
Comparative Silk Transcriptomics Illuminates Distinctive Impact of Artificial Selection in Silkworm Modern Breeding
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and RNA Extraction
2.2. Data Collection
2.3. RNA-Seq
2.4. RNA-Seq Data Processing and SNP Calling
2.5. Expression Quantification and Identification of Differential Expressed Genes
2.6. GO and KEGG Enrichment Analysis
2.7. Analysis of the Artificial Signature
3. Results
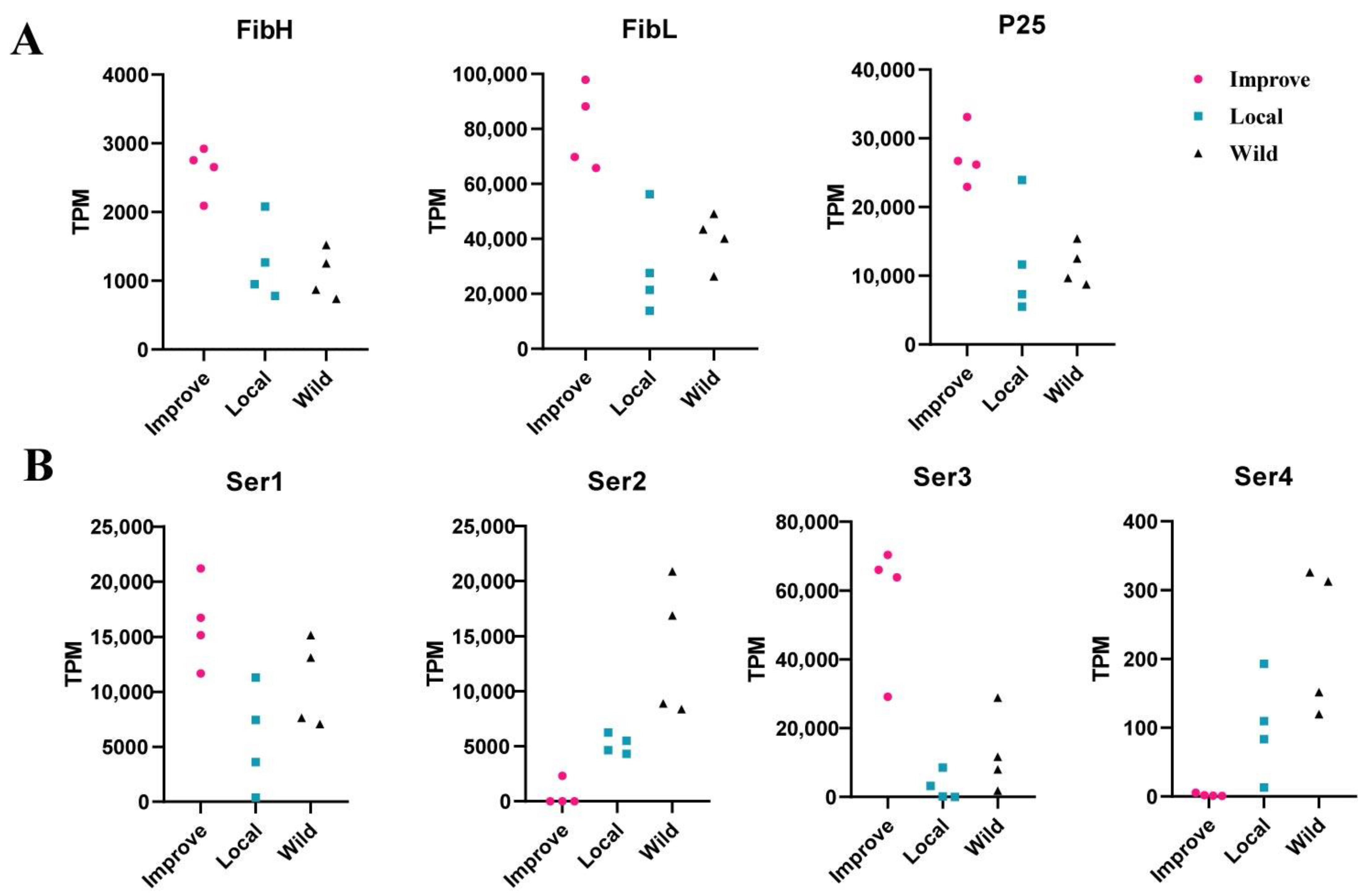
3.1. Silk Cocoon Protein Genes Were Obviously More Highly Expressed in the Silk Gland of Improved Silkworm Strains
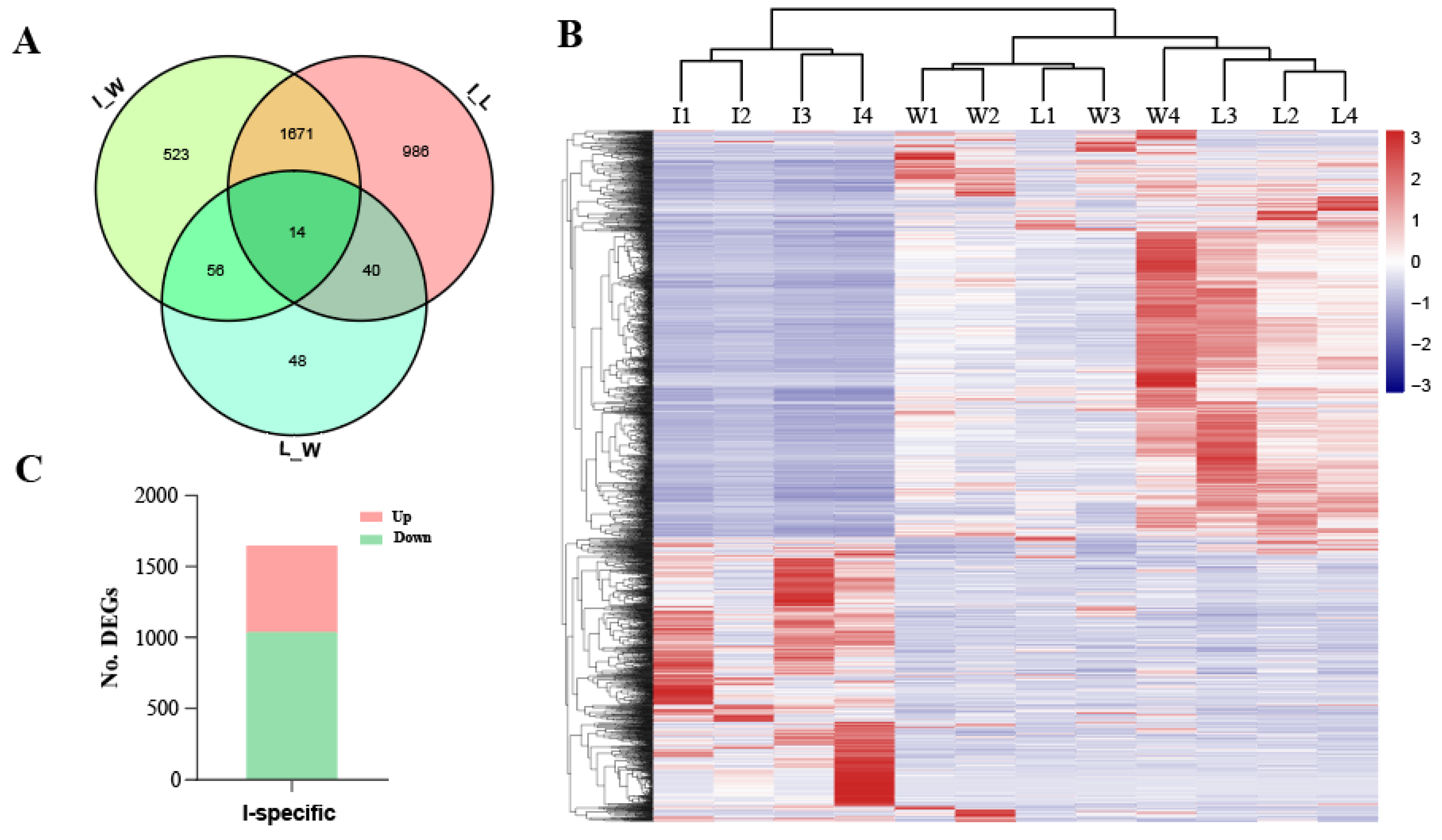
3.2. Improved Silkworms Showed Specific Expression Patterns in the Silk Gland Compared with the Early Domestic and Wild Silkworms
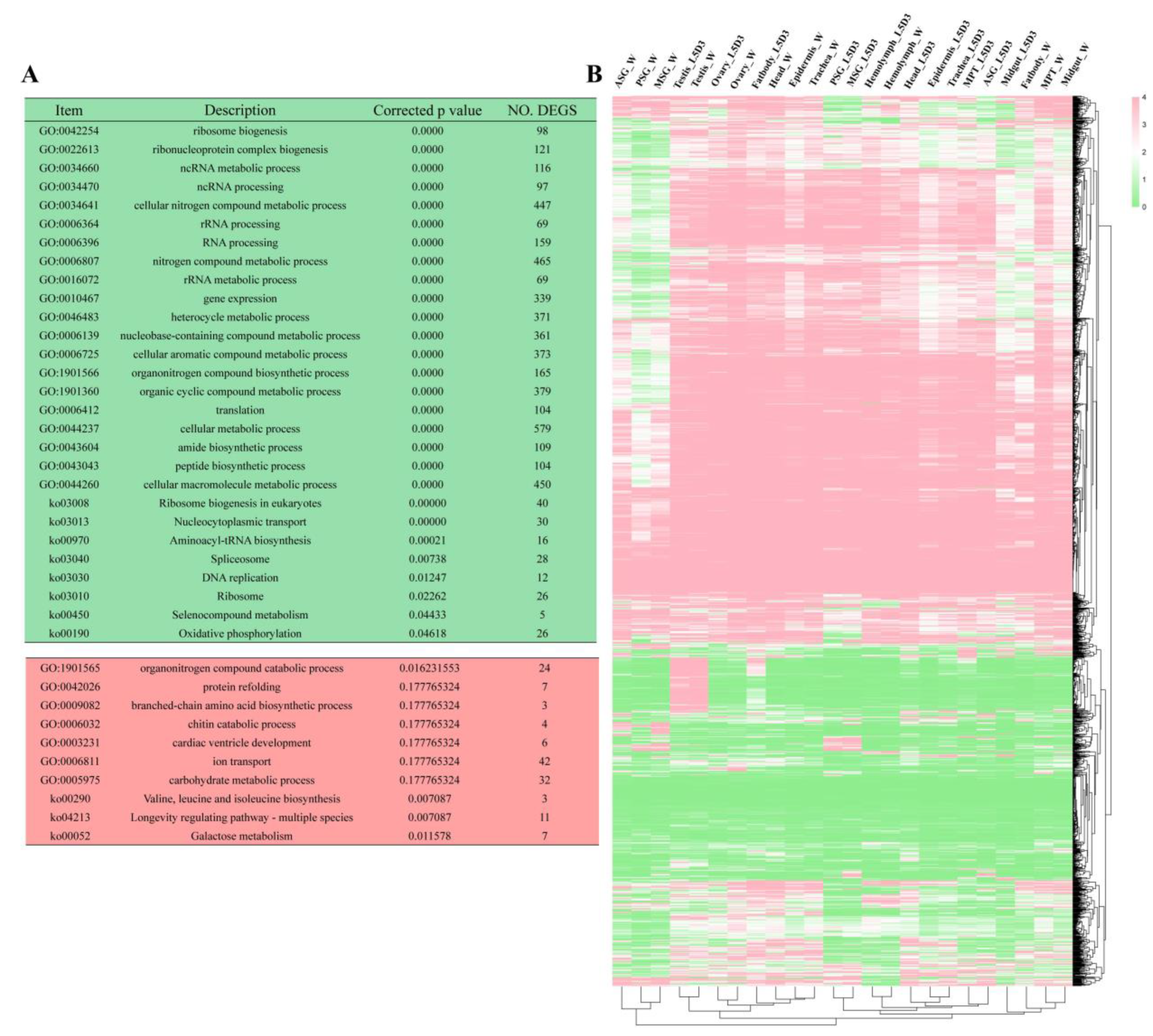
3.3. Up- and Down-Regulated IS-DEGs in the above Enriched Biological Processes and Pathways Showed Different Tissue Expression Patterns
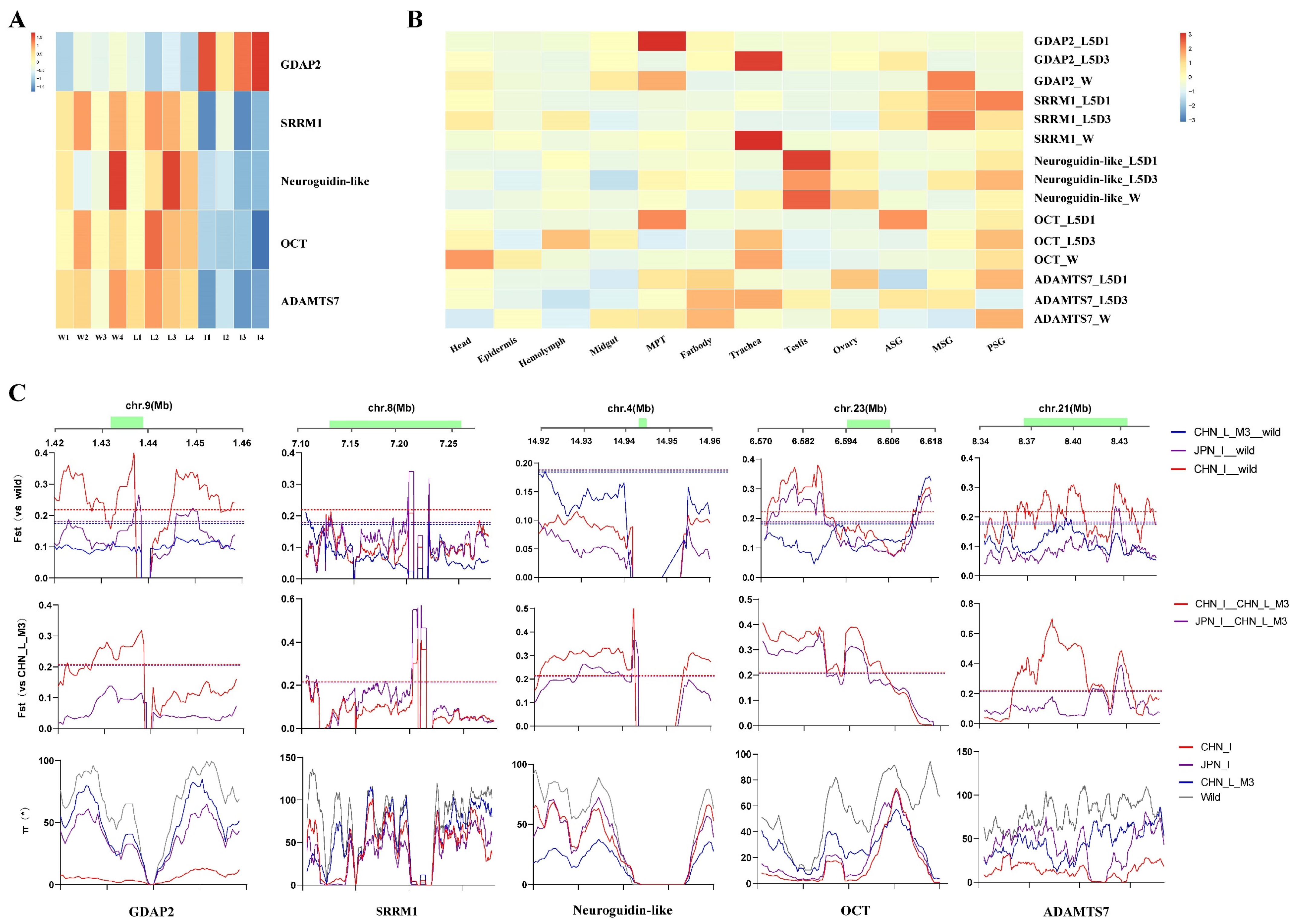
3.4. Five Candidate Improvement Genes Showed Specific Expression Patterns in the Improved Silkworm Strains
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Inoue, S.; Tanaka, K.; Arisaka, F.; Kimura, S.; Ohtomo, K.; Mizuno, S. Silk fibroin of Bombyx mori is secreted, assembling a high molecular mass elementary unit consisting of H-chain, L-chain, and P25, with a 6:6:1 molar ratio. J. Biol. Chem. 2000, 