
Effects of Vitamin A on Immune Responses and Vitamin A Metabolism in Broiler Chickens Challenged with Necrotic Enteritis
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Bird Management
2.2. Co-infection of Eimeria spp. and C. perfringens
2.3. Sample Collection
2.4. Jejunal NE Lesion Score
2.5. Determination of Serum Biochemical Indices
2.6. Serum Immunoglobulin (Ig) A and IgG Assay
2.7. Immunohistochemical Analysis
2.8. Measurement of VitA Content in Liver
2.9. Serum PGE2 Assay
2.10. RNA Isolation and Quantitative Real-Time PCR
2.11. Statistical Analysis
3. Results
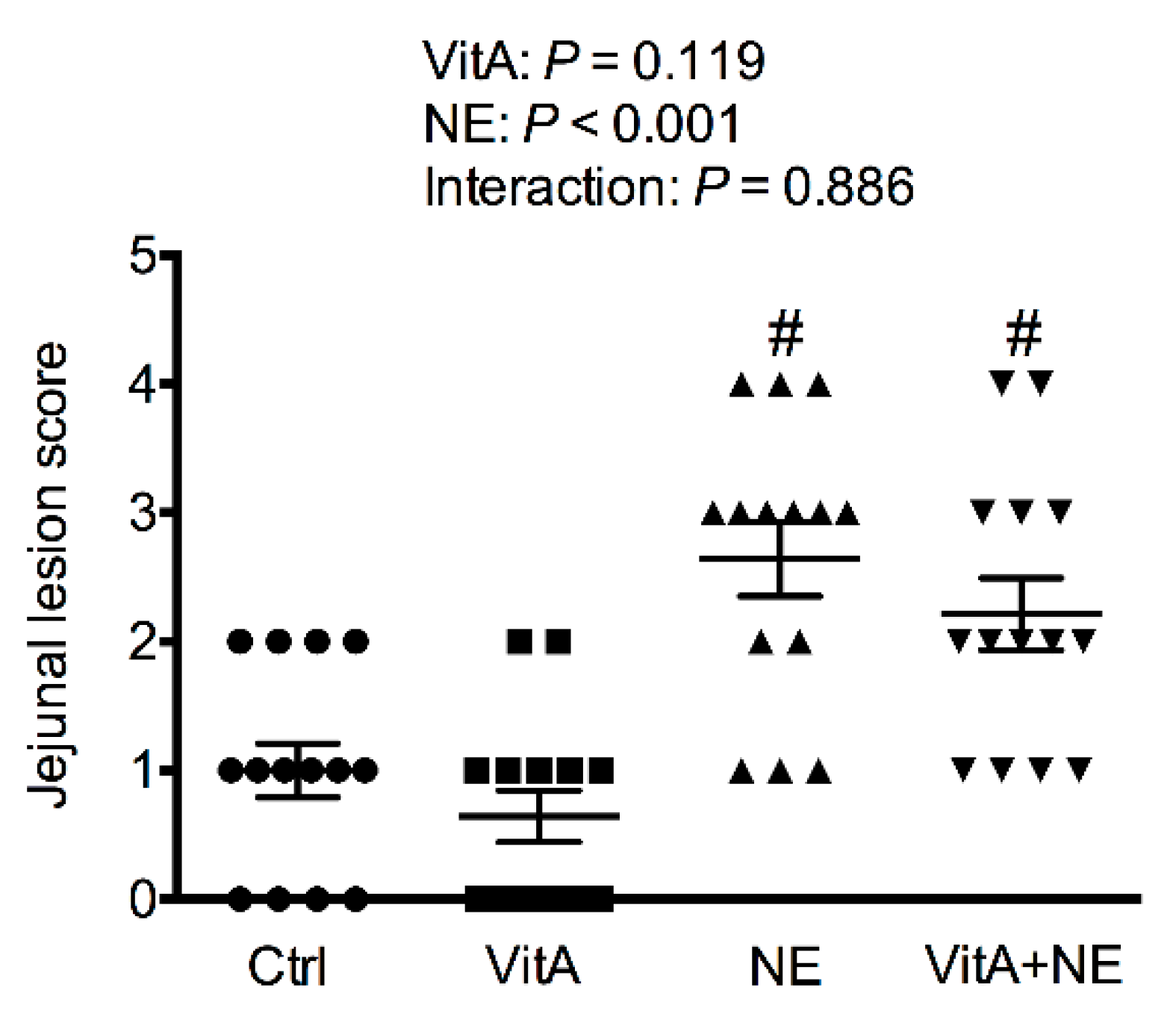
3.1. Jejunal NE Lesion Score
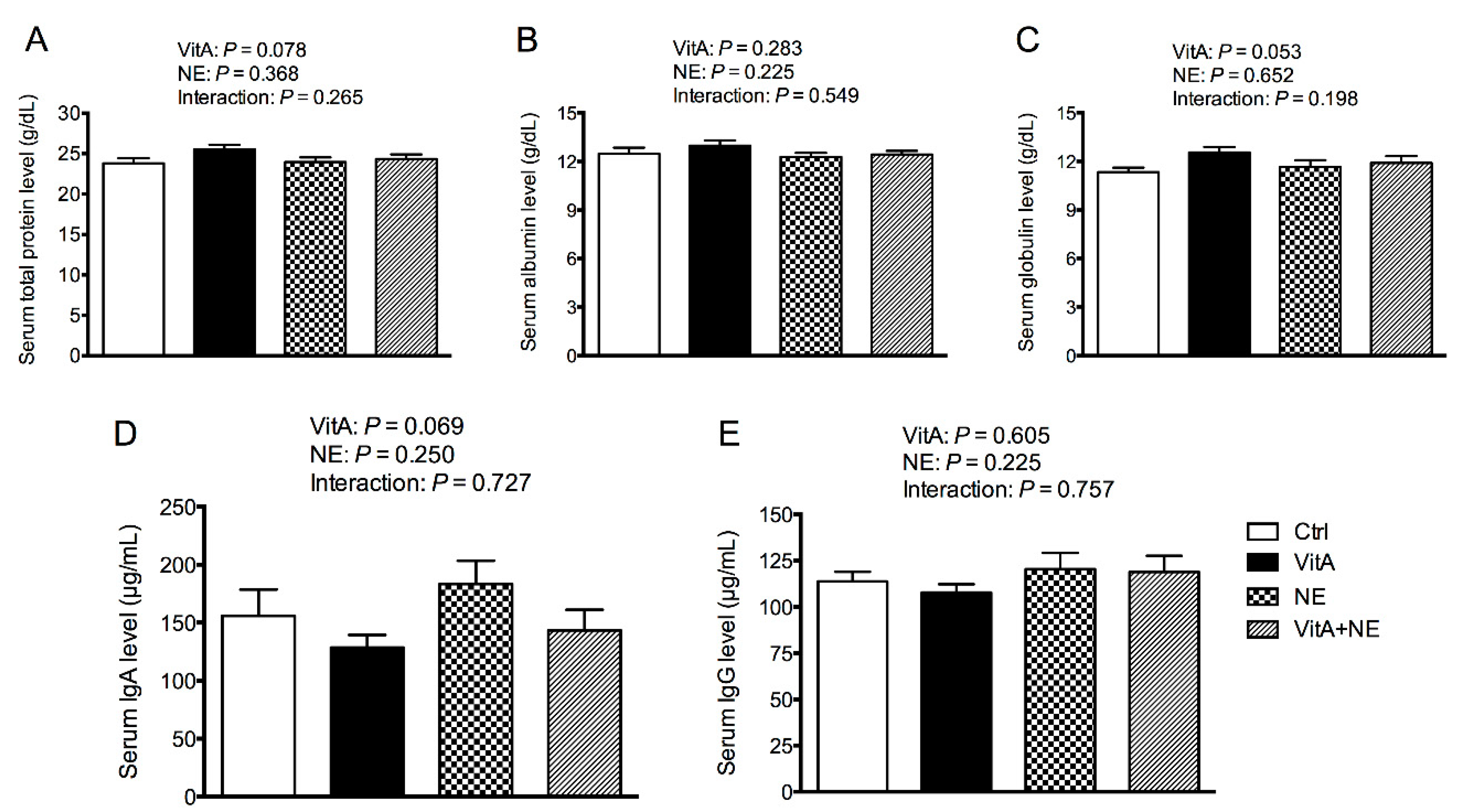
3.2. Serum Biochemical Indices
3.3. Serum Immune Parameters
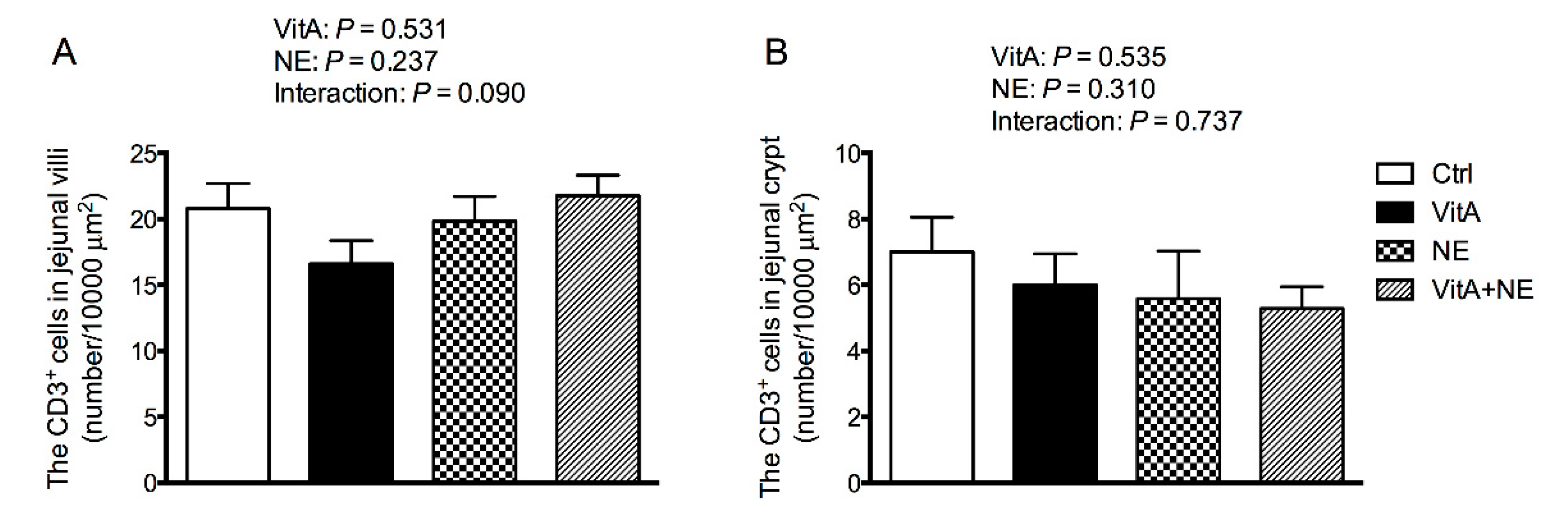
3.4. Jejunal CD3+ T Cell Density
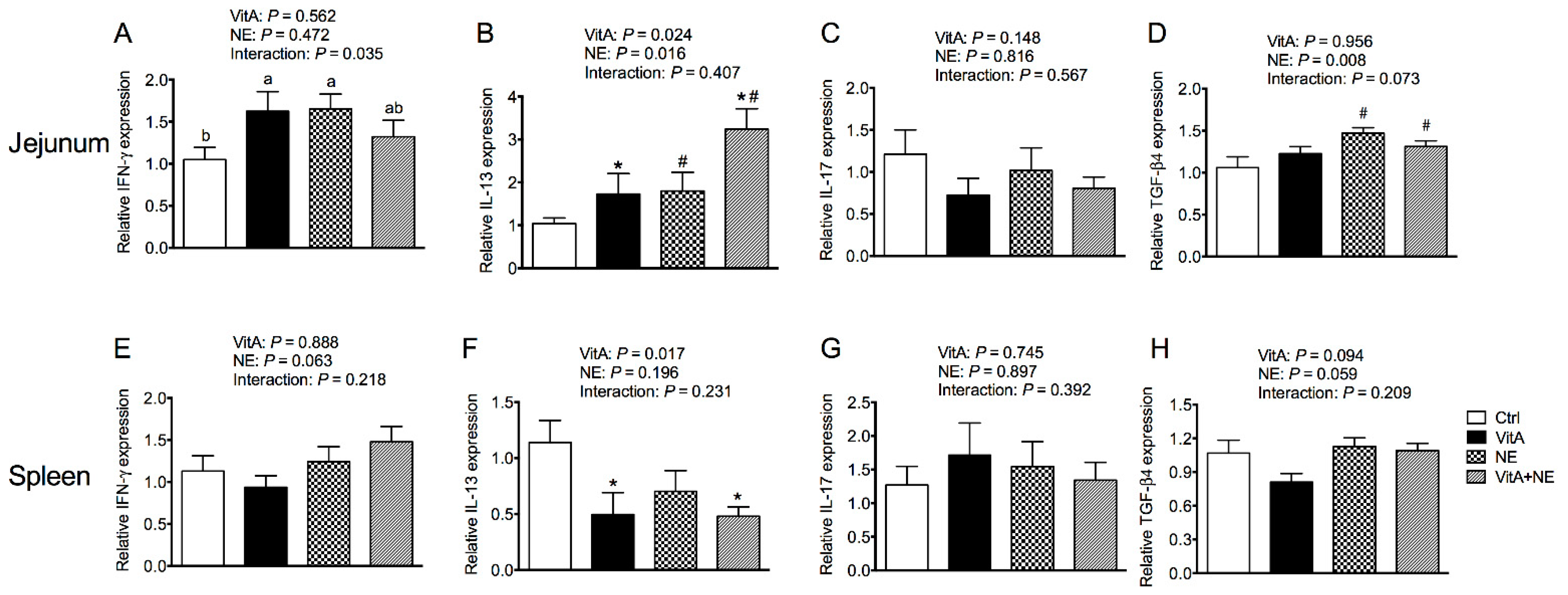
3.5. Immune Gene Expression in the Jejunum and Spleen
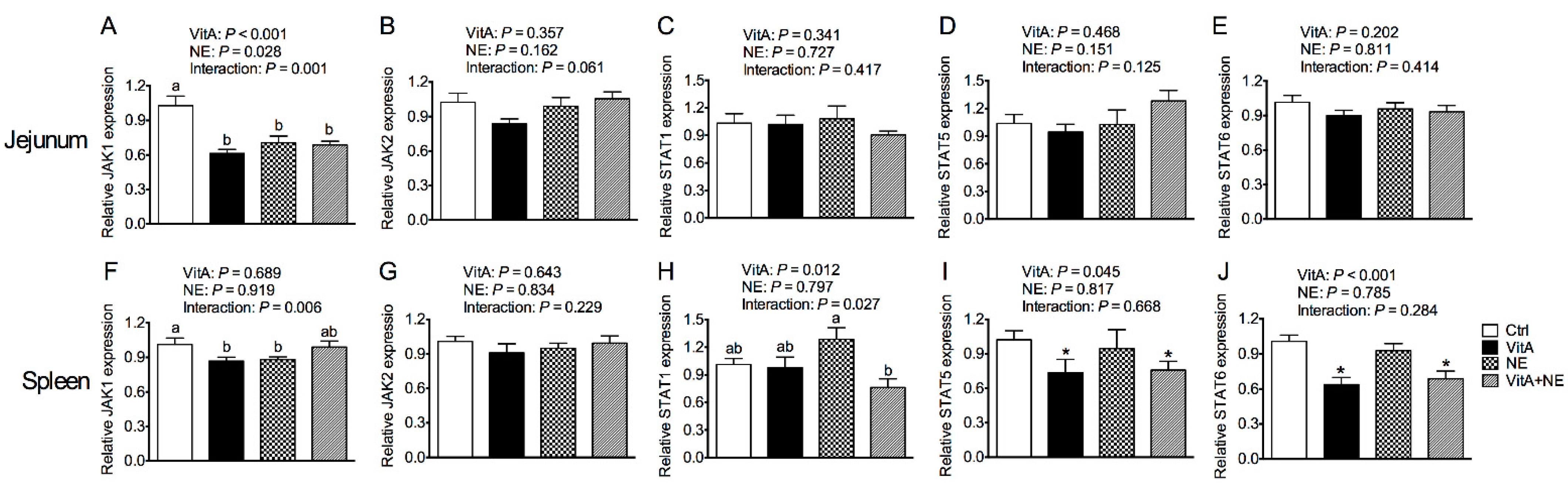
3.6. Gene Expression of the JAK-STAT Signaling Pathway Components in the Jejunum and Spleen
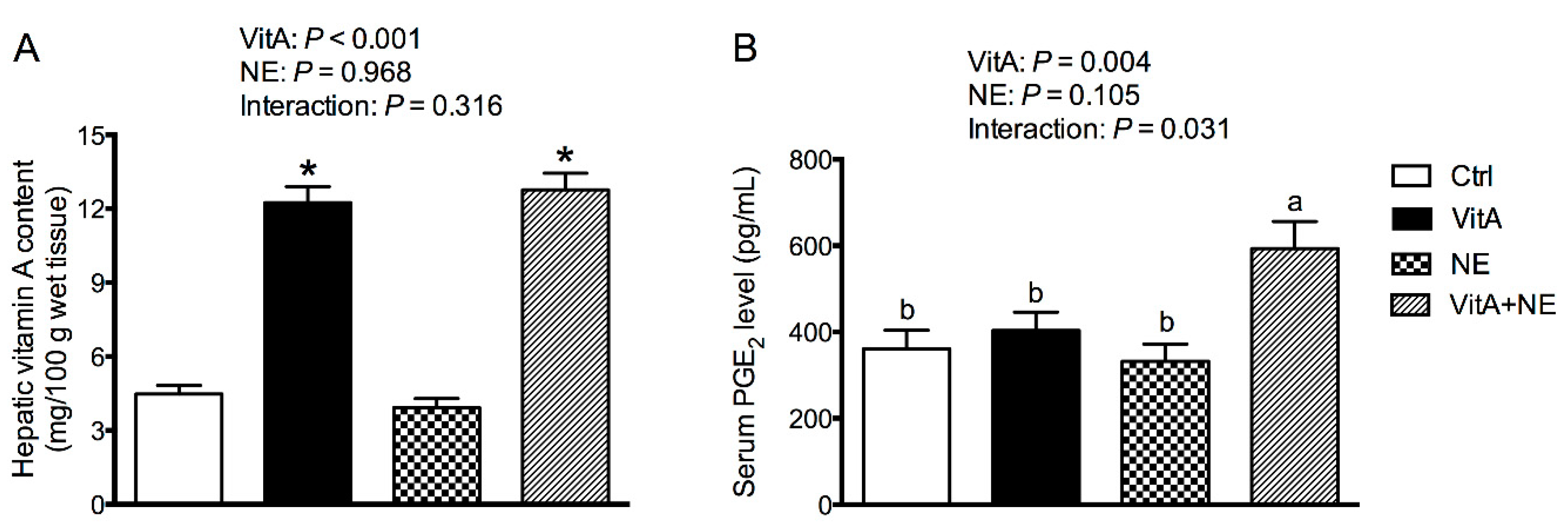
3.7. Hepatic VitA Content and Serum PGE2 Level
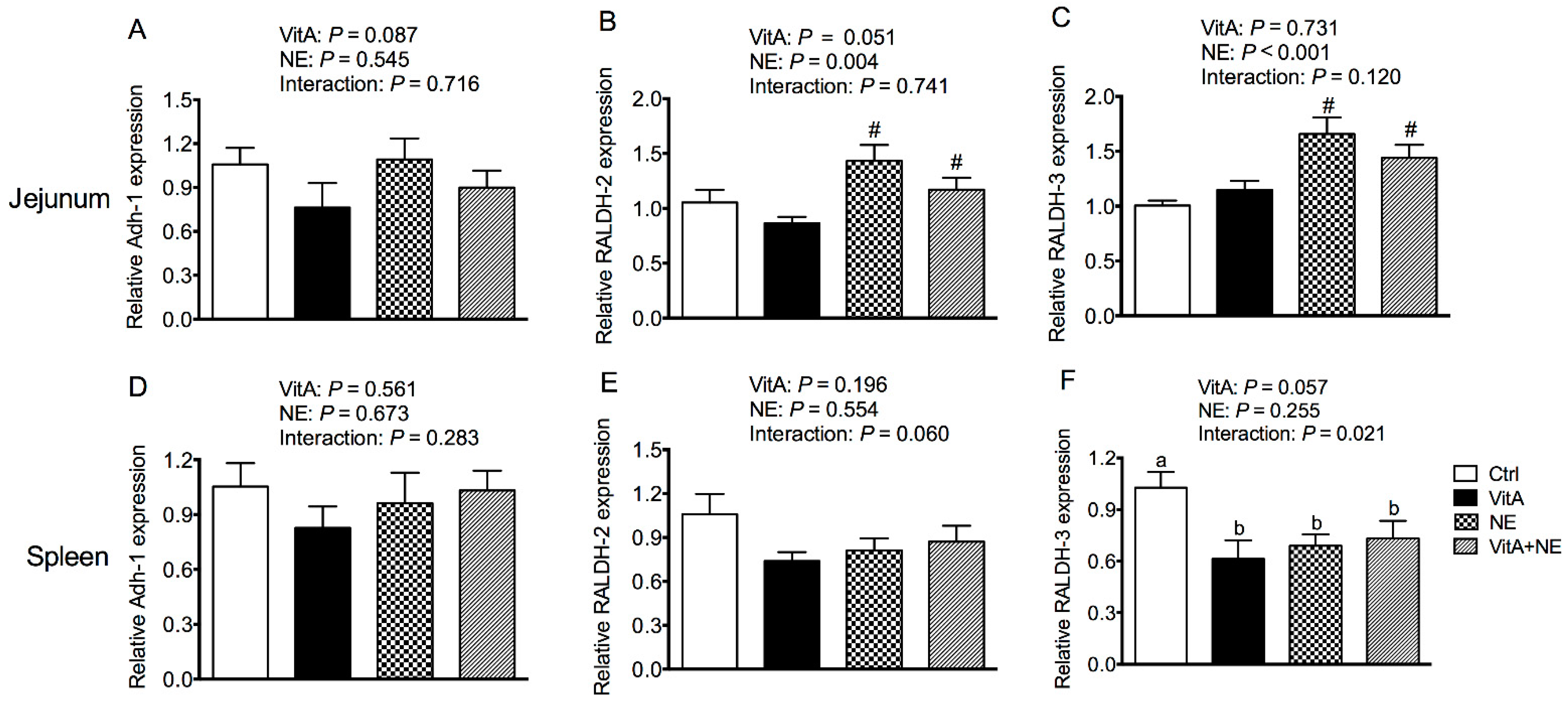
3.8. The Relative mRNA Expression of Genes Involved in Retinoic Acid Synthesis in the Jejunum and Spleen
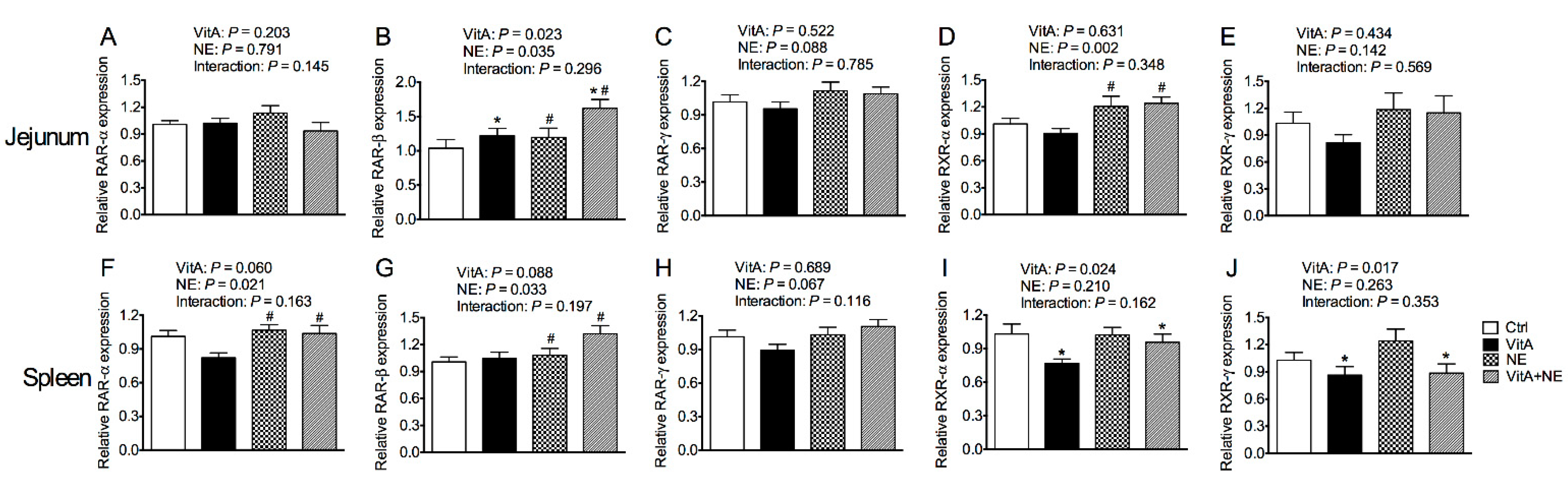
3.9. The Relative mRNA expression of Genes Encoding Retinoic Acid Receptors in the Jejunum and Spleen
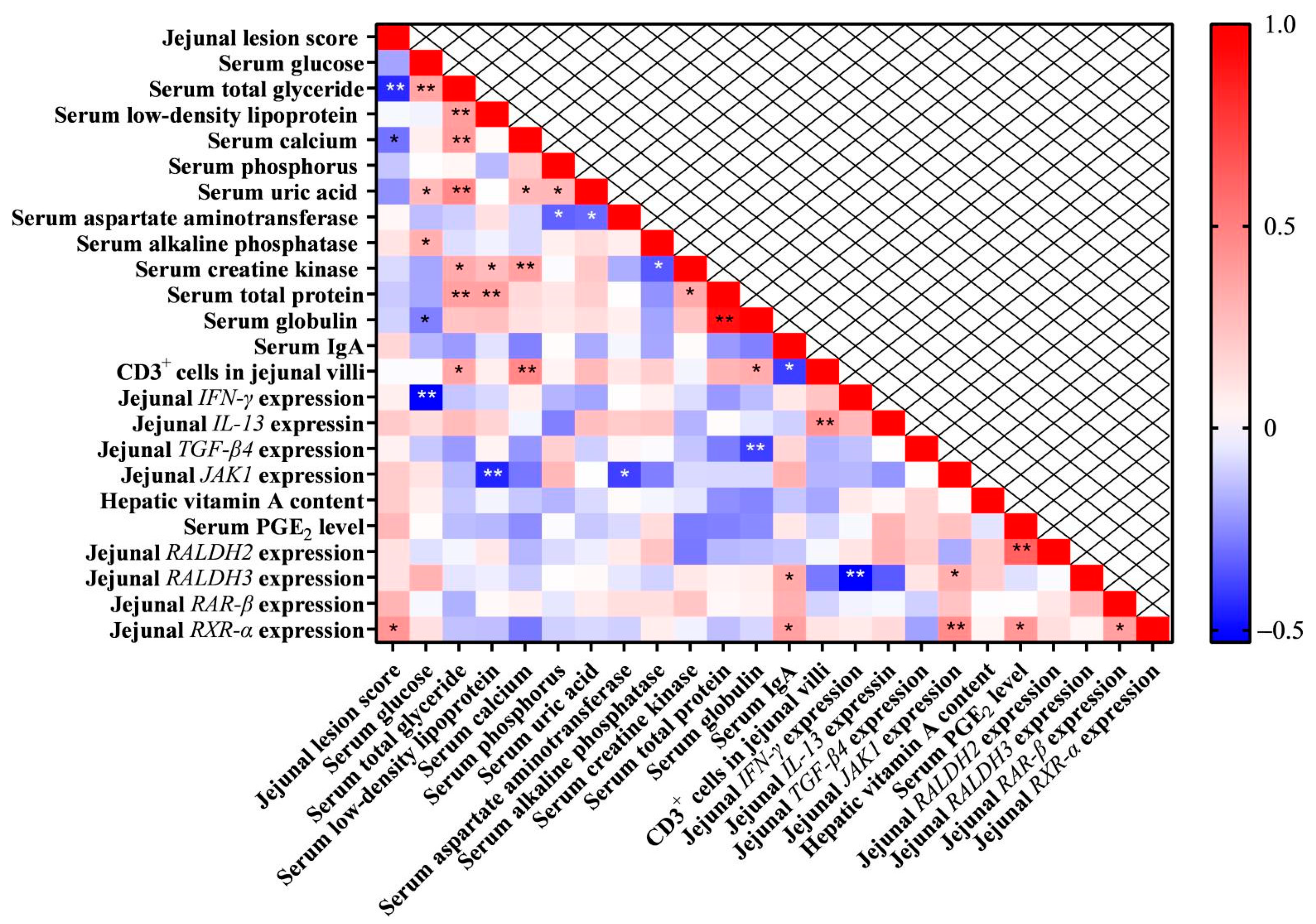
3.10. Pearson Correlation Analysis of Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, Z.; Liu, Y.; Qi, G.; Brand, D.; Zheng, S.G. Role of vitamin A in the immune system. J. Clin. Med. 2018, 7, 258. [Google Scholar] [CrossRef] [PubMed]
- Munteanu, C.; Schwartz, B. The relationship between nutrition and the immune system. Front. Nutr. 2022, 9, 1082500. [Google Scholar] [CrossRef] [PubMed]
- Lessard, M.; Hutchings, D.; Cave, N.A. Cell-mediated and humoral immune responses in broiler chickens maintained on diets containing different levels of vitamin A. Poult. Sci. 1997, 76, 1368–1378. [Google Scholar] [CrossRef] [PubMed]
