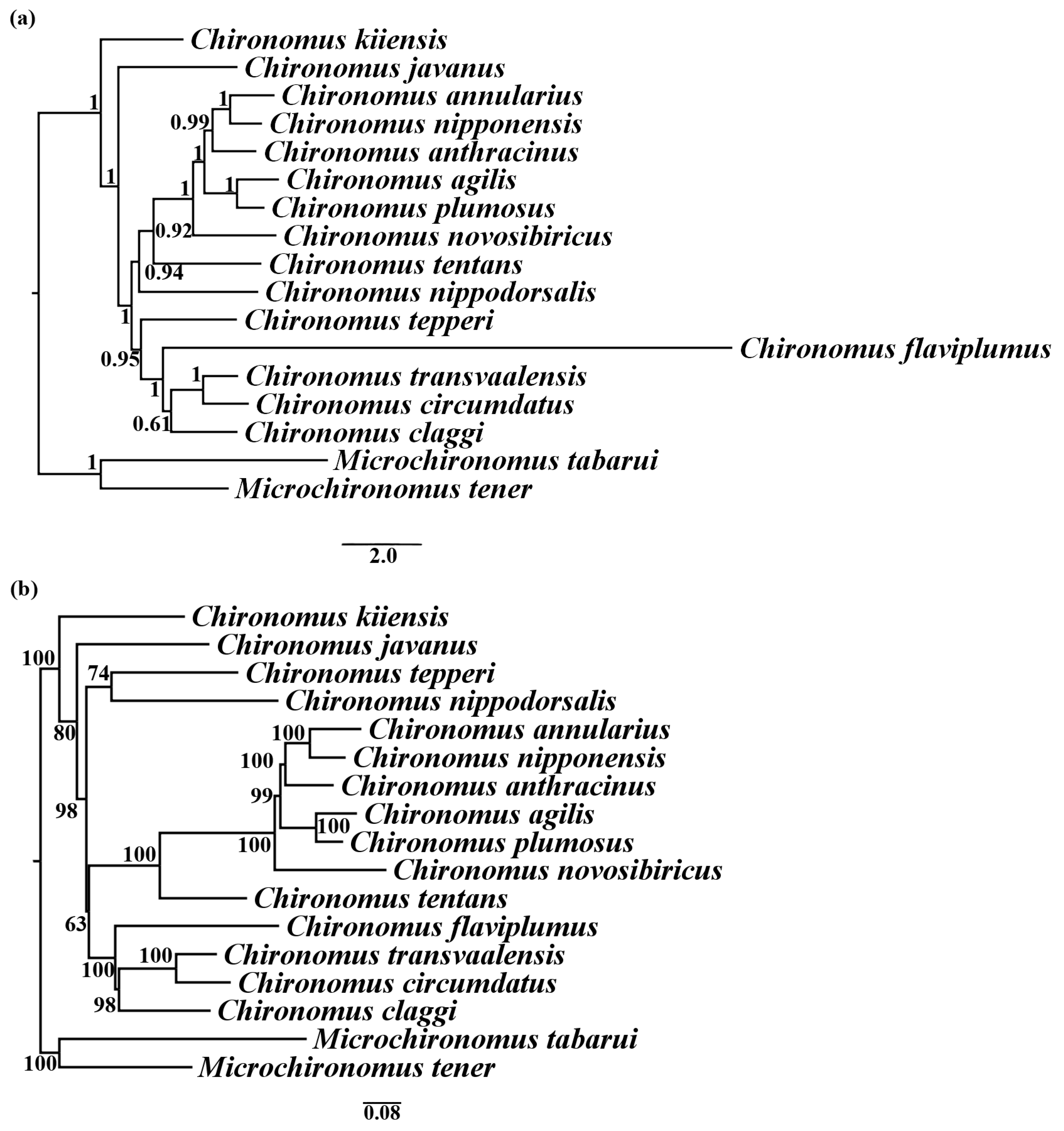
3.2. Phylogenetic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, W.M. The mitochondrial genome of animals. Mol. Evol. Genet. 1985, 22, 95–130. [Google Scholar]
- Zhang, X.; Yang, D.; Kang, Z. New data on the mitochondrial genome of Nematocera (lower Diptera): Features, structures and phylogenetic implications. Zool. J. Linn. Soc. 2022, zlac012. [Google Scholar] [CrossRef]
- Wang, Q.; Huang, J.; Wu, H. Mitogenomes provide insights into the phylogeny of Mycetophilidae (Diptera: Sciaroidea). Gene 2021, 783, 145564. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Zang, H.; Ye, X.; Peng, L.; Wang, B.; Lian, G.; Sun, C. Comparative Mitogenomic Analyses of Hydropsychidae Revealing the Novel Rearrangement of Protein-Coding Gene and tRNA (Trichoptera: Annulipalpia). Insects 2022, 13, 759. [Google Scholar] [CrossRef]
- Ma, Z.; Li, R.; Zhu, B.; Zheng, X.; Zhou, C. Comparative Mitogenome Analyses of Subgenera and Species Groups in Epeorus (Ephemeroptera: Heptageniidae). Insects 2022, 13, 599. [Google Scholar] [CrossRef]
- Brown, W.M.; George, M.; Wilson, A.C. Rapid evolution of animal mitochondrial DNA. Proc. Natl. Acad. Sci. USA 1979, 76, 1967–1971. [Google Scholar] [CrossRef] [Green Version]
- Curole, J.P.; Kocher, T.D. Mitogenomics: Digging deeper with complete mitochondrial genomes. Trends Ecol. Evol. 1999, 14, 394–398. [Google Scholar] [CrossRef]
- Zheng, C.-G.; Liu, Z.; Zhao, Y.-M.; Wang, Y.; Bu, W.-J.; Wang, X.-H.; Lin, X.-L. First Report on Mitochondrial Gene Rearrangement in Non-Biting Midges, Revealing a Synapomorphy in Stenochironomus Kieffer (Diptera: Chironomidae). Insects 2022, 13, 115. [Google Scholar] [CrossRef]
- Lin, X.L.; Zhao, Y.M.; Yan, L.P.; Liu, W.B.; Bu, W.J.; Wang, X.H.; Zheng, C.G. Mitogenomes provide new insights into the evolutionary history of Prodiamesinae (Diptera: Chironomidae). Zool. Scr. 2022, 51, 119–132. [Google Scholar] [CrossRef]
- Zheng, C.-G.; Zhu, X.-X.; Yan, L.-P.; Yao, Y.; Bu, W.-J.; Wang, X.-H.; Lin, X.-L. First complete mitogenomes of Diamesinae, Orthocladiinae, Prodiamesinae, Tanypodinae (Diptera: Chironomidae) and their implication in phylogenetics. PeerJ. 2021, 9, e11294. [Google Scholar] [CrossRef]
- Chen, Z.-T.; Du, Y.-Z. Comparison of the complete mitochondrial genome of the stonefly Sweltsa longistyla (Plecoptera: Chloroperlidae) with mitogenomes of three other stoneflies. Gene 2015, 558, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Lei, Z.; Li, W.; Zhang, W.; Zhou, C. Comparative mitogenomic analysis of heptageniid mayflies (Insecta: Ephemeroptera): Conserved intergenic spacer and trna gene duplication. Insects 2021, 12, 170. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Ye, Z.; Zhu, X.; Zhang, H.; Dong, X.; Chen, P.; Bu, W. Integrative taxonomy uncovers hidden species diversity in the rheophilic genus Potamometra (Hemiptera: Gerridae). Zool. Scr. 2020, 49, 174–186. [Google Scholar] [CrossRef]