275, 40517–40528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Guo, K.; Zhang, X.; Zhang, T.; Zhang, Y.; Ma, S.; Chang, H.; Tang, M.; An, L.; Xia, Q.; et al. Identification of Bombyx mori sericin 4 protein as a new biological adhesive. Int. J. Biol. Macromol. 2019, 132, 1121–1130. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Sun, Y.; Wu, Y.; Cheng, T.; Liu, C. Bmsage is involved in the determination of cell number in the silk gland of Bombyx mori. Insect Biochem. Mol. Biol. 2019, 113, 103205. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Chen, Y.; Lin, Y.; Xia, Q. Developmental and transcriptomic features characterize defects of silk gland growth and silk production in silkworm naked pupa mutant. Insect Biochem. Mol. Biol. 2019, 111, 103175. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Ma, S.; He, T.; Peng, J.; Zhang, T.; Chen, X.; Wang, X.; Chang, J.; Xia, Q.; Zhao, P. Deep Insight into the Transcriptome of the Single Silk Gland of Bombyx mori. Int. J. Mol. Sci. 2019, 20, 2491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Liu, H.; Shi, Z.; Chen, Q.; Kang, X.; Wang, Y.; Zhao, P. Enhanced silk yield in transgenic silkworm (Bombyx mori) via ectopic expression of BmGT1-L in the posterior silk gland. Insect Mol. Biol. 2020, 29, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.; Guo, Y.; Zhang, Z.; Li, D.; Xuan, Z.; Li, Z.; Dai, F.; Li, Y.; Cheng, D.; Li, R. Complete resequencing of 40 genomes reveals domestication events and genes in silkworm (Bombyx). Science 2009, 326, 433–436. [Google Scholar] [CrossRef] [Green Version]