- Pino-Lagos, K.; Guo, Y.; Noelle, R.J. Retinoic acid: A key player in immunity. BioFactors 2010, 36, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Rochette-Egly, C.; Germain, P. Dynamic and combinatorial control of gene expression by nuclear retinoic acid receptors (RARs). Nucl. Recept. Signal. 2009, 7, e005. [Google Scholar] [CrossRef]
- Collins, C.B.; Aherne, C.M.; Kominsky, D.; McNamee, E.N.; Lebsack, M.D.P.; Eltzschig, H.; Jedlicka, P.; Rivera-Nieves, J. Retinoic acid attenuates ileitis by restoring the balance between T-helper 17 and T regulatory cells. Gastroenterology 2011, 141, 1821–1831. [Google Scholar] [CrossRef]
- Manicassamy, S.; Ravindran, R.; Deng, J.; Oluoch, H.; Denning, T.L.; Kasturi, S.P.; Rosenthal, K.M.; Evavold, B.D.; Pulendran, B. Toll-like receptor 2-dependent induction of vitamin A-metabolizing enzymes in dendritic cells promotes T regulatory responses and inhibits autoimmunity. Nat. Med. 2009, 15, 401–409. [Google Scholar] [CrossRef]
- Uniyal, S.; Dhasmana, A.; Tyagi, A.; Muyal, J.P. ATRA reduces inflammation and improves alveolar epithelium regeneration in emphysematous rat lung. Biomed. Pharmacother. 2018, 108, 1435–1450. [Google Scholar] [CrossRef]
- Choi, W.-H.; Ji, K.-A.; Jeon, S.-B.; Yang, M.-S.; Kim, H.; Min, K.-J.; Shong, M.; Jou, I.; Joe, E.-H. Anti-inflammatory roles of retinoic acid in rat brain astrocytes: Suppression of interferon-γ-induced JAK/STAT phosphorylation. Biochem. Biophys. Res. Commun. 2005, 329, 125–131. [Google Scholar] [CrossRef]
- Shojadoost, B.; Vince, A.R.; Prescott, J.F. The successful experimental induction of necrotic enteritis in chickens by Clostridium perfringens: A critical review. Vet. Res. 2012, 43, 74. [Google Scholar] [CrossRef]
- Fasina, Y.O.; Lillehoj, H.S. Characterization of intestinal immune response to Clostridium perfringens infection in broiler chickens. Poult. Sci. 2019, 98, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Li, P.; Yan, S.; Liu, D.; Gao, M.; Lv, H.; Lv, Z.; Guo, Y. Effects of dietary astragalus polysaccharide supplementation on the Th17/Treg balance and the gut microbiota of broiler chickens challenged with necrotic enteritis. Front. Immunol. 2022, 13, 781934. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Gan, L.; Shahid, M.S.; Lv, Z.; Fan, H.; Liu, D.; Guo, Y. In vivo and in vitro protective effect of arginine against intestinal inflammatory response induced by Clostridium perfringens in broiler chickens. J. Anim. Sci. Biotechnol. 2019, 10, 73. [Google Scholar] [CrossRef] [PubMed]