- Ge, X.; Peng, L.; Vogler, A.P.; Morse, J.C.; Yang, L.; Sun, C.; Wang, B. Massive gene rearrangements of mitochondrial genomes and implications for the phylogeny of Trichoptera (Insecta). Syst. Entomol. 2022, in press. [Google Scholar] [CrossRef]
- Gao, Y.; Ge, Y.; Yan, L.; Vikhrev, N.E.; Wang, Q.; Butterworth, N.J.; Zhang, D. Phylogenetic Analyses Support the Monophyly of the Genus Lispe Latreille (Diptera: Muscidae) with Insights into Intrageneric Relationships. Insects 2022, 13, 1015. [Google Scholar] [CrossRef]
- Cranston, P.S.; Dillon, M.E.; Pinder, L.C.V.; Reiss, F. The Adult Males of Chironominae (Diptera, Chironomidae) of the Holarctic Region—Keys and Diagnoses. In Chironomidae of the Holarctic Region. Keys and Diagnoses. Part 3—Adult Males; Wiederholm, T., Ed.; Entomologica Scandinavica: Lund, Sweden, 1989; Volume 34, pp. 353–502. [Google Scholar]
- Cranston, P.S.; Hardy, N.B.; Morse, G.E. A dated molecular phylogeny for the Chironomidae (Diptera). Syst. Entomol. 2012, 37, 172–188. [Google Scholar] [CrossRef]
- Martin, J.; Blinov, A.; Alieva, E.; Hirabayashi, K. A molecular phylogenetic investigation of the genera closely related to Chironomus Meigen (Diptera: Chironomidae). In Contributions to the Systematics and Ecology of Aquatic Diptera. A Tribute to Ole A. Sæther; Andersen, T., Ed.; Caddis Press: Columbus, OH, USA, 2007; pp. 193–203. [Google Scholar]
- Beckenbach, A.T. Mitochondrial genome sequences of Nematocera (lower Diptera): Evidence of rearrangement following a complete genome duplication in a winter crane fly. Genome Biol. Evol. 2012, 4, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Park, K.; Kim, W.-S.; Park, J.-W.; Kwak, I.-S. Complete mitochondrial genome of Chironomus flaviplumus (Diptera: Chironomidae) collected in Korea. Mitochondrial DNA Part B 2021, 6, 2843–2844. [Google Scholar] [CrossRef]
- Liu, C.; Xu, G.; Lei, T.; Qi, X. The complete mitochondrial genome of a tropical midge Chironomus kiiensis Tokunaga, 1936 (Diptera: Chironomidae). Mitochondrial DNA Part B 2022, 7, 211–212. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Kong, F.-Q.; Zhao, Y.-C.; Chen, J.-L.; Lin, X.