- Xiang, H.; Liu, X.; Li, M.; Zhu, Y.; Wang, L.; Cui, Y.; Liu, L.; Fang, G.; Qian, H.; Xu, A.; et al. The evolutionary road from wild moth to domestic silkworm. Nat. Ecol. Evol. 2018, 2, 1268–1279. [Google Scholar] [CrossRef]
- Tong, X.; Han, M.J.; Lu, K.; Tai, S.; Liang, S.; Liu, Y.; Hu, H.; Shen, J.; Long, A.; Zhan, C.; et al. High-resolution silkworm pan-genome provides genetic insights into artificial selection and ecological adaptation. Nat. Commun. 2022, 13, 5619. [Google Scholar] [CrossRef]
- Fang, S.M.; Hu, B.L.; Zhou, Q.Z.; Yu, Q.Y.; Zhang, Z. Comparative analysis of the silk gland transcriptomes between the domestic and wild silkworms. BMC Genom. 2015, 16, 60. [Google Scholar] [CrossRef]
- Zhou, Q.Z.; Fu, P.; Li, S.S.; Zhang, C.J.; Yu, Q.Y.; Qiu, C.Z.; Zhang, H.B.; Zhang, Z. A Comparison of Co-expression Networks in Silk Gland Reveals the Causes of Silk Yield Increase During Silkworm Domestication. Front. Genet. 2020, 11, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Qin, S.; Yu, H.; Zhang, J.; Liu, N.; Yu, Y.; Hou, C.; Li, M. Comparative Transcriptome Analysis Reveals Different Silk Yields of Two Silkworm Strains. PLoS ONE 2016, 