- Tejón, G.; Manríquez, V.; De Calisto, J.; Flores-Santibáñez, F.; Hidalgo, Y.; Crisóstomo, N.; Fernández, D.; Sauma, D.; Mora, J.R.; Bono, M.R.; et al. Vitamin A impairs the reprogramming of Tregs into IL-17-producing cells during intestinal inflammation. BioMed Res. Int. 2015, 2015, 137893. [Google Scholar] [CrossRef]
- Penny, H.L.; Prestwood, T.R.; Bhattacharya, N.; Sun, F.; Kenkel, J.A.; Davidson, M.G.; Shen, L.; Zuniga, L.A.; Seeley, E.S.; Pai, R.; et al. Restoring retinoic acid attenuates intestinal inflammation and tumorigenesis in APCMin/+ mice. Cancer Immunol. Res. 2016, 4, 917–926. [Google Scholar] [CrossRef]
- Moore, R.J. Necrotic enteritis predisposing factors in broiler chickens. Avian Pathol. 2016, 45, 275–281. [Google Scholar] [CrossRef]
- Dalloul, R.A.; Lillehoj, H.S.; Shellem, T.A.; Doerr, J.A. Effect of vitamin A deficiency on host intestinal immune response to Eimeria acervulina in broiler chickens. Poult. Sci. 2002, 81, 1509–1515. [Google Scholar] [CrossRef]
- Dalloul, R.A.; Lillehoj, H.S.; Shellem, T.A.; Doerr, J.A. Intestinal immunomodulation by vitamin A deficiency and lactobacillus-based probiotic in Eimeria acervulina—Infected broiler chickens. Avian Dis. 2003, 47, 1313–1320. [Google Scholar] [CrossRef]
- Preclik, G.; Stange, E.F.; Ditschuneit, H. Activation of PGE2-secretion from gastric mucosa by a type I phospholipase C is mediated by a direct release of arachidonic acid. Clin. Physiol. Biochem. 1992, 9, 78–86. [Google Scholar]
- Stock, A.; Booth, S.; Cerundolo, V. Prostaglandin E2 suppresses the differentiation of retinoic acid-producing dendritic cells in mice and humans. J. Exp. Med. 2011, 208, 761–773. [Google Scholar] [CrossRef]
