Analysis of the Metaphase Chromosome Karyotypes in Imaginal Discs of Aedes communis, Ae. punctor, Ae. intrudens, and Ae. rossicus (Diptera: Culicidae) Mosquitoes

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lacto-Aceto-Orcein Staining
2.2. DAPI Staining
2.3. C-Banding
2.4. Statistical Analysis
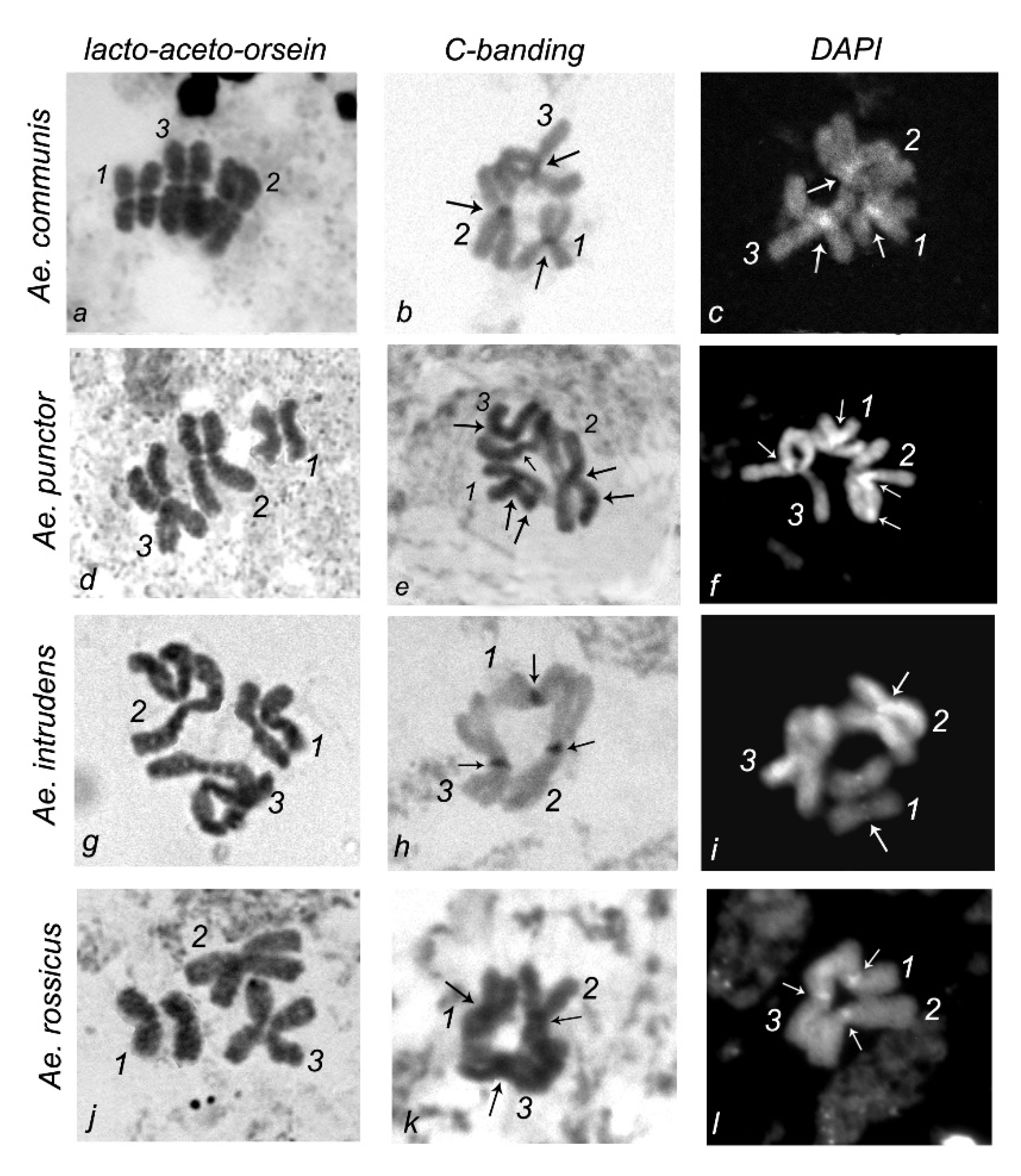
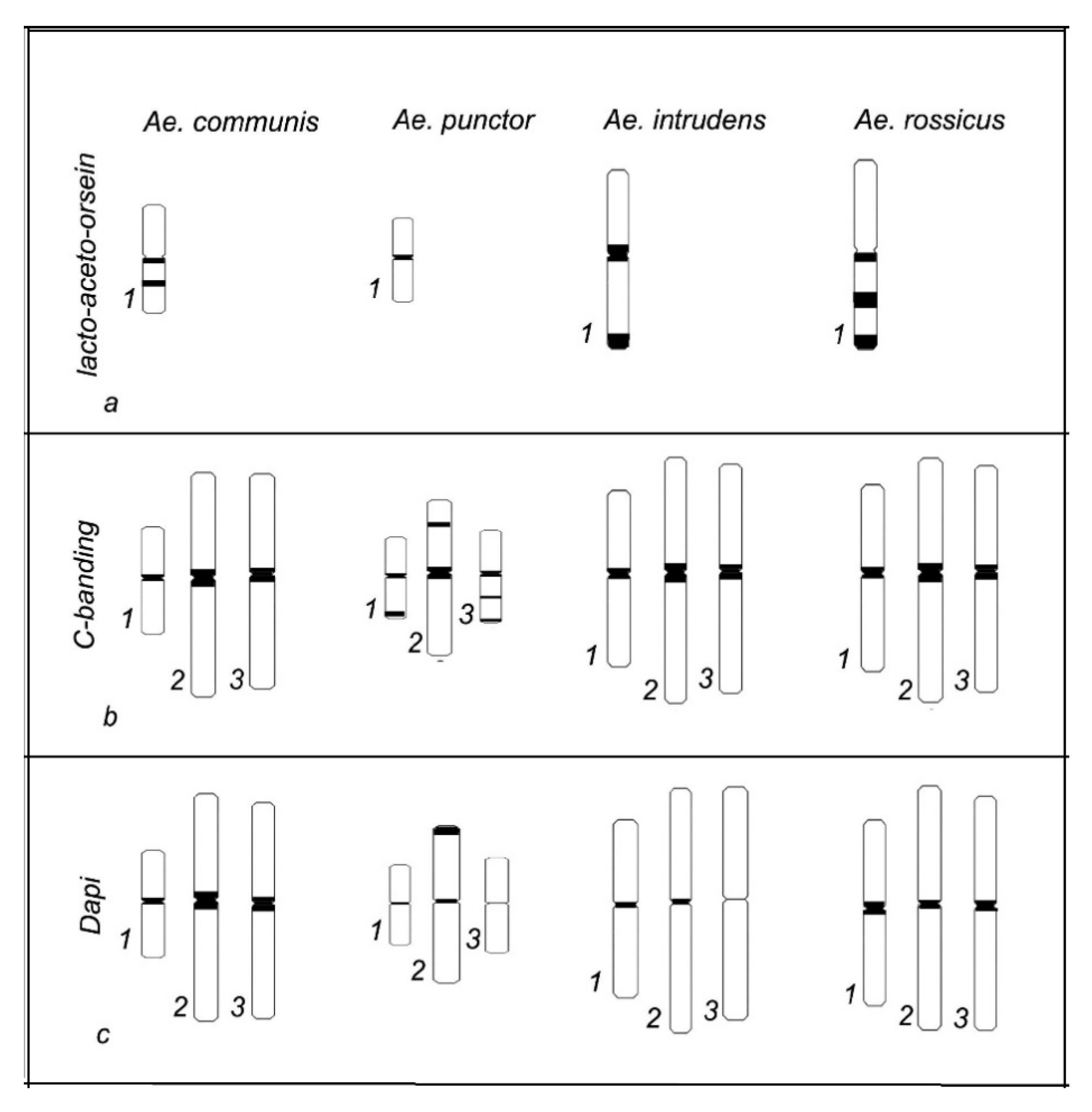
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kononova, Y.V. West Nile Virus in Different Ecosystems of the South of Western Siberia. Author’s Abstract of Candidate of Science (Biology) Dissertation. Vladimir. 2010. Available online: http://www.arriah.ru/sites/default/files/autoreferats/kononova.avtoref.pdf (accessed on 14 December 2019). (In Russian).
