Drosophila melanogaster Responses against Entomopathogenic Nematodes: Focus on Hemolymph Clots

Abstract
:1. Introduction
1.1. Two Key Immune Pathways Are Dispensable in a Drosophila EPN Model
1.2. A Search for Alternative Immune Reactions against EPNs
2. Hemolymph Coagulation in Non-Insect Arthropods
3. Hemolymph Coagulation in Insects
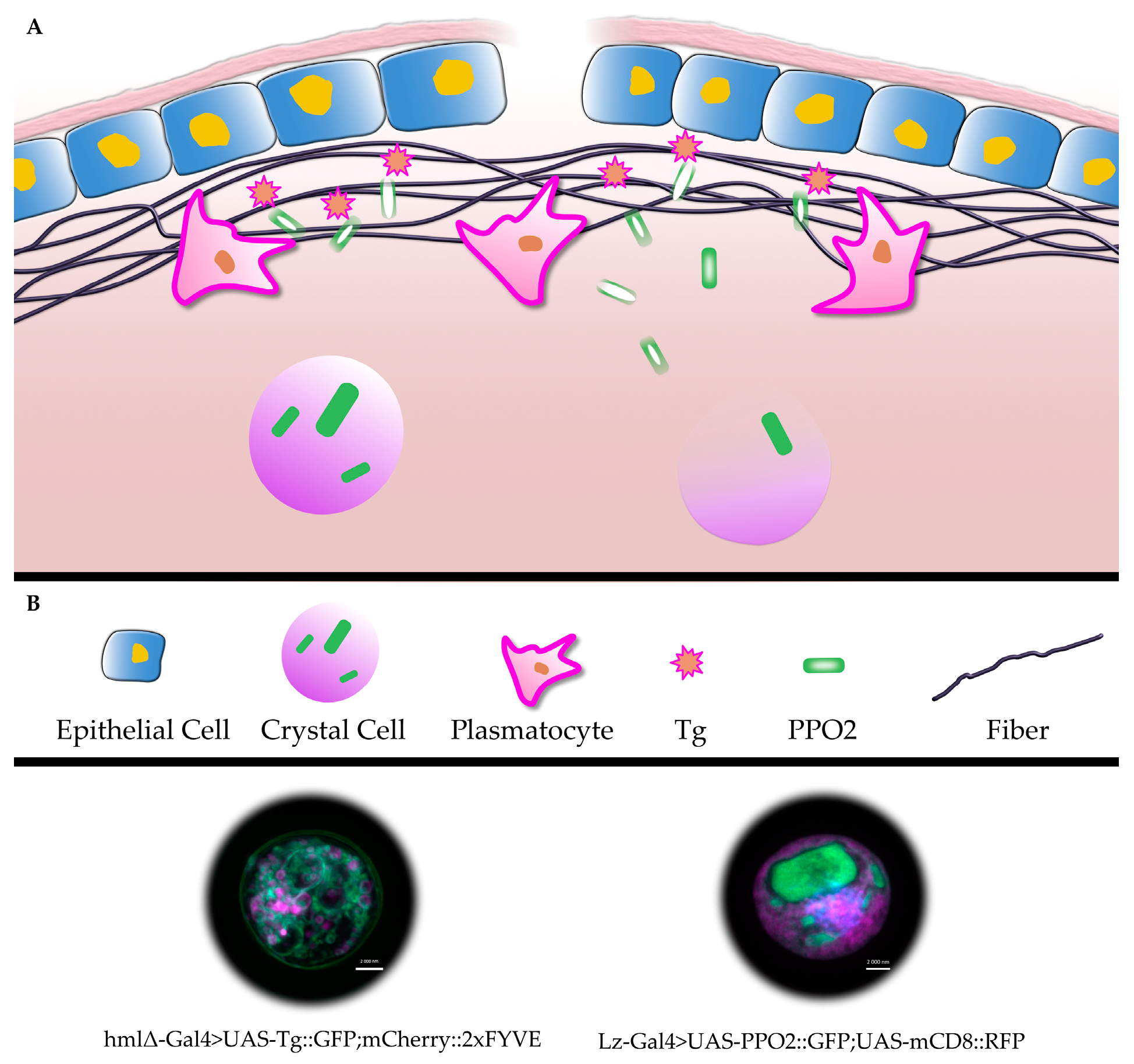
4. Non-Classical Secretion (NCS) of Drosophila Coagulation Factors
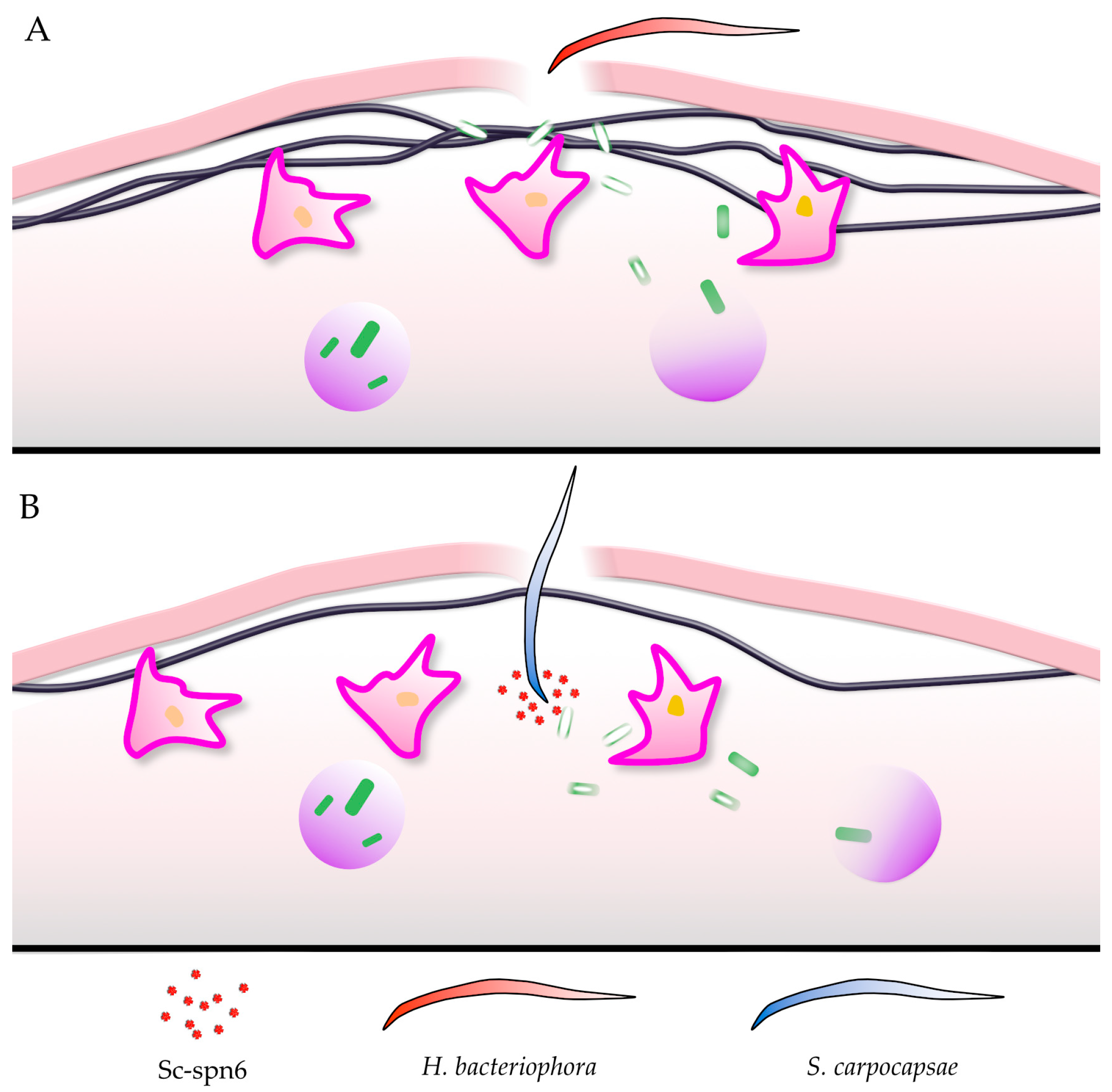
5. Drosophila Clots Protect against EPNs
6. Other Immune Factors—Eicosanoids as Mediators of Anti-EPN Responses
7. Moving beyond Insects—How Evolution Shapes Clot Formation
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Gene | Effect on EPNs | EPN Used | References |
|---|---|---|---|
| imd | - | Heterorhabditis/Photorhabdus | [2] |
| Tl | - | Heterorhabditis/Photorhabdus | [2] |
| Transglutaminase | + | Heterorhabditis/Photorhabdus | [38] |
| Tiggrin | - | Heterorhabditis/Photorhabdus | [46] |
| Fondue | + (Fb) | Heterorhabditis/Photorhabdus | [46] |
| Eig71Ee | + (Hc) | Heterorhabditis/Photorhabdus | [46] |
| Fbp1 | - | Heterorhabditis/Photorhabdus | [46] |
| Hemomucin | - | Heterorhabditis/Photorhabdus | [46] |
| Hemolectin | - | Heterorhabditis/Photorhabdus | [38] |
| GNBP-like 3 | + (Fb) | Heterorhabditis/Photorhabdus | [36] |
| PGRP-LF | + | Heterorhabditis/Photorhabdus | [36] |
| TEP3 | + | Heterorhabditis/Photorhabdus | [36] |
| Glutactin | + (Hc) | Heterorhabditis/Photorhabdus | [36] |
| Idgf3 | + | Heterorhabditis/Photorhabdus | [37] |
| CG 14507 (PLA2) | + | Heterorhabditis/Photorhabdus | [46] |
| PPO1 | + | Steinernema/Xenorhabdus | [9] |
| PPO2 | + | Steinernema/Xenorhabdus | [9] |
| PPO3 | + | Steinernema/Xenorhabdus | [9] |
| Bc | - | Heterorhabditis/Photorhabdus | [38] |
| Sp7 (PPO activation) | - | Heterorhabditis/Photorhabdus | [38] |
References
- Castillo, J.C.; Reynolds, S.E.; Eleftherianos, I. Insect immune responses to nematode parasites. Trends Parasitol. 2011, 27, 537–547. [Google Scholar] [CrossRef]
- Hallem, E.A.; Rengarajan, M.; Ciche, T.A.; Sternberg, P.W. Nematodes, bacteria, and flies: A tripartite model for nematode parasitism. Curr. Biol. 2007, 17, 898–904. [Google Scholar] [CrossRef] [Green Version]
- Peters, A.; Ehlers, R.U. Encapsulation of the entomopathogenic nematode Steinernema feltiae in Tipula oleracea. J. Invertebr. Pathol. 1997, 69, 218–222. [Google Scholar] [CrossRef]
- Ebrahimi, L.; Niknam, G.; Dunphy, G.B. Hemocyte responses of the Colorado potato beetle, Leptinotarsa decemlineata, and the greater wax moth, Galleria mellonella, to the entomopathogenic nematodes, Steinernema feltiae and Heterorhabditis bacteriophora. J. Insect. Sci. 2011, 11, 75. [Google Scholar] [CrossRef] [Green Version]