- Wu, Y.; Shao, Y.; Song, B.; Zhen, W.; Wang, Z.; Guo, Y.; Shahid, M.S.; Nie, W. Effects of Bacillus coagulans supplementation on the growth performance and gut health of broiler chickens with Clostridium perfringens-induced necrotic enteritis. J. Anim. Sci. Biotechnol. 2018, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhen, W.; Geng, Y.; Wang, Z.; Guo, Y. Pretreatment with probiotic Enterococcus faecium NCIMB 11181 ameliorates necrotic enteritis-induced intestinal barrier injury in broiler chickens. Sci. Rep. 2019, 9, 10256. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Xi, Y.; Xia, Y.; Wu, T.; Zhao, D.; Zhang, Z.; Ding, B. Dietary Lactobacillus fermentum and Bacillus coagulans supplementation modulates intestinal immunity and microbiota of broiler chickens challenged by Clostridium perfringens. Front. Vet. Sci. 2021, 8, 680742. [Google Scholar] [CrossRef] [PubMed]
- Idi, A.; Permin, A.; Jensen, S.K.; Murrell, K.D. Effect of a minor vitamin A deficiency on the course of infection with Ascaridia galli (Schrank, 1788) and the resistance of chickens. Helminthologia 2007, 44, 3–9. [Google Scholar] [CrossRef]
- Heid, C.A.; Stevens, J.; Livak, K.J.; Williams, P.M. Real time quantitative PCR. Genome Res. 1996, 6, 986–994. [Google Scholar] [CrossRef]
- Kulkarni, R.R.; Gaghan, C.; Correll, K.; Sharif, S.; Taha-Abdelaziz, K. Probiotic as alternatives to antibiotics for the prevention and control of necrotic enteritis in chickens. Pathogens 2022, 11, 692. [Google Scholar] [CrossRef]
- Li, P.; Liu, C.; Niu, J.; Zhang, Y.; Li, C.; Zhang, Z.; Guo, S.; Ding, B. Effects of dietary supplementation with vitamin A on antioxidant and intestinal barrier function of broilers co-infected with coccidia and Clostridium perfringens. Animals 2022, 12, 3431. [Google Scholar] [CrossRef]
- Xue, G.D.; Barekatain, R.; Wu, S.B.; Chot, M.; Swick, R.A. Dietary L-glutamine supplementation improves growth performance, gut morphology, and serum biochemical indices of broiler chickens during necrotic enteritis challenge. Poult. Sci. 2018, 97, 1334–1341. [Google Scholar] [CrossRef]
- Savaris, V.D.L.; Souza, C.; Wachholz, L.; Broch, J.; Polese, C.; Carvalho, P.L.O.; Pozza, P.C.; Eyng, C.; Nunes, R.V. Interactions between lipid source and vitamin A on broiler performance, blood parameters, fat and protein deposition rate, and bone development. Poult. Sci. 2021, 100, 174–185. [Google Scholar] [CrossRef]
