

Evaluation of Phosphine Resistance in Populations of Sitophilus oryzae, Oryzaephilus surinamensis and Rhyzopertha dominica in the Czech Republic



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tested Insects and Populations
2.2. Laboratory Resistance Tests and Statistics
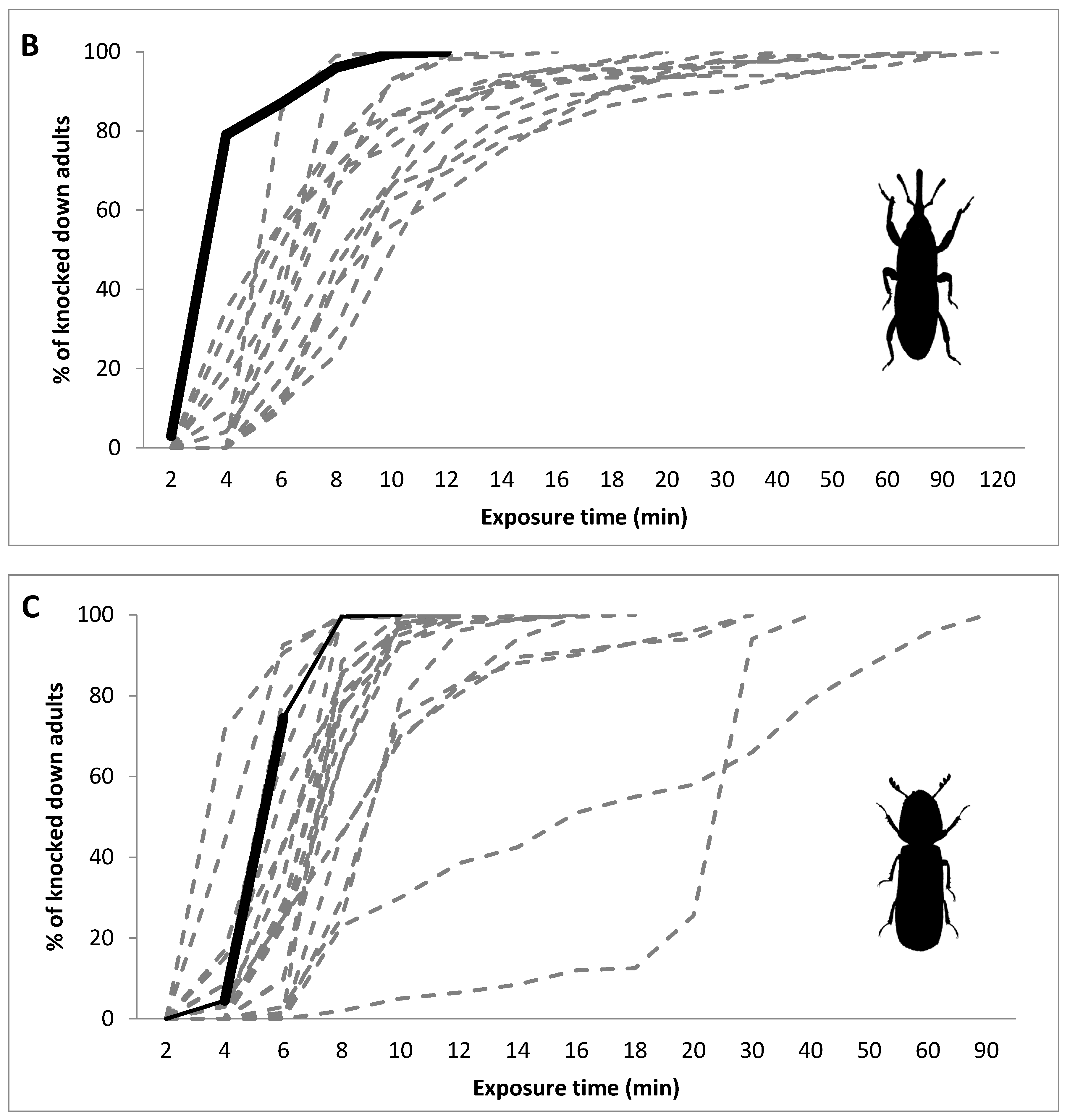
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding

Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gerken, A.R.; Morrison, W.R. Pest management in the postharvest agricultural supply chain under climate change. Front. Agron. 2022, 4, 918845. [Google Scholar] [CrossRef]
- Athanassiou, C.G.; Arthur, F.H. Recent Advances in Stored Product Protection; Springer: Berlin/Heidelberg, Germany, 2018; p. 273. [Google Scholar]
- Stejskal, V.; Bostlova, M.; Nesvorna, M.; Volek, V.; Dolezal, V.; Hubert, J. Comparison of the resistance of mono- and multilayer packaging films to stored-product insects in a laboratory test. Food Control 2017, 73, 566–573. [Google Scholar] [CrossRef]
- Hubert, J.; Erban, T.; Nesvorna, M.; Stejskal, V. Emerging risk of infestation and contamination of dried fruits by mites in the Czech Republic. Food Addit. Contam. Part A 2011, 28, 1129–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, C.; Athanassiou, C.; Carvalho, M.O.; Emekci, M.; Gvozdenac, S.; Hamel, D.; Riudavets, J.; Stejskal, V.; Trdan, S.; Trematerra, P. Changes in the distribution and pest risk of stored product insects in Europe due to global warming: Need for pan-European pest monitoring and improved food-safety. J. Stored Prod. Res. 2022, 97, 101977. [Google Scholar] [CrossRef]
- Stejskal, V.; Vendl, T.; Kolar, V.; Li, Z.; Aulicky, R. First population quantification of the infestation of legumes by stored-product bruchids imported in freight containers into Europe. Bull. Insectol. 2020, 73, 233–239. [Google Scholar]
- Stejskal, V.; Vendl, T.; Aulicky, R.; Athanassiou, C. Synthetic and Natural Insecticides: Gas, Liquid, Gel and Solid Formulations for Stored-Product and Food-Industry Pest Control. Insects 2021, 12, 590. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, S.; Casada, M.E.; Maghirang, R.G.; Wei, M. Evolution of phosphine from aluminum phosphide pellets. Trans. ASABE 2021, 64, 615–624. [Google Scholar] [CrossRef]
- Bell, C.H. Fumigation in the 21st century. Crop Prot. 2000, 19, 563–569. [Google Scholar] [CrossRef]
- Agrafioti, P.; Kaloudis, E.; Bantas, S.; Sotiroudas, V.; Athanassiou, C.G. Modeling the distribution of phosphine and insect mortality in cylindrical grain silos with computational fluid dynamics: Validation with field trials. Comput. Electron. Agric. 2020, 173, 105383. [Google Scholar] [CrossRef]
- Brabec, D.; Kaloudis, E.; Athanassiou, C.G.; Campbell, J.; Agrafioti, P.; Scheff, D.S.; Bantas, S.; Sotiroudas, V. Fumigation monitoring and modeling of hopper-bottom railcars loaded with corn grits. J. Biosyst. Eng. 2022, 47, 358–369. [Google Scholar] [CrossRef]