- Stock, I. Dengue fever—Not just a tropical infectious disease. Med. Monatsschr. Pharm. 2016, 39, 117–122. [Google Scholar] [PubMed]
- Epelboin, Y.; Talaga, S.; Epelboin, L.; Dusfour, I. Zika virus: An updated review of competent or naturally infected mosquitoes. PLoS Negl. Trop. Dis. 2017, 11, e0005933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matějů, J.; Chanová, M.; Modrý, D.; Mitková, B.; Hrazdilová, K.; Žampachová, V.; Kolářová, L. Dirofilaria repens: Emergence of autochthonous human infections in the Czech Republic (case reports). BMC Infect. Dis. 2016, 16, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atsumi, E.; Matsumoto, H.; Taira, N.; Yohena, T.; Kawasaki, H.; Kawabata, T.; Yoshimi, N. Thirteen cases of pulmonary dirofilariasis in a single institution in Okinawa Island. Virchows Arch. 2019, 475, 335–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaenson, T.G. Diel activity patterns of blood-seeking anthropophilic mosquitoes in central Sweden. Med. Vet. Entomol. 1988, 2, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Gendernalik, A.; Weger-Lucarelli, J.; Garcia Luna, S.M.; Fauver, J.R.; Rückert, C.; Murrieta, R.A.; Bergren, N.; Samaras, D.; Nguyen, C.; Kading, R.C.; et al. American Aedes vexans Mosquitoes are Competent Vectors of Zika Virus. Am. J. Trop. Med. Hyg. 2017, 96, 1338–1340. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Chan, K.; Marek, P.E.; Armstrong, P.M.; Liu, P.; Bova, J.E.; Bernick, J.N.; McMillan, B.E.; Weidlich, B.G.; Paulson, S.L. Cache Valley Virus in Aedes japonicus japonicus Mosquitoes, Appalachian Region, United States. Emerg. Infect. Dis. 2018, 24, 553–557. [Google Scholar] [CrossRef]
- Ciocchetta, S.; Prow, N.A.; Darbro, J.M.; Frentiu, F.D.; Savino, S.; Montarsi, F.; Capelli, G.; Aaskov, J.G.; Devine, G.J. The new European invader Aedes (Finlaya) koreicus: A potential vector of chikungunya virus. Pathog. Glob. Health 2018, 112, 107–114. [Google Scholar] [CrossRef]
- Dharmarajan, G.; Walker, K.D.; Lehmann, T. Variation in Tolerance to Parasites Affects Vectorial Capacity of Natural Asian Tiger Mosquito Populations. Curr. Biol. 2019, 29, 3946–3952. [Google Scholar] [CrossRef]
- Schaffner, F.; Medlock, J.M.; Van, B.W. Public health significance of invasive mosquitoes in Europe. Clin. Microbiol. Infect. 2013, 19, 685–692. [Google Scholar] [CrossRef] [Green Version]