- Rombout, J.H.W.M.; Sijtsma, S.R.; West, C.E.; Karabinis, Y.; Sijtsma, O.K.W.; Van Der Zijpp, A.J.; Koch, G. Effect of vitamin A deficiency and Newcastle disease virus infection on IgA and IgM secretion in chickens. Br. J. Nutr. 1992, 68, 753–763. [Google Scholar] [CrossRef]
- Alizadeh, M.; Astill, J.; Alqazlan, N.; Shojadoost, B.; Taha-Abdelaziz, K.; Bavananthasivam, J.; Doost, J.S.; Sedeghiisfahani, N.; Sharif, S. In ovo co-administration of vitamins (A and D) and probiotic lactobacilli modulates immune responses in broiler chickens. Poult. Sci. 2022, 101, 101717. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.T.; Lillehoj, H.S. The role of host genetic factors and host immunity in necrotic enteritis. Avian Pathol. 2016, 45, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Sersun Calefi, A.; Quinteiro-Filho, W.M.; de Siqueira, A.; Nacimento Lima, A.P.; Cruz, D.S.G.; Queiroz Hazarbassanov, N.; Auciello Salvagni, F.; Borsoi, A.; de Oliveira Massoco Salles Gomes, C.; Maiorka, P.C.; et al. Heat stress, Eimeria spp. and C. perfringens infections alone or in combination modify gut Th1/Th2 cytokine balance and avian necrotic enteritis pathogenesis. Vet. Immunol. Immunopathol. 2019, 210, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Shojadoost, B.; Alizadeh, M.; Taha-Abdelaziz, K.; Doost, J.S.; Astill, J.; Sharif, S. In ovo inoculation of vitamin A modulates chicken embryo immune functions. J. Interferon Cytokine Res. 2021, 41, 20–28. [Google Scholar] [CrossRef]
- Fan, X.; Liu, S.; Liu, G.; Zhao, J.; Jiao, H.; Wang, X.; Song, Z.; Lin, H. Vitamin A deficiency impairs mucin expression and suppresses the mucosal immune function of the respiratory tract in chicks. PLoS ONE 2015, 10, e0139131. [Google Scholar] [CrossRef]
- Banerjee, S.; Biehl, A.; Gadina, M.; Hasni, S.; Schwartz, D.M. JAK-STAT signaling as a target for inflammatory and autoimmune diseases: Current and future prospects. Drugs 2017, 77, 521–546. [Google Scholar] [CrossRef]
- Ou, A.; Ott, M.; Fang, D.; Heimberger, A.B. The role and therapeutic targeting of JAK/STAT signaling in glioblastoma. Cancers 2021, 13, 437. [Google Scholar] [CrossRef]