- Aulicky, R.; Stejskal, V. Efficacy and limitations of phosphine “spot-fumigation” against five Coleoptera species of stored product pests in wheat in a grain store—Short note. Plant Prot. Sci. 2015, 51, 33–38. [Google Scholar] [CrossRef]
- Nayak, M.K.; Daglish, G.J.; Phillips, T.W.; Ebert, P.R. resistance to the fumigant phosphine and its management in insect pests of stored products: A global perspective. Annu. Rev. Entomol. 2020, 65, 333–350. [Google Scholar] [CrossRef] [Green Version]
- Agrafioti, P.; Athanassiou, C.G.; Nayak, M.K. Detection of phosphine resistance in major stored-product insects in Greece and evaluation of a field resistance test kit. J. Stored Prod. Res. 2019, 82, 40–47. [Google Scholar] [CrossRef]
- Rafter, M.A.; McCulloch, G.A.; Daglish, G.J.; Walter, G.H. Progression of phosphine resistance in susceptible Tribolium castaneum (Herbst) populations under different immigration regimes and selection pressures. Evol. Appl. 2017, 10, 907–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Champ, R.; Dyte, C.E. Report of the FAO global survey of pesticide susceptibility of stored grain pests. In FAO Plant Production and Protection Series N 5 FAO; FAO: Rome, Italy, 1976. [Google Scholar]
- Rajendran, S.; Parveen, H.; Begum, K.; Chethana, R. Influence of phosphine on hatching of Cryptolestes ferrugineus (Coleoptera: Cucujidae), Lasioderma serricorne (Coleoptera: Anobiidae) and Oryzaephilus surinamensis (Coleoptera: Silvanidae). Pest Manag. Sci. 2004, 60, 1114–1118. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, M.A.G.; Lrďa, F.; Guedes, R.N.C.; Sousa, A.H.; Tótola, M.R. Phosphine resistance in Brazilian populations of Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae). J. Stored Prod. Res. 2009, 45, 71–74. [Google Scholar] [CrossRef]
- Opit, G.P.; Phillips, T.W.; Aikins, M.J.; Hasan, M.M. Phosphine resistance in Tribolium castaneum and Rhyzopertha dominica from stored wheat in Oklahoma. J. Econ. Entomol. 2012, 105, 1107–1114. [Google Scholar] [CrossRef] [Green Version]
- Kaur, R.; Nayak, M.K. Developing effective fumigation protocols to manage strongly phosphine resistant Cryptolestes ferrugineus (Stephens) (Coleoptera: Laemophloeidae). Pest Manag. Sci. 2015, 71, 1297–1302. [Google Scholar] [CrossRef] [Green Version]
- Mangoba, M.A.A.; de Guzman Alvindia, D. Phosphine resistance in psocid, Liposcelis bostrychophila (Psocoptera) in the Philippines. Int. J. Trop. Insect Sci. 2021, 41, 439–445. [Google Scholar] [CrossRef]
- Sakka, M.K.; Vontas, M.R.J.; Götze, M.C.; Allegra, J.; Gehard, J.; Athanassiou, C.G. Evaluation of tolerance/resistance to phosphine of stored product beetle populations from Europe, by using different diagnostic methods. In Proceedings of the 12th International Working Conference on Stored Product Protection, Berlin, Germany, 7–11 October 2018; JKI: Berlin, Germany; pp. 1003–1008.