- Medlock, J.M.; Hansford, K.M.; Versteirt, V.; Cull, B.; Kampen, H.; Fontenille, D.; Hendrickx, G.; Zeller, H.; Van, B.W.; Schaffner, F. An entomological review of invasive mosquitoes in Europe. Bull. Entomol. Res. 2015, 105, 637–663. [Google Scholar] [CrossRef] [PubMed]
- Ibañez-Justicia, A.; Gloria-Soria, A.; den Hartog, W.; Dik, M.; Jacobs, F.; Stroo, A. The first detected airline introductions of yellow fever mosquitoes (Aedes aegypti) to Europe, at Schiphol International airport, The Netherlands. Parasites Vectors 2017, 10, 603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metzger, M.E.; Hardstone, Y.M.; Padgett, K.A.; Hu, R.; Kramer, V.L. Detection and Establishment of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) Mosquitoes in California, 2011–2015. J. Med. Entomol. 2017, 54, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Kalan, K.; Buzan, V.E.; Ivović, V. Distribution of two invasive mosquito species in Slovenia in 2013. Parasites Vectors 2014, 7 (Suppl. 1), P9. [Google Scholar] [CrossRef] [Green Version]
- Kalan, K.; Šušnjar, J.; Ivović, V.; Buzan, E. First record of Aedes koreicus (Diptera, Culicidae) in Slovenia. Parasitol. Res. 2017, 116, 2355–2358. [Google Scholar] [CrossRef] [PubMed]
- Montarsi, F.; Martini, F.; Dal Pont, M.; Delai, N.; Ferro Milone, N.; Mazzucato, M.; Soppelsa, F.; Cazzola, L.; Cazzin, S.; Ravagnan, S.; et al. Distribution and habitat characterization of the recently introduced invasive mosquito Aedes koreicus [Hulecoeteomyia koreica], a new potential vector and pest in north-eastern Italy. Parasites Vectors 2013, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Werner, D.; Zielke, D.E.; Kampen, H. First record of Aedes koreicus (Diptera: Culicidae) in Germany. Parasitol. Res. 2016, 115, 1331–1334. [Google Scholar] [CrossRef]
- Akiner, M.; Demirci, B.; Babuadze, G.; Robert, V.; Schaffner, F. Correction: Spread of the Invasive Mosquitoes Aedes aegypti and Aedes albopictus in the Black Sea Region Increases Risk of Chikungunya, Dengue, and Zika Outbreaks in Europe. PLoS Negl. Trop. Dis. 2016, 10, e0004764. [Google Scholar] [CrossRef] [Green Version]
- Blomgren, E.; Hesson, J.C.; Schäfer, M.L.; Lundström, J.O. Pest occurrence of Aedes rossicus close to the Arctic Circle in northern Sweden. J. Vector Ecol. 2018, 43, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Prokof’eva-Bel’govskaya, A.A. Heterochromatin Regions of Chromosomes; Nauka: Moscow, Russia, 1986. (In Russian) [Google Scholar]