- Anderl, I.; Vesala, L.; Ihalainen, T.O.; Vanha-Aho, L.M.; Ando, I.; Ramet, M.; Hultmark, D. Transdifferentiation and Proliferation in Two Distinct Hemocyte Lineages in Drosophila melanogaster Larvae after Wasp Infection. PLoS Pathog. 2016, 12, e1005746. [Google Scholar] [CrossRef] [PubMed]
- Binda-Rossetti, S.; Mastore, M.; Protasoni, M.; Brivio, M.F. Effects of an entomopathogen nematode on the immune response of the insect pest red palm weevil: Focus on the host antimicrobial response. J. Invertebr. Pathol. 2016, 133, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Pena, J.M.; Carrillo, M.A.; Hallem, E.A. Variation in the susceptibility of Drosophila to different entomopathogenic nematodes. Infect. Immun. 2015, 83, 1130–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitten, M.M.A.; Coates, C.J. Re-evaluation of insect melanogenesis research: Views from the dark side. Pigment Cell Melanoma Res. 2017, 30, 386–401. [Google Scholar] [CrossRef]
- Cooper, D.; Wuebbolt, C.; Heryanto, C.; Eleftherianos, I. The prophenoloxidase system in Drosophila participates in the anti-nematode immune response. Mol. Immunol. 2019, 109, 88–98. [Google Scholar] [CrossRef]
- Brivio, M.F.; Mastore, M.; Nappi, A.J. A pathogenic parasite interferes with phagocytosis of insect immunocompetent cells. Dev. Comp. Immunol. 2010, 34, 991–998. [Google Scholar] [CrossRef]
- Honti, V.; Csordas, G.; Kurucz, E.; Markus, R.; Ando, I. The cell-mediated immunity of Drosophila melanogaster: Hemocyte lineages, immune compartments, microanatomy and regulation. Dev. Comp. Immunol. 2014, 42, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Sadekuzzaman, M.; Park, Y.; Lee, S.; Kim, K.; Jung, J.K.; Kim, Y. An entomopathogenic bacterium, Xenorhabdus hominickii ANU101, produces oxindole and suppresses host insect immune response by inhibiting eicosanoid biosynthesis. J. Invertebr. Pathol. 2017, 145, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, G.; Clarke, G.; Kavanagh, K. Characterisation of the cellular and proteomic response of Galleria mellonella larvae to the development of invasive aspergillosis. BMC Microbiol. 2018, 18, 63. [Google Scholar] [CrossRef] [PubMed]
- Ratcliffe, N.A.; Rowley, A.F. Role of hemocytes in defense against biological agents. In Insect Hemocytes; Gupta, A.P., Ed.; Cambridge University Press: Cambridge, UK, 1979; pp. 331–414. [Google Scholar]
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [Green Version]
- Galko, M.J.; Krasnow, M.A. Cellular and genetic analysis of wound healing in Drosophila larvae. PLoS Biol. 2004, 2, E239. [Google Scholar] [CrossRef] [Green Version]