- Aulicky, R.; Stejskal, V.; Frydova, B.; Athanassiou, C.G. Susceptibility of two strains of the confused flour beetle (Coleoptera: Tenebrionidae) following phosphine structural mill fumigation: Effects of concentration, temperature, and flour deposits. J. Econ. Entomol. 2015, 108, 2823–2830. [Google Scholar] [CrossRef]
- Aulicky, R.; Stejskal, V.; Frydova, B. Field validation of phosphine efficacy on the first recorded resistant strains of Sitophilus granarius and Tribolium castaneum from the Czech Republic. J. Stored Prod. Res. 2019, 81, 107–113. [Google Scholar] [CrossRef]
- FAO. Recommended methods for the detection and measurement of resistance of agricultural pests to pesticides: Tentative method for adults of some major species of stored cereals with methyl bromide and phosphine—FAO Method No 16. Plant Prot. Bull. 1975, 23, 12–25. [Google Scholar]
- Danso, J.K.; Opit, G.P.; Noden, B.H.; Giles, K.L. Estimating discriminating doses of phosphine for adults of eight species of psocids of genera Liposcelis (Psocodea: Liposcelididae) and Lepinotus (Psocodea: Trogiidae). J. Stored Prod. Res. 2022, 99, 102025. [Google Scholar] [CrossRef]
- Athanassiou, C.G.; Kavallieratos, N.G.; Brabec, D.L.; Oppert, B.; Guedes, R.N.C.; Campbell, J.F. From immobilization to recovery: Towards the development of a rapid diagnostic indicator for phosphine resistance. J. Stored Prod. Res. 2019, 80, 28–33. [Google Scholar] [CrossRef]
- Agrafioti, P.; Sotiroudas, V.; Kaloudis, S.; Bantas, S.; Athanassiou, C.G. Real time monitoring of phophine and insect mortality in different storage facilities. J. Stored Prod. Res. 2020, 89, 101726. [Google Scholar] [CrossRef]
- Reichmuth, C.A. Quick test to determine phosphine resistance in stored products research. GASGA Newsl. 1991, 15, 14–15. [Google Scholar]
- Bell, C.H.; Savvidou, N.; Mills, K.A.; Bradberry, S.; Barlow, M.L. A same-day test for detecting resistance to phosphine. In Proceedings of the 6th International Working Conference on Stored-Prod, Canberra, Australia, 17–23 April 1994; Volume 1, pp. 41–44. [Google Scholar]
- Cao, Y.; Wang, D. Relationship between phosphine resistance and narcotic knockdown in Tribolium castaneum (Herbst), Sitophilus oryzae (L.), and S. zeamais (Motsch). In Proceedings of the Conference Controlled Atmosphere and Fumigation in Stored-products, Adelaide, Australia, 1–4 August 2000; pp. 609–616. [Google Scholar]
- Cato, A.; Afful, E.; Nayak, M.N.; Phillips, T.W. Evaluation of knockdown bioassay methods to assess phosphine resistance in the red flour beetle, Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae). Insects 2019, 10, 140. [Google Scholar] [CrossRef] [Green Version]
- Nayak, M.K.; Kaur, R.; Jagadeesan, R.; Pavic, H.; Phillips, T.W.; Daglish, G.J. Development of a quick knockdown test for diagnosing resistance to Phosphine in Sitophilus oryzae (Coleoptera: Curculionidae), a major pest of stored products. J. Econ. Entomol. 2019, 112, 1975–1982. [Google Scholar] [CrossRef]
- Afful, E.; Cato, A.; Nayak, M.K.; Phillips, T.W. A rapid assay for the detection of resistance to phosphine in the lesser grain borer, Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae). J. Stored Prod. Res. 2021, 91, 101776. [Google Scholar] [CrossRef]
- Steuerwald, R.; Dierks-Lange, H.; Schmitt, S. Rapid bioassay for determining the phosphine tolerance. In Proceedings of the 9th International Working Conference on Stored-Product Prot., Campinas, Sao Paolo, BRAPOS, Passo Fundo, Brazil, 15–18 October 2006; pp. 306–311. [Google Scholar]
