
Investigations of Ascorbic Acid Synthesis and Distribution in Broiler Tissues at Different Post-Hatch Days
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Management, and Sampling
2.2. Measurement of L-gulonolactone Oxidase Activity in Kidneys
2.3. Ascorbic Acid Tissue Deposition
2.4. Quantitative Real-Time PCR
2.5. Statistical Analysis
3. Results
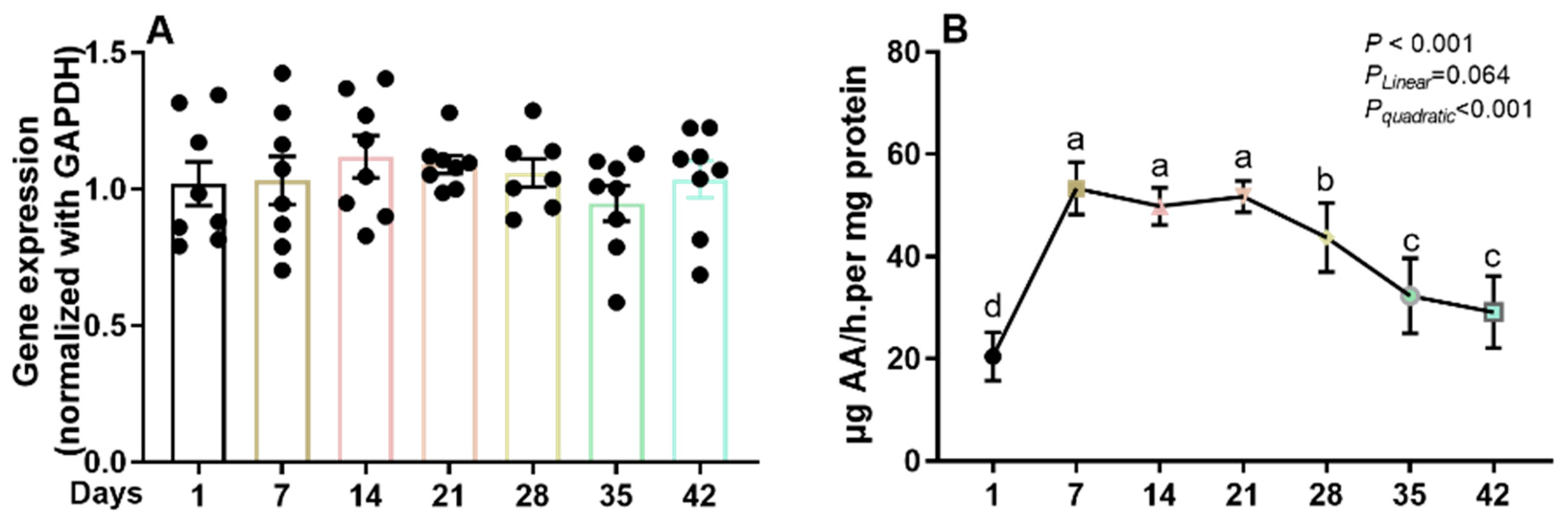
3.1. GLO Enzyme Activities
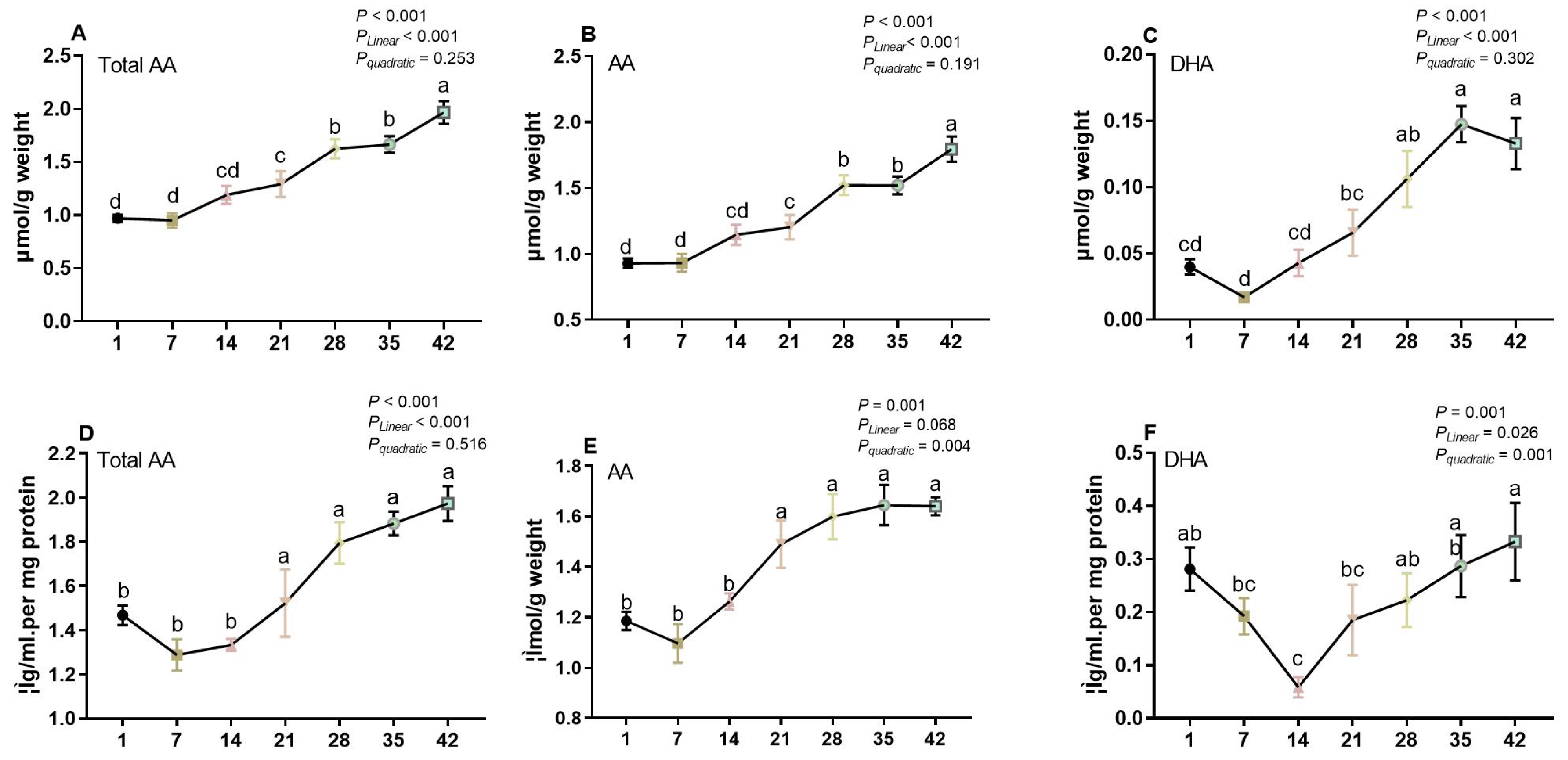
3.2. The Concentration of AA in Liver and Spleen of Broilers
3.3. The Gene Expression Levels of SVCTs in Ileum and Kidney of Broilers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Figueroa-Méndez, R.; Rivas-Arancibia, S. Vitamin C in Health and Disease: Its Role in the Metabolism of Cells and Redox State in the Brain. Front. Physiol. 2015, 6, 397. [Google Scholar] [CrossRef] [PubMed]
- Young, J.I.; Zuchner, S.; Wang, G. Regulation of the Epigenome by Vitamin C. Annu. Rev. Nutr. 2015, 35, 545–564. [Google Scholar] [CrossRef] [PubMed]
- Council, N.R. Nutrient Requirements of Poultry, 9th ed.; National Academies Press: Washington, DC, USA, 1994. [Google Scholar] [CrossRef]
- Hooper, C.L.; Maurice, D.V.; Lightsey, S.F.; Toler, J.E. Factors affecting ascorbic acid biosynthesis in chickens. I. Adaptation of an assay and the effect of age, sex, and food deprivation. J. Anim. Physiol. Anim. Nutr. 2000, 84, 48–56. [Google Scholar] [CrossRef]
- Gan, L.; Fan, H.; Nie, W.; Guo, Y. Ascorbic acid synthesis and transportation capacity in old laying hens and the effects of dietary supplementation with ascorbic acid. J. Anim. Sci. Biotechnol. 2018, 9, 71. [Google Scholar] [CrossRef] [PubMed]
- Johnston, L.; Laverty, G. Vitamin C transport and SVCT1 transporter expression in chick renal proximal tubule cells in culture. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 146, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Kuo, S.-M.; Tan, C.-H.; Dragan, M.; Wilson, J.X. Endotoxin Increases Ascorbate Recycling and Concentration in Mouse Liver. J. Nutr. 2005, 135, 2411–2416. [Google Scholar] [CrossRef] [PubMed]