-L. First report of the complete mitogenome of Microchironomus tabarui Sasa, 1987 (Diptera, Chironomidae) from Hebei Province, China. Mitochondrial DNA Part B 2021, 6, 2845–2846. [Google Scholar] [CrossRef]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Leung, H.C.; Yiu, S.-M.; Chin, F.Y. IDBA-UD: A de novo assembler for single-cell and metagenomic sequencing data with highly uneven depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, W.; Le, S.; Li, Y.; Hu, F. SeqKit: A cross-platform and ultrafast toolkit for FASTA/Q file manipulation. PLoS ONE 2016, 11, e0163962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Grant, J.R.; Stothard, P. The CGView Server: A comparative genomics tool for circular genomes. Nucleic Acids Res. 2008, 36, 181–184. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [Green Version]
- Kück, P.; Longo, G.C. FASconCAT-G: Extensive functions for multiple sequence alignment preparations concerning phylogenetic studies. Front. Zool. 2014, 11, 81. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.H. DAMBE5: A comprehensive software package for data analysis in molecular biology and evolution. Mol. Biol. Evol. 2013, 30, 1720–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kück, P.; Meid, S.A.; Groß, C.; Wägele, J.W.; Misof, B. AliGROOVE–visualization of heterogeneous sequence divergence within multiple sequence alignments and detection of inflated branch support. BMC Bioinform. 2014, 15, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.L.; Liu, Z.; Yan, L.P.; Duan, X.; Bu, W.J.; Wang, X.H.; Zheng, C.G. Mitogenomes provide new insights of evolutionary history of Boreheptagyiini and Diamesini (Diptera: Chironomidae: Diamesinae). Ecol. Evol. 2022, 12, e8957. [Google Scholar] [CrossRef]
- Ekrem, T.; Willassen, E. Exploring Tanytarsini relationships (Diptera: Chironomidae) using mitochondrial COII gene sequences. Insect Syst. Evol. 2004, 35, 263–276. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sample ID | Life Stage | Sampling Metadata | GenBank Accession | Reference |
|---|---|---|---|---|---|
| Microchironomus tabarui | XL3993 | Adult male | Hengshui, Hebei, China, 37.651626°N, 115.650831°E, 1 September 2020, leg. X.-Y. Liu | MZ261913 | [23] |
| Microchironomus tener | XL1462 | Adult male | Changjiang, Hainan, China, 19.11463°N, 109.08419°E, 13 March 2016, leg. B.-J. Sun | ON975027 | this study |
| Chironomus tepperi | JN861749 | NA | NA | JN861749 | [19] |
| Chironomus flaviplumus | CNUISI-020005203 | Larva | Yeondeung stream, Yeosu, South Korea 34°45′26.0″ N, 127°42′51.2″ E, May 2020 | MW770891 | [20] |
| Chironomus kiiensis | BSZ21 | Larva | Lishui, Zhejiang, China, 28°39′30′′ N, 120°5′29′′ E, August 2019, leg. X. Qi | MZ150770 | [21] |