- Lesch, C.; Jo, J.; Wu, Y.; Fish, G.S.; Galko, M.J. A targeted UAS-RNAi screen in Drosophila larvae identifies wound closure genes regulating distinct cellular processes. Genetics 2010, 186, 943–957. [Google Scholar] [CrossRef] [Green Version]
- Theopold, U.; Krautz, R.; Dushay, M.S. The Drosophila clotting system and its messages for mammals. Dev. Comp. Immunol. 2014, 42, 42–46. [Google Scholar] [CrossRef]
- Krzemien, J.; Dubois, L.; Makki, R.; Meister, M.; Vincent, A.; Crozatier, M. Control of blood cell homeostasis in Drosophila larvae by the posterior signalling centre. Nature 2007, 446, 325–328. [Google Scholar] [CrossRef]
- Markus, R.; Laurinyecz, B.; Kurucz, E.; Honti, V.; Bajusz, I.; Sipos, B.; Somogyi, K.; Kronhamn, J.; Hultmark, D.; Ando, I. Sessile hemocytes as a hematopoietic compartment in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2009, 106, 4805–4809. [Google Scholar] [CrossRef] [Green Version]
- Dudzic, J.P.; Kondo, S.; Ueda, R.; Bergman, C.M.; Lemaitre, B. Drosophila innate immunity: Regional and functional specialization of prophenoloxidases. BMC Biol. 2015, 13, 81. [Google Scholar] [CrossRef] [Green Version]
- Irving, P.; Ubeda, J.M.; Doucet, D.; Troxler, L.; Lagueux, M.; Zachary, D.; Hoffmann, J.A.; Hetru, C.; Meister, M. New insights into Drosophila larval haemocyte functions through genome-wide analysis. Cell Microbiol. 2005, 7, 335–350. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, C. Hemolymph coagulation. In The Physiology of Insecta, 2nd ed.; Rockstein, M., Ed.; Academic Press: New York, NY, USA, 1974; Volume 5, pp. 309–360. [Google Scholar]
- Osaki, T.; Kawabata, S. Structure and function of coagulogen, a clottable protein in horseshoe crabs. Cell Mol. Life Sci. 2004, 61, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Iwanaga, S. The limulus clotting reaction. Curr. Opin. Immunol. 1993, 5, 74–82. [Google Scholar] [CrossRef]
- Osaki, T.; Okino, N.; Tokunaga, F.; Iwanaga, S.; Kawabata, S. Proline-rich cell surface antigens of horseshoe crab hemocytes are substrates for protein cross-linking with a clotting protein coagulin. J. Biol. Chem. 2002, 277, 40084–40090. [Google Scholar] [CrossRef]
- Iwanaga, S.; Muta, T.; Shigenaga, T.; Miura, Y.; Seki, N.; Saito, T.; Kawabata, S. Role of hemocyte-derived granular components in invertebrate defense. Ann. N. Y. Acad. Sci. 1994, 712, 102–116. [Google Scholar] [CrossRef]
- Cerenius, L.; Soderhall, K. Coagulation in invertebrates. J. Innate Immun. 2011, 3, 3–8. [Google Scholar] [CrossRef]