- Truong, A.D.; Rengaraj, D.; Hong, Y.; Hoang, C.T.; Hong, Y.H.; Lillehoj, H.S. Differentially expressed JAK-STAT signaling pathway genes and target microRNAs in the spleen of necrotic enteritis-afflicted chicken lines. Res. Vet. Sci. 2017, 115, 235–243. [Google Scholar] [CrossRef]
- Truong, A.D.; Rengaraj, D.; Hong, Y.; Hoang, C.T.; Hong, Y.H.; Lillehoj, H.S. Analysis of JAK-STAT signaling pathway genes and their microRNAs in the intestinal mucosa of genetically disparate chicken lines induced with necrotic enteritis. Vet. Immunol. Immunopathol. 2017, 187, 1–9. [Google Scholar] [CrossRef]
- Blomhoff, R.; Wake, K. Perisinusoidal stellate cells of the liver: Important roles in retinol metabolism and fibrosis. FASEB J. 1991, 5, 271–277. [Google Scholar] [CrossRef]
- Yuan, J.; Roshdy, A.R.; Guo, Y.; Wang, Y.; Guo, S. Effect of dietary vitamin A on reproductive performance and immune response of broiler breeders. PLoS ONE 2014, 9, e105677. [Google Scholar] [CrossRef] [PubMed]
- Szymański, Ł.; Skopek, R.; Palusińska, M.; Schenk, T.; Stengel, S.; Lewicki, S.; Kraj, L.; Kamiński, P.; Zelent, A. Retinoic acid and its derivatives in skin. Cells 2020, 9, 2660. [Google Scholar] [CrossRef] [PubMed]
- Cattin, A.; Wacleche, V.S.; Fonseca Do Rosario, N.; Marchand, L.R.; Dias, J.; Gosselin, A.; Cohen, E.A.; Estaquier, J.; Chomont, N.; Routy, J.-P.; et al. RALDH activity induced by bacterial/fungal pathogens in CD16+ monocyte-derived dendritic cells boosts HIV infection and outgrowth in CD4+ T Cells. J. Immunol. 2021, 206, 2638–2651. [Google Scholar] [CrossRef] [PubMed]
- Broadhurst, M.J.; Leung, J.M.; Lim, K.C.; Girgis, N.M.; Gundra, U.M.; Fallon, P.G.; Premenko-Lanier, M.; McKerrow, J.H.; McCune, J.M.; Loke, P. Upregulation of retinal dehydrogenase 2 in alternatively activated macrophages during retinoid-dependent type-2 immunity to helminth infection in mice. PLoS Pathog. 2012, 8, e1002883. [Google Scholar] [CrossRef]
- Owusu, S.A.; Ross, A.C. Retinoid homeostatic gene expression in liver, lung and kidney: Ontogeny and response to vitamin A-retinoic acid (VARA) supplementation from birth to adult age. PLoS ONE 2016, 11, e0145924. [Google Scholar] [CrossRef]