- Linster, C.L.; Van Schaftingen, E. Vitamin C Biosynthesis, recycling and degradation in mammals. FEBS J. 2007, 274, 1–22. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, J.C. Detoxification effects of ascorbic acid on the oxidative stress, neurotoxicity, and metallothionein (MT) gene expression in juvenile rockfish, Sebastes schlegelii by the dietary chromium exposure. Fish Shellfish Immunol. 2023, 132, 108464. [Google Scholar] [CrossRef]
- Carr, A.C.; Maggini, S. Vitamin C and Immune Function. Nutrients 2017, 9, 1211. [Google Scholar] [CrossRef]
- Maurice, D.V.; Lightsey, S.F.; Abudabos, A.; Toler, J.E. Factors affecting ascorbic acid biosynthesis in chickens. III. Effectof dietary fluoride on L-gulonolactone oxidase activity and tissue ascorbic acid (AsA) concentration. J. Anim. Physiol. Anim. Nutr. 2002, 86, 383–388. [Google Scholar] [CrossRef]
- Maurice, D.V.; Lightsey, S.F. Sexual difference in ascorbic acid synthesis, tissue ascorbic acid and plasma total antioxidant capacity in mature chickens. Br. Poult. Sci. 2007, 48, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Saki, A.A.; Rahmati, M.M.H.; Zamani, P.; Zaboli, K.; Matin, H.R.H. Can Vitamin C Elevate Laying Hen Performance, Egg and Plasma Characteristics Under Normal Environmental Temperature? Ital. J. Anim. Sci. 2010, 9, e60. [Google Scholar] [CrossRef]
- Gan, L.; Fan, H.; Mahmood, T.; Guo, Y. Dietary supplementation with vitamin C ameliorates the adverse effects of Salmonella Enteritidis-challenge in broilers by shaping intestinal microbiota. Poult. Sci. 2020, 99, 3663–3674. [Google Scholar] [CrossRef] [PubMed]
- Mbajiorgu, C.A. Effect of dietary ascorbic acid supplementation on egg production, egg quality and hatchability of indignous Venda chicken hens. Asian J. Anim. Vet. Adv. 2011, 6, 823–830. [Google Scholar] [CrossRef]
- Cheng, T.K.; Coon, C.N.; Hamre, M.L. Effect of environmental stress on the ascorbic acid requirement of laying hens. Poult. Sci. 1990, 69, 774–780. [Google Scholar] [CrossRef]