- Perdomo-Morales, R.; Montero-Alejo, V.; Perera, E. The clotting system in decapod crustaceans: History, current knowledge and what we need to know beyond the models. Fish Shellfish Immunol. 2019, 84, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.; Wang, R.; van Antwerpen, R.; Sottrup-Jensen, L.; Söderhäll, K. The crayfish plasma clotting protein: A vitellogenin-related protein responsible for clot formation in crustacean blood. Proc. Natl. Acad. Sci. USA 1999, 96, 1965–1970. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, G.; Garvey, A.; Croke, M.; Kavanagh, K. Innate humoral immune defences in mammals and insects: The same, with differences? Virulence 2018, 9, 1625–1639. [Google Scholar] [CrossRef] [Green Version]
- Theopold, U.; Li, D.; Fabbri, M.; Scherfer, C.; Schmidt, O. The coagulation of insect hemolymph. Cell. Mol. Life Sci. 2002, 59, 363–372. [Google Scholar] [CrossRef]
- Scherfer, C.; Karlsson, C.; Loseva, O.; Bidla, G.; Goto, A.; Havemann, J.; Dushay, M.S.; Theopold, U. Isolation and Characterization of Hemolymph Clotting Factors in Drosophila melanogaster by a Pullout Method. Curr. Biol. 2004, 14, 625–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, C.; Korayem, A.M.; Scherfer, C.; Loseva, O.; Dushay, M.S.; Theopold, U. Proteomic analysis of the Drosophila larval hemolymph clot. J. Biol. Chem. 2004, 279, 52033–52041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theopold, U.; Samakovlis, C.; Erdjument-Bromage, H.; Dillon, N.; Axelsson, B.; Schmidt, O.; Tempst, P.; Hultmark, D. Helix pomatia lectin, an inducer of Drosophila immune response binds to hemomucin, a novel surface mucin. J. Biol. Chem. 1996, 271, 12708–12715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arefin, B.; Kucerova, L.; Dobes, P.; Markus, R.; Strnad, H.; Wang, Z.; Hyrsl, P.; Zurovec, M.; Theopold, U. Genome-wide transcriptional analysis of Drosophila larvae infected by entomopathogenic nematodes shows involvement of complement, recognition and extracellular matrix proteins. J. Innate Immun. 2014, 6, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Kucerova, L.; Broz, V.; Arefin, B.; Maaroufi, H.O.; Hurychova, J.; Strnad, H.; Zurovec, M.; Theopold, U. The Drosophila Chitinase-Like Protein IDGF3 Is Involved in Protection against Nematodes and in Wound Healing. J. Innate Immun. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Wilhelmsson, C.; Hyrsl, P.; Loof, T.G.; Dobes, P.; Klupp, M.; Loseva, O.; Morgelin, M.; Ikle, J.; Cripps, R.M.; et al. Pathogen entrapment by transglutaminase-a conserved early innate immune mechanism. PLoS Pathog. 2010, 6, e1000763. [Google Scholar] [CrossRef] [Green Version]