- Barrea, L.; Di Somma, C.; Muscogiuri, G.; Tarantino, G.; Tenore, G.C.; Orio, F.; Colao, A.; Savastano, S. Nutrition, inflammation and liver-spleen axis. Crit. Rev. Food Sci. Nutr. 2018, 58, 3141–3158. [Google Scholar] [CrossRef]
- Viera-Vera, J.; García-Arrarás, J.E. Retinoic acid signaling is associated with cell proliferation, muscle cell dedifferentiation, and overall rudiment size during intestinal regeneration in the sea cucumber, Holothuria glaberrima. Biomolecules 2019, 9, 873. [Google Scholar] [CrossRef]
- Krüger, K.A.; Blum, J.W.; Greger, D.L. Expression of nuclear receptor and target genes in liver and intestine of neonatal calves fed colostrum and vitamin A. J. Dairy Sci. 2005, 88, 3971–3981. [Google Scholar] [CrossRef]
- Dong, P.; Tao, Y.; Yang, Y.; Wang, W. Expression of retinoic acid receptors in intestinal mucosa and the effect of vitamin A on mucosal immunity. Nutrition 2010, 26, 740–745. [Google Scholar] [CrossRef]
- Luo, M.; Yeruva, S.; Liu, Y.; Chodisetti, G.; Riederer, B.; Menon, M.B.; Tachibana, K.; Doi, T.; Seidler, U.E. IL-1β-induced downregulation of the multifunctional PDZ adaptor PDZK1 is attenuated by ERK inhibition, RXRα, or PPARα stimulation in enterocytes. Front. Physiol. 2017, 8, 00061. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item (%, Unless Otherwise Indicated) | Starter Diet (d 1–21) | Grower Diet (d 22–28) |
|---|---|---|
| Ingredients | ||
| Corn (crude protein, 7.8%) | 51.73 | 57.68 |
| Soybean meal (crude protein, 43.0%) | 40.73 | 35.15 |
| Soybean oil | 3.36 | 3.66 |
| Dicalcium phosphate | 1.92 | 1.33 |
| Limestone | 1.16 | 1.26 |
| Sodium chloride | 0.35 | 0.35 |
| DL-Methionine (98%) | 0.26 | 0.13 |
| Choline chloride (50%) | 0.25 | 0.20 |
| Vitamin premix 1 | 0.04 | 0.04 |
| Trance mineral premix 2 | 0.20 | 0.20 |
| Calculated nutrient levels | ||
| Metabolic energy (Mcal/kg) | 2.92 | 3.00 |
| Crude protein | 21.50 | 19.50 |
| Calcium | 1.00 | 0.90 |
| Available Phosphorus | 0.45 | 0.35 |
| Lysine | 1.17 | 1.04 |
| Methionine | 0.57 | 0.40 |
| Threonine | 0.82 | 0.74 |
| Gene Name | Accession Number | Forward Sequence (5′-3′) | Reverse Sequence (5′-3′) | Annealing and Extension Temperature (°C) |
|---|---|---|---|---|