- Torki, M.; Zangeneh, S.; Habibian, M. Performance, Egg Quality Traits, and Serum Metabolite Concentrations of Laying Hens Affected by Dietary Supplemental Chromium Picolinate and Vitamin C Under a Heat-Stress Condition. Biol. Trace Elem. Res. 2013, 157, 120–129. [Google Scholar] [CrossRef]
- Ching, S.; Mahan, D.C.; Moreau, R.é.; Dabrowski, K. Modification of analytical procedures for determining vitamin C enzyme (L-gulonolactone oxidase) activity in swine liver. J. Nutr. Biochem. 2003, 14, 139–146. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Mahan, D.C.; Ching, S.; Dabrowski, K. Developmental aspects and factors influencing the synthesis and status of ascorbic Acid in the pig. Annu. Rev. Nutr. 2004, 24, 79–103. [Google Scholar] [CrossRef]
- Abidin, Z.; Khatoon, A. Heat stress in poultry and the beneficial effects of ascorbic acid (vitamin C) supplementation during periods of heat stress. Worlds Poult. Sci. J. 2013, 69, 135–152. [Google Scholar] [CrossRef]
- Chatterjee, I.B.; Mckee, R.W. Biosynthesis of l-ascorbic acid in rat liver microsomes: Influences of age, sex, dietary changes, and whole-body X-irradiation. Arch. Biochem. Biophys. 1965, 109, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Yew, M.S. Biosynthesis of ascorbic acid in chick embryos. Experientia 1985, 41, 943–944. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhao, J.; Wang, C.; Zhang, F.; Huang, X.; Ren, Z.; Yang, X.; Liu, Y.; Yang, X. Exploring the effectiveness of in ovo feeding of vitamin C based on the embryonic vitamin C synthesis and absorption in broiler chickens. J. Anim. Sci. Biotechnol. 2021, 12, 86. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.M.; Neha, J.; Hagen, T.M. Age-related decline of sodium-dependent ascorbic acid transport in isolated rat hepatocytes. Arch. Biochem. Biophys. 2003, 410, 112–120. [Google Scholar]