11, e0155329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; You, Z.; Feng, M.; Che, J.; Zhang, Y.; Qian, Q.; Komatsu, S.; Zhong, B. Analyses of the Molecular Mechanisms Associated with Silk Production in Silkworm by iTRAQ-Based Proteomics and RNA-Sequencing-Based Transcriptomics. J. Proteome Res. 2016, 15, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.H.; You, Z.Y.; Ye, L.P.; Che, J.; Qian, Q.; Nanjo, Y.; Komatsu, S.; Zhong, B.X. Quantitative proteomic and transcriptomic analyses of molecular mechanisms associated with low silk production in silkworm Bombyx mori. J. Proteome Res. 2014, 13, 735–751. [Google Scholar] [CrossRef]
- Meyer, R.S.; Purugganan, M.D. Evolution of crop species: Genetics of domestication and diversification. Nat. Rev. Genet. 2013, 14, 840–852. [Google Scholar] [CrossRef]
- Lu, F.; Wei, Z.; Luo, Y.; Guo, H.; Zhang, G.; Xia, Q.; Wang, Y. SilkDB 3.0: Visualizing and exploring multiple levels of data for silkworm. Nucleic Acids Res. 2020, 48, D749–D755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobin, A.; Gingeras, T.R. Mapping RNA-seq Reads with STAR. Curr. Protoc. Bioinform. 2015, 51, 11.14.11–11.14.19. [Google Scholar] [CrossRef] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. Gigascience 2015, 4, 7. [Google Scholar] [CrossRef]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Dai, F.; Ren, Y.; Liu, H.; Chen, L.; Yang, P.; Liu, Y.; Li, X.; Wang, W.; Xiang, H. Comparative transcriptome analyses on silk glands of six silkmoths imply the genetic basis of silk structure and coloration. BMC Genom. 