- Goetze, C.; Steuerwald, R.; Agrafioti, P.; Jakob, G.K.; Athanassiou, C.G. It all started with a kit: The Detia Degesch phosphine tolerance test and new data in the field of phosphine fumigation. In Proceedings of the 11th International Conference on Controlled Atmospheres and Fumigation in Stored Products (CAF 2020), CAF Permanent Committee Secretariat, Winnipeg, MB, Canada, 11–13 June 2021; pp. 35–41. [Google Scholar]
- Goetze, C.; Sakka, M.K.; Agrafioti, P.; Athanassiou, C.G. Resistance here, resistance there, resistance everywhere! A dispute about phosphine and its use in the light of best management practice. IOBC-WPRS Bull. 2022, 159, 206–209. [Google Scholar]
- Stejskal, V.; Hubert, J.; Aulicky, R.; Kucerova, Z. Overview of present and past and pest associated risks in stored food and feed products: European perspective. J. Stored Prod. Res. 2015, 64, 122–132. [Google Scholar] [CrossRef]
- Niang, E.H.A.; Konata, L.; Diallo, M.; Faye, O.; Dia, I. Patterns of insecticide resistance and knock down resistance (kdr) in malaria vectors An. arabiensis, An. coluzzii and An. gambiae from sympatric areas in Senegal. Parasit. Vect. 2016, 9, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Hu, C.T.; Yang, J.; Joyce, L.A.; Qiu, M.; Ward, M.D.; Kahr, B. Manipulating solid forms of contact insecticides for infectious disease prevention. J. Am. Chem. Soc. 2019, 141, 16858–16864. [Google Scholar] [CrossRef] [PubMed]
- Koçak, E.; Yılmaz, A.; Alpkent, Y.N.; Ertürk, S. Phosphine resistance to some coleopteran pests in stored grains across Turkey. In Proceedings of the 11th International Conference on the IOBC-WPRS Bulletin, Ljubljana, Slovenija, 3–5 July 2017; Volume 130, pp. 303–310. [Google Scholar]
- Stejskal, V.; Vendl, T.; Li, Z.; Aulicky, R. Efficacy of visual evaluation of insect-damaged kernels of malting barley by Sitophilus granarius from various observation perspectives. J. Stored Prod. Res. 2020, 89, 101711. [Google Scholar] [CrossRef]
- Vendl, T.; Frankova, M.; Aulicky, R.; Stejskal, V. First record of the development of Sitophilus oryzae on two rodent bait formulations and literature overview of stored product arthropods infestations in rodent baits. J. Stored Prod. Res. 2020, 86, 101557. [Google Scholar] [CrossRef]
- Agrafioti, P.; Sotiroudas, V.; Bantas, S.; Athanassiou, C.G. Concentration-Time Relationships in Phosphine Fumigation on Different Species and Strains: The UTH Protocol. In Proceedings of the 13th IOBC-WPRS Meeting, Barcelona, Spain, 3–6 October 2022; Agustí, N., Castañé, C., Riudavets, J., Eds.; The Publication Commission of the IOBC-WPRS: Bragança, Portugal, 2022; Volume 159, pp. 214–216. [Google Scholar]
- Lorini, I.; Collins, P.J.; Daglish, G.J.; Nayak, M.K.; Pavic, H. Detection and characterisation of strong resistance to phosphine in Brazilian Rhyzopertha dominica (F.) (Coleoptera: Bostrychidae). Pest Manag. Sci. 2007, 63, 358–364. [Google Scholar] [CrossRef]
- Monro, H.A.U.; Upitis, E.; Bond, E.J. Resistance of a laboratory strain of S. granarius (L.) (Coleoptera: Curculionidae) to phosphine. J. Stored Prod. Res. 1972, 8, 199–207. [Google Scholar] [CrossRef]
- Athanassiou, C.G.; Buchelos, C.T. Detection of stored-wheat beetle species and estimation of population density using unbaited probe traps and grain trier samples. Entomol. Exp. Appl. 2001, 98, 67–78. [Google Scholar] [CrossRef]