- Corpe, C.P.; Tu, H.; Eck, P.; Wang, J.; Faulhaber-Walter, R.; Schnermann, J.; Margolis, S.; Padayatty, S.; Sun, H.; Wang, Y.; et al. Vitamin C transporter Slc23a1 links renal reabsorption, vitamin C tissue accumulation, and perinatal survival in mice. J. Clin. Investig. 2010, 120, 1069–1083. [Google Scholar] [CrossRef]
- Iwama, M.; Amano, A.; Shimokado, K.; Maruyama, N.; Ishigami, A. The content of ascorbic acid in plasma and in 14 tissues of lambs of different ages during normal feeding and feeding restricted from time to time during the suckling period. J. Nutr. Sci. Vitaminol. 2012, 58, 169–174. [Google Scholar] [CrossRef]
- Maldonado, M.; Inostroza, E.; Pena, E.; Moncada, N.; Mardones, L.; Medina, J.L.; Munoz, A.; Gatica, M.; Villagran, M.; Escobar, E.; et al. Sustained blockade of ascorbic acid transport associated with marked SVCT1 loss in rat hepatocytes containing increased ascorbic acid levels after partial hepatectomy. Free Radic. Biol. Med. 2017, 108, 655–667. [Google Scholar] [CrossRef]
- Mohammed, B.M.; Fisher, B.J.; Huynh, Q.K.; Wijesinghe, D.S.; Chalfant, C.E.; Brophy, D.F.; Fowler, A.A.; Natarajan, R. Resolution of sterile inflammation: Role for vitamin C. Mediat. Inflamm. 2014, 2014, 173403. [Google Scholar] [CrossRef]
- Manning, J.; Mitchell, B.; Appadurai, D.A.; Shakya, A.; Pierce, L.J.; Wang, H.; Nganga, V.; Swanson, P.C.; May, J.M.; Tantin, D.; et al. Vitamin C promotes maturation of T-cells. Antioxid. Redox Signal. 2013, 19, 2054–2067. [Google Scholar] [CrossRef]
- May, J.M.; Li, L.; Qu, Z.C.; Huang, J. Ascorbate uptake and antioxidant function in peritoneal macrophages. Arch. Biochem. Biophys. 2005, 440, 165–172. [Google Scholar] [CrossRef]
- Mebius, R.E.; Kraal, G. Structure and function of the spleen. Nat. Rev. Immunol. 2005, 5, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Elliott, K.E.C.; Durojaye, O.A.; Fatemi, S.A.; Schilling, M.W.; Peebles, E.D. Effects of in ovo injection of L-ascorbic acid on growth performance, carcass composition, plasma antioxidant capacity, and meat quality in broiler chickens. Poult. Sci. 2019, 98, 3617–3625. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Item, % | 0~3 Weeks | 4~6 Weeks |