2015, 16, 203. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Shen, Y.H.; Sun, W.; Kishino, H.; Xiang, Z.H.; Zhang, Z. Nucleotide diversity and selection signature in the domesticated silkworm, Bombyx mori, and wild silkworm, Bombyx mandarina. J. Insect Sci. 2011, 11, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Chen, L.; Tong, X.L.; Hu, H.; Liu, L.Y.; Liu, G.C.; Zhu, Y.N.; Zhao, R.P.; Wang, W.; Dai, F.Y.; et al. Comprehensive silk gland multi-omics comparison illuminates two alternative mechanisms in silkworm heterosis. Zool. Res. 2022, 43, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Y.; Xun, L.J.; Liao, P.F. Main Factors Influencing Silk Production of Silkworm. Shandong Agric. Sci. 2016, 48, 5. (In Chinese) [Google Scholar]
- Ikegaya, M.; Miyazaki, T.; Park, E.Y. Biochemical characterization of Bombyx mori alpha-N-acetylgalactosaminidase belonging to the glycoside hydrolase family 31. Insect Mol. Biol. 2021, 30, 367–378. [Google Scholar] [CrossRef]
- Xu, J.; Morio, A.; Morokuma, D.; Nagata, Y.; Hino, M.; Masuda, A.; Li, Z.; Mon, H.; Kusakabe, T.; Lee, J.M. A functional polypeptide N-acetylgalactosaminyltransferase (PGANT) initiates O-glycosylation in cultured silkworm BmN4 cells. Appl. Microbiol. Biotechnol. 2018, 102, 8783–8797. [Google Scholar] [CrossRef]