- Collins, P.J.; Daglish, G.J.; Bengston, M.; Lambkin, T.M.; Pavic, H. Genetics of resistance to phosphine in Rhyzopertha dominica (Coleoptera: Bostrichidae). J. Econ. Entomol. 2002, 95, 862–869. [Google Scholar] [CrossRef]
- Daglish, G.J. Effect of exposure period on degree of dominance of phosphine resistance in adults of Rhyzopertha dominica (Coleoptera: Bostrychidae) and Sitophilus oryzae (Coleoptera: Curculionidae). Pest Manag. Sci. 2004, 60, 822–826. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Collins, P.J.; Ebert, P.R. Inheritance and characterization of strong resistance to phosphine in Sitophilus oryzae (L.). PLoS ONE 2015, 10, e0124335. [Google Scholar] [CrossRef] [Green Version]
- Gautam, S.G.; Opit, G.P.; Konemann, C.; Shakya, K.; Hosoda, E. Phosphine resistance in saw-toothed grain beetle, Oryzaephilus surinamensis in the United States. J. Stored Prod. Res. 2020, 89, 101690. [Google Scholar] [CrossRef]
- Emery, R.N. A Western Australian farm survey for phosphine-resistant grain beetles. In Proceedings of the 6th International. Working Conference on Stored Product Protection, Canberra, Australia, 17–23 April; Highley, E., Wright, E.J., Banks, H.J., Champ, B.R., Eds.; CAB International: Wallingford, UK, 1994; pp. 98–103. [Google Scholar]
- Herron, G.A. Resistance to grain protectants and phosphine in coleopterous pests of grain stored on farms in New South Wales. J. Aust. Entomol. Soc. 1990, 29, 183–189. [Google Scholar] [CrossRef]
- Pimentel, M.A.G.; Faroni, L.R.D.A.; Tótola, M.R.; Guedes, R.N.C. Phosphine resistance, respiration rate and fitness consequences in stored-product insects. Pest Manag. Sci. 2007, 63, 876–881. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, M.A.G.; Faroni, L.R.D.A.; Batista, M.D.; Silva, F.H.D. Resistance of stored-product insects to phosphine. Pesqui. Agropecuária Bras. 2008, 43, 1671–1676. [Google Scholar] [CrossRef]
- Pimentel, M.A.; Faroni, L.R.D.A.; da Silva, F.H.; Batista, M.D.; Guedes, R.N. Spread of phosphine resistance among Brazilian populations of three species of stored product insects. Neotrop. Entomol. 2010, 39, 101–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, A.H.; Lrd’a, F.; Pimentel, M.A.G.; Guedes, R.N.C. Developmental and population growth rates of phosphine-resistant and-susceptible populations of stored-product insect pests. J. Stored Prod. Res. 2009, 45, 241–246. [Google Scholar] [CrossRef]
- Suthisut, D.; Kengkanpanich, R.; Pedchote, A. Phosphine resistance on Oryzaephilus surinamensis L. in Thailand. Entomol. Zool. Gaz. 2014, 18, 40–48. [Google Scholar]
- Hubhachen, Z.; Opit, G.; Gautam, S.G.; Konemann, C.; Hosoda, E. Phosphine resistance in saw-toothed grain beetle, Oryzaephilus surinamensis (Coleoptera: Silvanidae) in the United States. Jul. Kühn. Arch. 2018, 463, 635–642. [Google Scholar]
- Ferizli, A.G.; Emekci, M.; Tütüncü, S.; Navarro, S. The efficacy of phosphine fumigation against dried fruit pests in Turkey. IOBC WPRS (OILB SROP) Integr. Prot. Stored Prod. 2004, 27, 265–269. [Google Scholar]
- Carvalho, M.O.; Pires, I.; Barbosa, A.; Barros, G.; Riudavets, J.; Garcia, A.C.; Navarro, S. The use of modified atmospheres to control Sitophilus zeamais and Sitophilus oryzae on stored rice in Portugal. J. Stored Prod. Res. 2012, 50, 49–56. [Google Scholar] [CrossRef]