- Loof, T.G.; Morgelin, M.; Johansson, L.; Oehmcke, S.; Olin, A.I.; Dickneite, G.; Norrby-Teglund, A.; Theopold, U.; Herwald, H. Coagulation, an ancestral serine protease cascade, exerts a novel function in early immune defense. Blood 2011, 118, 2589–2598. [Google Scholar] [CrossRef]
- Myers, A.L.; Harris, C.M.; Choe, K.M.; Brennan, C.A. Inflammatory production of reactive oxygen species by Drosophila hemocytes activates cellular immune defenses. Biochem. Biophys. Res. Commun. 2018, 505, 726–732. [Google Scholar] [CrossRef]
- Schmid, M.R.; Dziedziech, A.; Arefin, B.; Kienzle, T.; Wang, Z.; Akhter, M.; Berka, J.; Theopold, U. Insect hemolymph coagulation: Kinetics of classically and non-classically secreted clotting factors. Insect. Biochem. Mol. Biol. 2019, 109, 63–71. [Google Scholar] [CrossRef]
- Dziedziech, A.; Schmid, M.; Arefin, B.; Kienzle, T.; Krautz, R.; Theopold, U. Data on Drosophila clots and hemocyte morphologies using GFP-tagged secretory proteins: Prophenoloxidase and transglutaminase. Data Brief 2019, 25, 104229. [Google Scholar] [CrossRef]
- Lindgren, M.; Riazi, R.; Lesch, C.; Wilhelmsson, C.; Theopold, U.; Dushay, M.S. Fondue and transglutaminase in the Drosophila larval clot. J. Insect. Physiol. 2008, 54, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Hadano, J.; Kawasaki, D.; Dong, X.; Kawabata, S.I. Drosophila TG-A transglutaminase is secreted via an unconventional Golgi-independent mechanism involving exosomes and two types of fatty acylations. J. Biol. Chem. 2017, 292, 10723–10734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibata, T.; Kawabata, S.I. Pluripotency and a secretion mechanism of Drosophila transglutaminase. J. Biochem. 2018, 163, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Hyrsl, P.; Dobes, P.; Wang, Z.; Hauling, T.; Wilhelmsson, C.; Theopold, U. Clotting factors and eicosanoids protect against nematode infections. J. Innate Immun. 2011, 3, 65–70. [Google Scholar] [CrossRef]
- Bajzek, C.; Rice, A.M.; Andreazza, S.; Dushay, M.S. Coagulation and survival in Drosophila melanogaster fondue mutants. J. Insect. Physiol. 2012, 58, 1376–1381. [Google Scholar] [CrossRef]
- Scherfer, C.; Qazi, M.R.; Takahashi, K.; Ueda, R.; Dushay, M.S.; Theopold, U.; Lemaitre, B. The Toll immune-regulated Drosophila protein Fondue is involved in hemolymph clotting and puparium formation. Dev. Biol. 2006, 295, 156–163. [Google Scholar] [CrossRef] [Green Version]
- Arneth, B. Coevolution of the coagulation and immune systems. Inflamm. Res. 2019, 68, 117–123. [Google Scholar] [CrossRef]
- Papareddy, P.; Kasetty, G.; Alyafei, S.; Smeds, E.; Salo-Ahen, O.M.H.; Hansson, S.R.; Egesten, A.; Herwald, H. An ecoimmunological approach to study evolutionary and ancient links between coagulation, complement and Innate immunity. Virulence 2018, 9, 724–737. [Google Scholar] [CrossRef] [Green Version]