|---|---|---|
| Corn | 53.82 | 59.76 |
| Soybean meal | 38.54 | 33.22 |
| Soybean oil | 3.52 | 3.5 |
| Calcium carbonate | 1.76 | 1.21 |
| Calcium phosphate | 1.26 | 1.39 |
| Sodium chloride | 0.35 | 0.3 |
| Choline chloride (50%) | 0.2 | 0.2 |
| L-Lys × HCl (78%) | 0.2 | 0.16 |
| DL-Met (98%) | 0.15 | 0.1 |
| Mineral premix 1 | 0.18 | 0.13 |
| Vitamin premix 2 | 0.02 | 0.02 |
| Total | 100 | 100 |
| Nutrient Level 3 | ||
| AME, MJ/kg | 12.36 | 12.6 |
| Crude protein, % | 22.02 | 19.72 |
| Calcium, % | 1 | 0.9 |
| Available phosphorus, % | 0.45 | 0.35 |
| Lysine, % | 1.19 | 1.04 |
| Methionine, % | 0.5 | 0.43 |
| Methionine + Cystine, % | 0.85 | 0.75 |
| Gene Name | Primer Sequence 1, 5′-3′ | NCBI Number | Product Size | |
|---|---|---|---|---|
| GAPDH | F 2 | GACCCCAGCAACATCAAATG | NM_204305.1 | 110 bp |
| R | TTAGCACCACCCTTCAGATG | |||
| GLO | F | TCTCCTCTGGATCAGCACCT | XM_015285218.1 | 131 bp |
| R | AGCGGCACTCGTAGTTGAAG | |||
| SVCT1 | F | GGGATACCCACGGTGACCTC | XM_004944768.2 | 100 bp |
| R | GCCGTGCACAGGAGTAGTAA | |||
| SVCT2 | F | TGTCTTGTGCTCCTCCTCCT | NM_001145227.1 | 101 bp |
| R | TCCATTCCCTGTCCCAAATA | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gan, L.; Zhao, Y.; Wang, P.; Zhao, C.; Li, Y.; Huang, W.; Shi, L.; Cui, Y.; Qiao, H.; Wang, J.; et al. Investigations of Ascorbic Acid Synthesis and Distribution in Broiler Tissues at Different Post-Hatch Days. Life 2023, 13, 1137. https://doi.org/10.3390/life13051137
Gan L, Zhao Y, Wang P, Zhao C, Li Y, Huang W, Shi L, Cui Y, Qiao H, Wang J, et al. Investigations of Ascorbic Acid Synthesis and Distribution in Broiler Tissues at Different Post-Hatch Days. Life. 2023; 13(5):1137. https://doi.org/10.3390/life13051137
Chicago/Turabian StyleGan, Liping, Yifeng Zhao, Peng Wang, Chenkai Zhao, Yilei Li, Weihao Huang, Liuying Shi, Yaoming Cui, Hanzhen Qiao, Jinrong Wang, and et al. 2023. "Investigations of Ascorbic Acid Synthesis and Distribution in Broiler Tissues at Different Post-Hatch Days" Life 13, no. 5: 1137. https://doi.org/10.3390/life13051137