- Stejskal, V.; Vendl, T.; Li, Z.; Aulicky, R. Minimal thermal requirements for development and activity of stored product and food industry pests (Acari, Coleoptera, Lepidoptera, Psocoptera, Diptera and Blattodea): A review. Insects 2019, 10, 149. [Google Scholar] [CrossRef] [Green Version]
- Aulicky, R.; Shah, J.A.; Kolar, V.; Li, Z.; Stejskal, V. Control of stored agro-commodity pests Sitophilus granarius and Callosobruchus chinensis by nitrogen hypoxic atmospheres: Laboratory and Field Validations. Agronomy 2022, 12, 2748. [Google Scholar] [CrossRef]
- EPPO (European and Mediterranean Plant Protection Organization). Recommendations on Fumigation Standards. In Phosphine Fumigation of Stored Products; Fumigation Standard No 18. EPPO Bulletin; John Wiley & Sons: Hoboken, NJ, USA, 1984; Volume 14, pp. 598–599. [Google Scholar]
- Agrafioti, P.; Kaloudis, E.; Athanassiou, C.G. Utilizing low oxygen to mitigate resistance of stored product insects to phosphine. J. Sci. Food Agric. 2022, 102, 6080–6087. [Google Scholar] [CrossRef] [PubMed]
- Aulicky, R.; Kolar, V.; Prokop, J.; Stejskal, V. Field efficacy of brief exposure of adults of six storage pests to nitrogen-controlled atmospheres. Plant Prot. Sci. 2017, 53, 169–176. [Google Scholar]
- Plumier, B.M.; Schramm, M.; Ren, Y.; Maier, D.E. Modeling post-fumigation desorption of phosphine in bulk stored grain. J. Stored Prod. Res. 2020, 85, 101548. [Google Scholar] [CrossRef]
- Rajendran, S.; Gunasekaran, N. The response of phosphine-resistant lesser grain borer Rhyzopertha dominica and rice weevil Sitophilus oryzae in mixed-age cultures to varying concentrations of phosphine. Pest Manag. Sci. 2002, 58, 277–281. [Google Scholar] [CrossRef]
- Lukas, J.; Stejskal, V.; Jarosik, V.; Hubert, J.; Zdarkova, E. Differential natural performance of four Cheyletus predatory mite species in Czech grain stores. J. Stored Prod. Res. 2007, 43, 97–102. [Google Scholar] [CrossRef]
- Kosa-Tass, A.; Bajomi, D.A.; Szilagyi, J.; Verwilghen, F. Sustainable storage of grains by implementing a novel protectant and versatile application technology. In Proceedings of the 13th IOBC-WPRS Meeting, Barcelona, Spain, 3–6 October 2022; Agustí, N., Castañé, C., Riudavets, J., Eds.; The Publication Commission of the IOBC-WPRS: Bragança, Portugal, 2022; Volume 159, p. 229. [Google Scholar]
- Hervet, V.A.; Morrison III, W.R. Prospects for use of biological control of insect and mites for the food industry in North America. Agronomy 2021, 11, 1969. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Population | n | Susceptibility Time Period (min) | KT100 Average (min) | Resistance Coefficient KT100 | Slope ± SE | KT99 (95% CI) | Resistance Coefficient KT99 | χ2 | df | p |
|---|---|---|---|---|---|---|---|---|---|---|
| SoLab | 20 | 10 | 8.1 ± 0.1 a | 0.8 | 13.72 ± 3.99 | 7.90 (6.73–13.07) | 1.0 | 52.99 | 1 | <0.0001 |
| SoRnk | 20 | 10 | 7.6 ± 0.3 a | 0.8 | 8.54 ± 2.59 | 7.81 (6.19–17.32) | 0.7 | 49.56 | 1 | <0.0001 |
| SoBor | 20 | 10 | 7.6 ± 0.2 a | 0.8 | 7.80 ± 2.25 | 7.25 (5.65–15.57) | 0.7 | 32.01 | 1 | <0.0001 |
| SoSml | 20 | 10 | 8.0 ± 0.0 a | 0.8 | 9.64 ± 2.79 | 9.03 (7.25–18.42) | 0.9 | 31.97 | 1 | <0.0001 |
| SoMil | 20 | 10 | 8.0 ± 0.0 a | 0.8 | 14.16 ± 4.63 | 8.90 (7.54–17.46) | 0.9 | 33.82 | 1 | <0.0001 |
| SoZb3 | 20 | 10 | 8.2 ± 0.2 a | 0.8 | 11.08 ± 2.94 | 8.30 (6.88–13.92) | 0.8 | 59.16 | 1 | <0.0001 |
| SoSlc | 20 | 10 | 10.0 ± 0.0 ab | 1.0 | 27.01 ± 9.08 | 8.81 (8.06–12.21) | 8.8 | 57.71 | 1 | <0.0001 |