- Kajiura, H.; Eguchi, T.; Uchino, K.; Tatematsu, K.I.; Tamura, T.; Sezutsu, H.; Fujiyama, K. Temporal analysis of N-acetylglucosamine extension of N-glycans in the middle silk gland of silkworm Bombyx mori. J. Biosci. Bioeng. 2022, 133, 533–540. [Google Scholar] [CrossRef]
- Kajiura, H.; Miyauchi, R.; Kakudo, A.; Ohashi, T.; Misaki, R.; Fujiyama, K. Bombyx mori beta1,4-N-acetylgalactosaminyltransferase possesses relaxed donor substrate specificity in N-glycan synthesis. Sci. Rep. 2021, 11, 5505. [Google Scholar] [CrossRef]
- Ramakrishnan, B.; Qasba, P.K. Role of a single amino acid in the evolution of glycans of invertebrates and vertebrates. J. Mol. Biol. 2007, 365, 570–576. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zuo, W.; Tong, X.; Han, M.; Gao, R.; Hu, H.; Lu, K.; Luan, Y.; Zhang, B.; Liu, Y.; et al. Whole-genome resequencing reveals loci under selection during silkworm improvement. J. Anim. Breed. Genet. 2021, 138, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Liu, Z.L.; Li, C.C.; Wei, X.M.; Lin, Y.J.; You, L.; Zhu, Z.D.; Deng, H.M.; Feng, Q.L.; Huang, Y.P.; et al. Role of juvenile hormone receptor Methoprene-tolerant 1 in silkworm larval brain development and domestication. Zool. Res. 2021, 42, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Eidhof, I.; Baets, J.; Kamsteeg, E.J.; Deconinck, T.; van