- Shokal, U.; Eleftherianos, I. Evolution and Function of Thioester-Containing Proteins and the Complement System in the Innate Immune Response. Front. Immunol. 2017, 8, 759. [Google Scholar] [CrossRef]
- Shokal, U.; Kopydlowski, H.; Harsh, S.; Eleftherianos, I. Thioester-Containing Proteins 2 and 4 Affect the Metabolic Activity and Inflammation Response in Drosophila. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [Green Version]
- Theopold, U.; Schmid, M. Thioester-containing proteins: At the crossroads of immune effector mechanisms. Virulence 2017, 8, 1468–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toubarro, D.; Avila, M.M.; Hao, Y.; Balasubramanian, N.; Jing, Y.; Montiel, R.; Faria, T.Q.; Brito, R.M.; Simoes, N. A serpin released by an entomopathogen impairs clot formation in insect defense system. PLoS ONE 2013, 8, e69161. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.; Kim, Y. Prostaglandins and Other Eicosanoids in Insects: Biosynthesis and Biological Actions. Front. Physiol. 2018, 9, 1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Ji, D.; Cho, S.; Park, Y. Two groups of entomopathogenic bacteria, Photorhabdus and Xenorhabdus, share an inhibitory action against phospholipase A2 to induce host immunodepression. J. Invertebr. Pathol. 2005, 89, 258–264. [Google Scholar] [CrossRef]
- Park, Y.; Kim, Y. Xenorhabdus nematophilus inhibits p-bromophenacyl bromide (BPB)-sensitive PLA2 of Spodoptera exigua. Arch. Insect. Biochem. Physiol. 2003, 54, 134–142. [Google Scholar] [CrossRef]
- Barletta, A.B.F.; Trisnadi, N.; Ramirez, J.L.; Barillas-Mury, C. Mosquito Midgut Prostaglandin Release Establishes Systemic Immune Priming. iScience 2019, 19, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Scarpati, M.; Qi, Y.; Govind, S.; Singh, S. A combined computational strategy of sequence and structural analysis predicts the existence of a functional eicosanoid pathway in Drosophila melanogaster. PLoS ONE 2019, 14, e0211897. [Google Scholar] [CrossRef]
- Panettieri, S.; Paddibhatla, I.; Chou, J.; Rajwani, R.; Moore, R.; Goncharuk, T.; John, G.; Govind, S. Discovery of aspirin-triggered eicosanoid-like mediators in a Drosophila metainflammation-blood tumor model. J. Cell Sci. 2019. [Google Scholar] [CrossRef] [Green Version]
- Haine, E.R.; Rolff, J.; Siva-Jothy, M.T. Functional consequences of blood clotting in insects. Dev. Comp. Immunol. 2007, 31, 456–464. [Google Scholar] [CrossRef]
- Krautz, R.; Arefin, B.; Theopold, U. Damage signals in the insect immune response. Front. Plant. Sci. 2014, 5, 342. [Google Scholar] [CrossRef] [Green Version]