| SoTrn | 20 | 10 | 10.1 ± 0.1 ab | 1.0 | 10.78 ± 2.86 | 10.63 (8.89–17.57) | 1.1 | 56.79 | 1 | <0.0001 |
| SoOur | 20 | 10 | 10.3 ± 0.3 ab | 1.0 | 7.49 ± 1.74 | 12.04 (9.55–21.08) | 1.2 | 47.89 | 1 | <0.0001 |
| SoIns | 20 | 10 | 10.3 ± 0.2 ab | 1.0 | 14.56 ± 3.94 | 10.74 (9.32–16.25) | 1.1 | 61.06 | 1 | <0.0001 |
| SoTuc | 20 | 10 | 10.6 ± 0.4 ab | 1.1 | 10.53 ± 2.57 | 11.31 (9.50–17.52) | 1.1 | 87.02 | 1 | <0.0001 |
| SoKov | 20 | 10 | 10.6 ± 0.2 ab | 1.1 | 12.38 ± 3.41 | 10.36 (8.81–16.71) | 1.0 | 60.10 | 1 | <0.0001 |
| SoT56 | 20 | 10 | 10.7 ± 0.3 ab | 1.1 | 10.72 ± 2.65 | 11.57 (9.74–18.03) | 1.2 | 68.10 | 1 | <0.0001 |
| SoKas | 20 | 10 | 10.8 ± 0.2 ab | 1.1 | 11.98 ± 2.67 | 11.98 (10.03–19.24) | 1.2 | 53.58 | 1 | <0.0001 |
| SoT59 | 20 | 10 | 10.9 ± 0.2 ab | 1.1 | 7.64 ± 1.79 | 11.97 (9.55–20.75) | 1.2 | 48.34 | 1 | <0.0001 |
| SoSla | 20 | 10 | 12.0 ± 0.5 ab | 1.2 | 11.03 ± 2.62 | 11.99 (10.20–17.76) | 1.2 | 80.48 | 1 | <0.0001 |
| SoCer | 20 | 10 | 12.8 ± 0.3 ab | 1.3 | 12.68 ± 2.95 | 13.43 (11.63–18.98) | 1.3 | 81.90 | 1 | <0.0001 |
| SoUnl | 20 | 10 | 14.4 ± 0.4 b | 1.4 | 6.26 ± 1.18 | 18.88 (14.99–29.97) | 1.9 | 66.39 | 1 | <0.0001 |
| So26 | 20 | 10 | 22.2 ± 1.7 c | 2.2 | 6.23 ± 1.11 | 21.83 (17.60–32.92) | 2.2 | 84.52 | 1 | <0.0001 |
| So27 | 20 | 10 | 22.4 ± 1.5 c | 2.2 | 6.74 ± 1.20 | 21.00 (17.17–30.92) | 2.1 | 88.67 | 1 | <0.0001 |
| So29 | 20 | 10 | 35.5 ± 1.1 d | 3.6 | 7.04 ± 1.30 | 47.10 (36.05–81.30) | 4.7 | 73.56 | 1 | <0.0001 |
| SoEip | 20 | 10 | 69.0 ± 4.5 e | 6.9 | 2.89 ± 0.43 | 111.17 (70.50–248.39) | 11.1 | 77.57 | 1 | <0.0001 |
| Population | n | Susceptibility Time Period (min) | KT100 Average (min) | Resistance Coefficient KT100 | Slope ± SE | KT99 (95% CI) | Resistance Coefficient KT99 | χ2 | df | p |
|---|---|---|---|---|---|---|---|---|---|---|
| OsLab | 20 | 14 | 8.1 ± 0.2 ab | 0.6 | 15.04 ± 4.96 | 7.94 (6.83–14.34) | 1.0 | 54.01 | 1 | <0.0001 |
| OsBlo | 20 | 14 | 6.8 ± 0.2 a | 0.5 | 9.29 ± 3.22 | 6.54 (5.20–17.85) | 0.5 | 35.07 | 1 | <0.0001 |
| Os37 | 20 | 14 | 8.0 ± 0.0 ab | 0.6 | 12.26 ± 3.48 | 8.20 (6.86–14.17) | 1.0 | 37.02 | 1 | <0.0001 |
| Os38 | 20 | 14 | 8.0 ± 0.1 ab | 0.6 | 11.05 ± 3.04 | 8.32 (6.88–14.57) | 1.0 | 48.31 | 1 | <0.0001 |
| OsOur | 20 | 14 | 8.0 ± 0.0 ab | 0.6 | 16.63 ± 6.75 | 7.96 (6.87–24.42) | 1.0 | 38.83 | 1 | <0.0001 |
| OsMal | 20 | 14 | 8.0 ± 0.0 ab | 0.6 | 13.04 ± 4.00 | 8.72 (7.35–15.99) | 0.6 | 34.95 | 1 | <0.0001 |
| OsDuj | 20 | 14 | 8.0 ± 0.0 ab | 0.6 | 7.77 ± 2.35 | 9.54 (7.32–24.30) | 0.7 | 28.89 | 1 | <0.0001 |
| OsKas | 20 | 14 | 8.0 ± 0.0 ab | 0.6 | 18.22 ± 6.98 | 7.23 (6.35–14.93) | 0.5 | 43.94 | 1 | <0.0001 |
| OsZb2 | 20 | 14 | 8.2 ± 0.1 ab | 0.6 | 12.32 ± 3.42 | 8.28 (6.96–13.80) | 1.0 | 50.72 | 1 | <0.0001 |
| OsRcp | 20 | 14 | 8.3± 0.2 abc | 0.6 | 16.61 ± 6.66 | 7.84 (6.80–21.31) | 0.6 | 54.78 | 1 | <0.0001 |
| OsZb1 | 20 | 14 | 8.6 ± 0.2 abc | 0.6 | 13.39 ± 4.06 | 8.53 (7.27–14.65) | 0.6 | 51.53 | 1 | <0.0001 |
| OsChr | 20 | 14 | 9.3 ± 0.3 bcd | 0.7 | 7.40 ± 1.85 | 10.41 (8.19–19.38) | 1.3 | 48.15 | 1 | <0.0001 |
| OsKuc | 20 | 14 | 9.8 ± 0.1 bcd | 0.7 | 12.33 ± 3.55 | 11.19 (9.42–19.55) | 0.8 | 40.39 | 1 | <0.0001 |
| OsPls | 20 | 14 | 10.0 ± 0.0 bcd | 0.7 | 12.57 ± 3.61 | 7.56 (6.33–13.18) | 0.5 | 39.00 | 1 | <0.0001 |
| OsPro | 20 | 14 | 10.0 ± 0.0 bcd | 0.7 | 5.33 ± 1.34 | 13.23 (9.52–31.89) | 1.7 | 29.20 | 1 | <0.0001 |