Ninhuijs, L.; Martin, J.J.; Schule, R.; Zuchner, S.; De Jonghe, P.; Schenck, A.; et al. GDAP2 mutations implicate susceptibility to cellular stress in a new form of cerebellar ataxia. Brain 2018, 141, 2592–2604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.; Guo, Y.; Wu, M.; Ma, Y.; Jing, W. GDAP2 Overexpression Affects the Development of Neurons and Dysregulates Neuronal Excitatory Synaptic Transmission. Neuroscience 2022, 488, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, X.; Chen, Q.; Hou, Y.; Xia, Q.; Zhao, P. Metabolomics Analysis of the Larval Head of the Silkworm, Bombyx mori. Int. J. Mol. Sci. 2016, 17, 1460. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, K.; Chen, Y.; Chen, L.; Xiang, H. Comparative Silk Transcriptomics Illuminates Distinctive Impact of Artificial Selection in Silkworm Modern Breeding. Insects 2022, 13, 1163. https://doi.org/10.3390/insects13121163
Zhu K, Chen Y, Chen L, Xiang H. Comparative Silk Transcriptomics Illuminates Distinctive Impact of Artificial Selection in Silkworm Modern Breeding. Insects. 2022; 13(12):1163. https://doi.org/10.3390/insects13121163
Chicago/Turabian StyleZhu, Kesen, Yanfei Chen, Lei Chen, and Hui Xiang. 2022. "Comparative Silk Transcriptomics Illuminates Distinctive Impact of Artificial Selection in Silkworm Modern Breeding" Insects 13, no. 12: 1163. https://doi.org/10.3390/insects13121163