- Srivastava, R.; Mishra, N. Understanding heterochromatin characterization through chromosome. J. Biol. Sci. Med. 2015, 1, 11–20. [Google Scholar]
- Chirino, M.G.; Rossi, L.F.; Bressa, M.J.; Luaces, J.P.; Merani, M.S. Comparative study of mitotic chromosomes in two blowflies, Lucilia sericata and L. cluvia (Diptera, Calliphoridae), by C- and G-like banding patterns and rRNA loci, and implications for karyotype evolution. Comp. Cytogen. 2015, 9, 103–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, U.R.; Bajpai, N.; Kurahashi, H.; Tewari, R.R. Metaphase karyotypes of four species of Calliphoridae (Diptera). Chromosome Sci. 2010, 13, 49–52. [Google Scholar]
- Baimai, V. Heterochromatin accumulation and karyotypic evolution in some dipteran insects. Zool. Stud. 1998, 37, 75–88. [Google Scholar]
- Marchi, A.; Mezzanotte, R. Inter- and intraspecific heterochromatin variation detected by restriction endonuclease digestion in two sibling species of the Anopheles maculipennis complex. Genet. Soc. Great Br. Hered. 1990, 65, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasserlauf, I.E.; Alekseeva, S.S.; Andreeva, Y.V.; Sibataev, A.K.; Stegniy, V.N. A comparative analysis of the metaphase karyotypes of Aedes excrucians, Ae. behningi, and Ae. euedes (Diptera: Culicidae) imaginal disсs. J. Vector Ecol. 2018, 43, 245–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreeva, Y.V.; Khrabrova, N.V.; Simakova, A.V.; Sibataeva, A.M.; Sibataev, A.K. Species diversity of blood-sucking mosquitoes (Diptera: Culicidae) in Tomsk Region. Int. J. Environ. Stud. 2017, 74, 782–789. [Google Scholar] [CrossRef]
- Gutsevich, A.V.; Monchadskii, A.S.; Shtakel’berg, A.A. Mosquitoes of the family Culicidae. In The Fauna of the USSR. Dipteran Insects; Nauka, USSR: Leningrad, Russia, 1970; Volume 3, p. 384. [Google Scholar]
- Gutsevich, A.V.; Dubitskii, A.M. New mosquito species in the fauna of the Soviet Union. Parazitol. Sbornik 1981, 30, 97–165. [Google Scholar]
- Becker, N.; Petric, D.; Zgomba, M.; Boase, C.; Madon, M.B.; Dahl, C.; Kaiser, A. Mosquitoes and Their Control; Springer: Berlin/Heidelberg, Germany, 2010; Volume 1, p. 594. [Google Scholar] [CrossRef]
- Kabanova, V.M.; Kartashova, N.N. Karyotypes of blood-sucking mosquitoes of the genus Aedes (Culicidae, Diptera). Genetika 1972, 8, 47–51. [Google Scholar]