| OsCho | 20 | 14 | 10.4 ± 0.2 cd | 0.7 | 9.57 ± 2.41 | 10.60 (8.76–17.45) | 1.3 | 54.68 | 1 | <0.0001 |
| OsPol | 20 | 14 | 10.4 ± 0.2 cd | 0.7 | 11.73 ± 3.08 | 11.36 (9.61–18.17) | 0.8 | 56.41 | 1 | <0.0001 |
| OsSla | 20 | 14 | 10.9 ± 0.3 d | 0.8 | 10.79 ± 2.64 | 12.06 (10.16–18.70) | 1.5 | 67.28 | 1 | <0.0001 |
| OsBL1 | 20 | 14 | 13.5 ± 0.5 e | 1.0 | 8.76 ± 1.81 | 14.55 (12.15–21.37) | 1.0 | 82.49 | 1 | <0.0001 |
| OsBus | 20 | 14 | 14.4 ± 0.4 e | 1.0 | 5.52 ± 1.02 | 19.05 (14.77–31.40) | 2.4 | 62.25 | 1 | <0.0001 |
| OsBur | 20 | 14 | 15.5 ± 1.4 e | 1.1 | 6.83 ± 1.29 | 16.24 (13.16–24.57) | 1.2 | 88.70 | 1 | <0.0001 |
| OsIt2 | 20 | 14 | 40.0 ± 1.0 f | 2.9 | 5.50 ± 0.96 | 41.89 (31.73–72.04) | 5.3 | 93.96 | 1 | <0.0001 |
| Population | n | Susceptibility Time Period (min) | KT100 Average (min) | Resistance Coefficient KT100 | Slope ± SE | KT99 (95% CI) | Resistance Coefficient KT99 | χ2 | df | p |
|---|---|---|---|---|---|---|---|---|---|---|
| RdLab | 20 | 15 | 8.7 ± 0.3 a | 0.6 | 5.78 ± 1.36 | 8.79 (6.57–17.40) | 1.0 | 37.40 | 1 | <0.0001 |
| RdKas | 20 | 15 | 8.0 ± 0.0 a | 0.5 | 8.54 ± 2.53 | 10.59 (8.19–25.70) | 0.7 | 26.72 | 1 | <0.0001 |
| RdMis | 20 | 15 | 8.1 ± 0.1 a | 0.5 | 18.70 ± 7.29 | 7.15 (6.31–14.54) | 0.5 | 58.19 | 1 | <0.0001 |
| RdPel | 20 | 15 | 11.8 ± 0.4 ab | 0.8 | 5.92 ± 1.22 | 14.27 (11.07–24.58) | 1.0 | 57.23 | 1 | <0.0001 |
| RdSml | 20 | 15 | 11.4 ± 0.3 ab | 0.8 | 7.53 ± 1.61 | 13.58 (10.91–22.08) | 0.9 | 57.25 | 1 | <0.0001 |
| RdChc | 20 | 15 | 16.5 ± 0.5 abc | 1.1 | 3.92 ± 0.71 | 21.79 (15.81–40.41) | 1.5 | 50.30 | 1 | <0.0001 |
| RdZer | 20 | 15 | 16.5 ± 0.6 abc | 1.1 | 5.86 ± 1.08 | 18.11 (14.38–28.30) | 1.2 | 73.00 | 1 | <0.0001 |
| RdBno | 20 | 15 | 22.4 ± 1.3 abcd | 1.5 | 3.52 ± 0.64 | 27.99 (19.50–57.06) | 1.9 | 44.83 | 1 | <0.0001 |
| RdAus | 20 | 15 | 23.7 ± 3.7 abcd | 1.6 | 4.74 ± 0.79 | 24.62 (18.88–40.04) | 1.6 | 87.11 | 1 | <0.0001 |
| RdPod | 20 | 15 | 26.0 ± 1.9 bcd | 1.7 | 5.01 ± 0.87 | 28.76 (22.00–48.04) | 1.9 | 80.97 | 1 | <0.0001 |
| RdBu2 | 20 | 15 | 27.6 ± 3.0 bcd | 1.8 | 5.03 ± 0.86 | 25.55 (19.78–41.11) | 1.7 | 82.92 | 1 | <0.0001 |
| RdOur | 20 | 15 | 33.0 ± 4.3 cde | 2.2 | 4.38 ± 0.73 | 31.84 (23.85–54.24) | 2.1 | 100.24 | 1 | <0.0001 |
| RdCer | 20 | 15 | 34.5 ± 7.3 de | 2.3 | 3.82 ± 0.62 | 28.77 (21.08–50.33) | 1.9 | 90,25 | 1 | <0.0001 |
| RdKlo | 20 | 15 | 44.3 ± 4.8 e | 3.0 | 4.05 ± 0.64 | 38.00 (27.96–65.65) | 2.5 | 99.22 | 1 | <0.0001 |
| RdBus | 20 | 15 | 61.4 ± 7.0 f | 4.1 | 3.29 ± 0.50 | 51.66 (35.92–97.93) | 3.4 | 93.49 | 1 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aulicky, R.; Stejskal, V.; Frydova, B.; Athanassiou, C. Evaluation of Phosphine Resistance in Populations of Sitophilus oryzae, Oryzaephilus surinamensis and Rhyzopertha dominica in the Czech Republic. Insects 2022, 13, 1162. https://doi.org/10.3390/insects13121162
Aulicky R, Stejskal V, Frydova B, Athanassiou C. Evaluation of Phosphine Resistance in Populations of Sitophilus oryzae, Oryzaephilus surinamensis and Rhyzopertha dominica in the Czech Republic. Insects. 2022; 13(12):1162. https://doi.org/10.3390/insects13121162
Chicago/Turabian StyleAulicky, Radek, Vaclav Stejskal, Barbora Frydova, and Christos Athanassiou. 2022. "Evaluation of Phosphine Resistance in Populations of Sitophilus oryzae, Oryzaephilus surinamensis and Rhyzopertha dominica in the Czech Republic" Insects 13, no. 12: 1162. https://doi.org/10.3390/insects13121162