- Bidla, G.; Hauling, T.; Dushay, M.S.; Theopold, U. Activation of Insect Phenoloxidase after Injury: Endogenous versus Foreign Elicitors. J. Innate Immun. 2009, 1, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Keehnen, N.L.P.; Rolff, J.; Theopold, U.; Wheat, C.W. Insect Antimicrobial Defences: A Brief History, Recent Findings, Biases, and a Way Forward in Evolutionary Studies. In Insect Immunity; Ligoxygakis, P., Ed.; Academic Press: Cambridge, MA, USA, 2017; Volume 52, pp. 1–33. [Google Scholar]
- Theopold, U.; Schmidt, O. Helix pomatia lectin and annexin V, two molecular probes for insect microparticles: Possible involvement in hemolymph coagulation. J. Insect Physiol. 1997, 43, 667–674. [Google Scholar] [CrossRef]
- Lacroix, R.; Dignat-George, F. Microparticles as a circulating source of procoagulant and fibrinolytic activities in the circulation. Thromb. Res. 2012, 129, S27–S29. [Google Scholar] [CrossRef] [PubMed]
- Loof, T.G.; Schmidt, O.; Herwald, H.; Theopold, U. Coagulation systems of invertebrates and vertebrates and their roles in innate immunity: The same side of two coins? J. Innate Immun. 2011, 3, 34–40. [Google Scholar] [CrossRef]
- Li, D.; Scherfer, C.; Korayem, A.M.; Zhao, Z.; Schmidt, O.; Theopold, U. Insect hemolymph clotting: Evidence for interaction between the coagulation system and the prophenoloxidase activating cascade. Insect. Biochem. Mol. Biol. 2002, 32, 919–928. [Google Scholar] [CrossRef]
- Fiusa, M.M.; Carvalho-Filho, M.A.; Annichino-Bizzacchi, J.M.; De Paula, E.V. Causes and consequences of coagulation activation in sepsis: An evolutionary medicine perspective. BMC Med. 2015, 13, 105. [Google Scholar] [CrossRef] [Green Version]
- Simonti, C.N.; Vernot, B.; Bastarache, L.; Bottinger, E.; Carrell, D.S.; Chisholm, R.L.; Crosslin, D.R.; Hebbring, S.J.; Jarvik, G.P.; Kullo, I.J.; et al. The phenotypic legacy of admixture between modern humans and Neandertals. Science 2016, 351, 737–741. [Google Scholar] [CrossRef] [Green Version]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dziedziech, A.; Shivankar, S.; Theopold, U. Drosophila melanogaster Responses against Entomopathogenic Nematodes: Focus on Hemolymph Clots. Insects 2020, 11, 62. https://doi.org/10.3390/insects11010062
Dziedziech A, Shivankar S, Theopold U. Drosophila melanogaster Responses against Entomopathogenic Nematodes: Focus on Hemolymph Clots. Insects. 2020; 11(1):62. https://doi.org/10.3390/insects11010062
Chicago/Turabian StyleDziedziech, Alexis, Sai Shivankar, and Ulrich Theopold. 2020. "Drosophila melanogaster Responses against Entomopathogenic Nematodes: Focus on Hemolymph Clots" Insects 11, no. 1: 62. https://doi.org/10.3390/insects11010062