- Saifitdinova, A.F. Two-dimensional fluorescence microscopy for analysis of biological samples. In Study Guide; SOLO: St. Petersburg, Russia, 2008; pp. 1–72. [Google Scholar]
- McDonald, P.T.; Rai, K.S. Correlation of linkage group with chromosomes in the mosquito. Aedes Aegypti. Genet. 1970, 66, 475–485. [Google Scholar]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric possions on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Trotta, E.; D’Ambrosio, E.; Ravagnan, G.; Pacil, M. Evidence for DAPI intercalation in CG sites of DNA oligomer [d(CGACGTCG)]2: A 1H NMR study. Nucleic Acids Res. 1995, 23, 1333–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
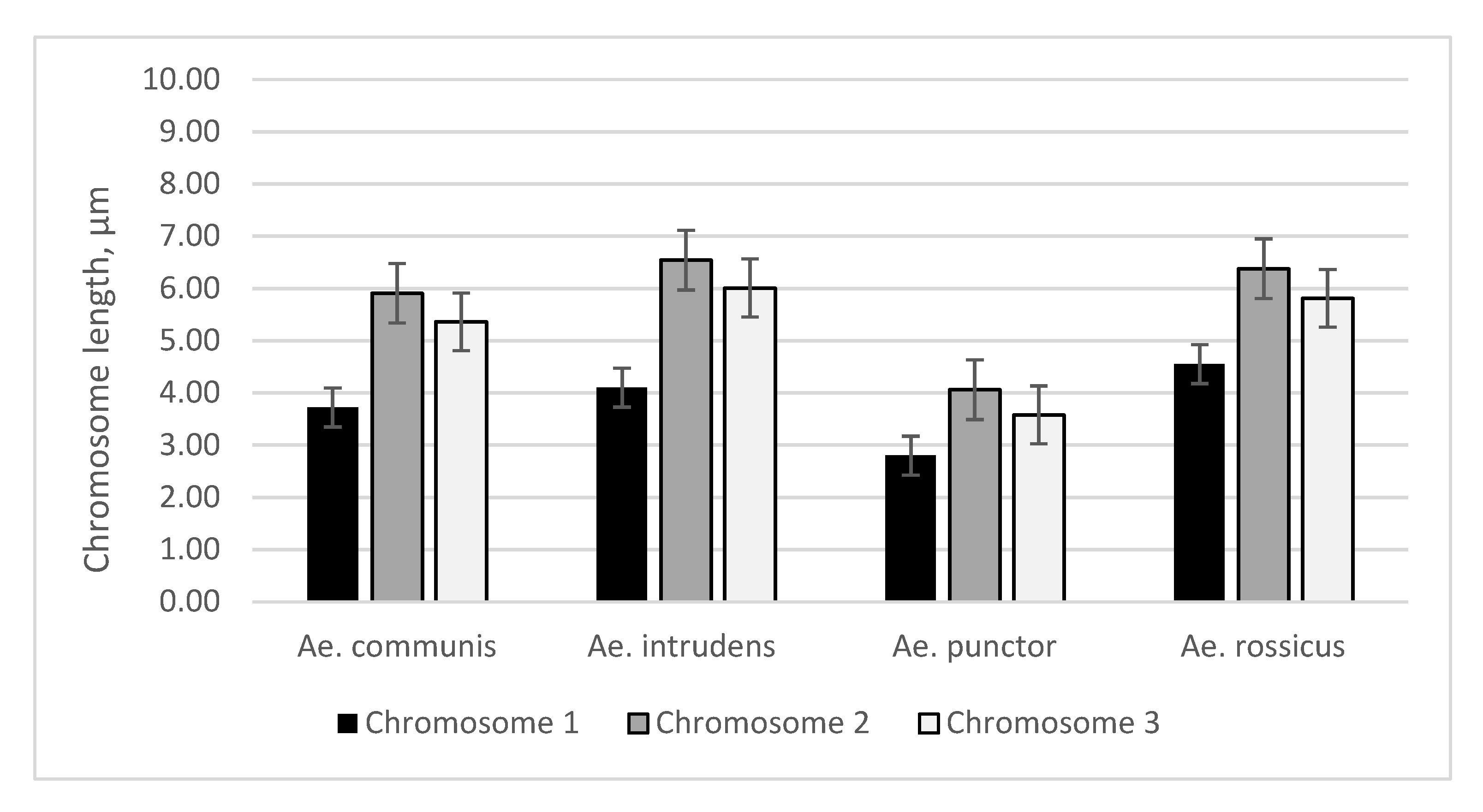
| Species | Length of the Chromosomes ± Standard Error of the Mean Value, μm | ||
|---|---|---|---|
| Chromosome 1 | Chromosome 2 | Chromosome 3 | |
| Ae. communis | 3.72 ± 0.16 | 5.91 ± 0.32 | 5.36 ± 0.21 |
| Ae. punctor | 2.8 ± 0.10 | 4.06 ± 0.11 | 3.58 ± 0.14 |
| Ae. intrudens | 4.1 ± 0.10 | 6.54 ± 0.19 | 6.01 ± 0.14 |
| Ae. rossicus | 4.55 ± 0.11 | 6.38 ± 0.22 | 5.81 ± 0.20 |
| Species | Chromosomes | |||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | ||||
| Lr, % | Jc, % | Lr, % | Jc, % | Lr, % | Jc, % | |
| Ae. communis | 24 | 48 | 40 | 45 | 35 | 47 |
| Ae. punctor | 27 | 51 | 39 | 54 | 34 | 51 |
| Ae. intrudens | 25 | 48 | 40 | 53 | 36 | 48 |
| Ae. rossicus | 27 | 49 | 38 | 45 | 35 | 45 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alekseeva, S.S.; Andreeva, Y.V.; Wasserlauf, I.E.; Sibataev, A.K.; Stegniy, V.N. Analysis of the Metaphase Chromosome Karyotypes in Imaginal Discs of Aedes communis, Ae. punctor, Ae. intrudens, and Ae. rossicus (Diptera: Culicidae) Mosquitoes. Insects 2020, 11, 63. https://doi.org/10.3390/insects11010063
Alekseeva SS, Andreeva YV, Wasserlauf IE, Sibataev AK, Stegniy VN. Analysis of the Metaphase Chromosome Karyotypes in Imaginal Discs of Aedes communis, Ae. punctor, Ae. intrudens, and Ae. rossicus (Diptera: Culicidae) Mosquitoes. Insects. 2020; 11(1):63. https://doi.org/10.3390/insects11010063
Chicago/Turabian StyleAlekseeva, Svetlana S., Yulia V. Andreeva, Irina E. Wasserlauf, Anuarbek K. Sibataev, and Vladimir N. Stegniy. 2020. "Analysis of the Metaphase Chromosome Karyotypes in Imaginal Discs of Aedes communis, Ae. punctor, Ae. intrudens, and Ae. rossicus (Diptera: Culicidae) Mosquitoes" Insects 11, no. 1: 63. https://doi.org/10.3390/insects11010063