| Chironomus transvaalensis | NAM96 | Larva | Goreangab Dam, Khomas, Windhoek, Namibia, 22.5267°S, 17.0153°E, 3 December 2018, leg. X.-L. Lin | ON975023 | this study |
| Chironomus circumdatus | NEC119 | Larva | Kouembélia, Tontouta, New Caledonia, 22.0083056°S, 166.2062775°E, 11 May 2020, leg. N. Mary | ON975024 | this study |
| Chironomus javanus | NLCH300 | Adult male | Yizhang, Chenzhou, Hunan, China, 24.9854183°N, 112.914357°E, 30 August 2020, leg. X.-L. Lin | ON975025 | this study |
| Chironomus anthracinus | XL575 | Adult male | Lian lake, Trondheim, Norway, 63.39989°N, 10.31761°E, 17 June 2016, leg. X.-L. Lin | ON975026 | this study |
| Chironomus nipponensis | X2896 | Larva | Laotuding, Huanren, Benxi, Liaoning, China, 41.2894°N, 124.8980°E, 3 September 2014, leg. C. Song | ON975028 | this study |
| Chironomus claggi | XL2930 | Larva | Maoyangzhen, Wuzhishan Hainan, China, 18.93696°N, 109.50804°E, 6 December 2010, leg. F.-Q. Kong | ON975029 | this study |
| Chironomus plumosus | XL3435 | Larva | Yuqiao Reservoir, Jizhou, Tianjin, China, 40.01974°N, 117.6389°E, 21 November 2019, leg. H.-J. Yu | ON975030 | this study |
| Chironomus tentans | XL3813 | Larva | Naqu, Xizang, China, 31.621813°N, 91.739874°E, 3 September 2020, leg. Y. Peng | ON975031 | this study |
| Chironomus novosibiricus | XL3834 | Larva | Zegucuo, Shannan, Xizang, China, 28.754153°N, 91.676359°E, 30 August 2020, leg. Y. Peng | ON975032 | this study |
| Chironomus annularius | XL3838 | Larva | Zegucuo, Shannan, Xizang, China, 28.754153°N, 91.676359°E, 30 August 2020, leg. Y. Peng | ON975033 | this study |
| Chironomus agilis | XL4188 | Adult male | Chun′an, Hangzhou, Zhejiang, China, 29.567168°N, 118.86825°E, 8 May 2021, leg. Y.-Y. Han | ON975034 | this study |
| Chironomus nippodorsalis | XL4371 | Adult female | Hefeng, Hubei, China, 29.89269645°N, 110.0287267°E, 12-July-15, leg. Q. Wang | ON975035 | this study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.-Y.; Zhao, Y.-M.; Guo, B.-X.; Li, C.-H.; Sun, B.-J.; Lin, X.-L. Comparative Analysis of Mitogenomes of Chironomus (Diptera: Chironomidae). Insects 2022, 13, 1164. https://doi.org/10.3390/insects13121164
Li S-Y, Zhao Y-M, Guo B-X, Li C-H, Sun B-J, Lin X-L. Comparative Analysis of Mitogenomes of Chironomus (Diptera: Chironomidae). Insects. 2022; 13(12):1164. https://doi.org/10.3390/insects13121164
Chicago/Turabian StyleLi, Shu-Yi, Yan-Min Zhao, Bing-Xin Guo, Chen-Hong Li, Bing-Jiao Sun, and Xiao-Long Lin. 2022. "Comparative Analysis of Mitogenomes of Chironomus (Diptera: Chironomidae)" Insects 13, no. 12: 1164. https://doi.org/10.3390/insects13121164