| β-actin | NM_205518 | GAGAAATTGTGCGTGACATCA | CCTGAACCTCTCATTGCCA | 60 |
| IFN-γ | Y07922 | AGCTGACGGTGGACCTATTATT | GGCTTTGCGCTGGATTC | 60 |
| IL-13 | AJ621735 | CCAGGGCATCCAGAAGC | CAGTGCCGGCAAGAAGTT | 62 |
| IL-17 | AJ493595 | CTCCGATCCCTTATTCTCCTC | AAGCGGTTGTGGTCCTCAT | 62 |
| TGF-β4 | M31160 | CGGGACGGATGAGAAGAAC | CGGCCCACGTAGTAAATGAT | 60 |
| JAK1 | XM_015290965 | TGCACCGTGACTTAGCAGCAAG | TCTGAATCAAGCATTCTGGAGCATACC | 60 |
| JAK2 | XM_015280061 | TCGCTATGGCATTATTCG | GTGGGGTTTGGTCCTTTT | 60 |
| STAT1 | XM_015289392 | TAAAGAGGGAGCAATCAC | ATCAGGGAAAGTAACAGC | 60 |
| STAT5 | XM_015299590 | TCCCACCTGGAGGATTCA | TTCTTCAGCACCTCCATCAC | 60 |
| STAT6 | XM_015274736 | GCAACCTCTACCCCAACA | TCCCTTTCGCTTTCCACT | 62 |
| Adh-1 | NM_204577 | GAAGGAGCTGGGATTGTG | GCTGCATTCTCCACACTG | 58 |
| RALDH-2 | AF064253 | CAAGACATGAACCCATCG | GAGCTGGAGCAATCTTCC | 60 |
| RALDH-3 | AF246710 | AGGCAGCATTCCAGAGAG | TCAGCCAGCTTGTGTAGG | 60 |
| RAR-α | NM_204536 | AGGAGCTGATCGAGAAGG | GAGCTGTTGTTCGTGGTG | 60 |
| RAR-β | NM_205326 | GCATCAGTGCAAAAGGTG | TGTCAGTGGTTCGTGTCC | 60 |
| RAR-γ | NM_205294 | GATGAAGATCACCGACCTG | TCCTCCTCGAACATCTCG | 60 |
| RXR-α | NM_204536 | GATGCGAGACATGCAGATG | GTCGGGGTATTTGTGCTTG | 60 |
| RXR-γ | NM_205294 | CCAAGACGGAGGC ATACAG | GGAGCGATGGGAGAAGGAT | 60 |
| Item | Ctrl | VitA | NE | VitA + NE | SEM | p-Values | ||
|---|---|---|---|---|---|---|---|---|
| VitA | NE | VitA × NE | ||||||
| Total bilirubin (mg/dL) | 1.14 | 1.03 | 1.07 | 1.06 | 0.02 | 0.145 | 0.665 | 0.263 |
| Glucose (mg/dL) | 255.23 | 253.46 | 245.45 | 236.21 | 2.31 | 0.204 | 0.003 | 0.386 |
| Total glyceride (mg/dL) | 44.54 | 55.71 | 36.92 | 32.70 | 2.53 | 0.453 | 0.002 | 0.100 |
| Total cholesterol (mg/dL) | 118.42 | 126.26 | 117.29 | 119.65 | 1.72 | 0.141 | 0.261 | 0.426 |
| High-density lipoprotein (mg/dL) | 108.26 | 112.61 | 108.57 | 110.99 | 1.63 | 0.310 | 0.844 | 0.771 |
| Low-density lipoprotein (mg/dL) | 19.49 | 25.74 | 19.22 | 21.44 | 0.71 | 0.002 | 0.077 | 0.119 |
| Calcium (mg/dL) | 10.89 | 11.03 | 10.49 | 10.57 | 0.10 | 0.564 | 0.036 | 0.872 |
| Phosphorus (mg/dL) | 6.56 | 5.68 | 5.98 | 5.37 | 0.11 | <0.001 | 0.015 | 0.452 |
| Uric acid (mg/dL) | 287.74 | 229.71 | 239.22 | 174.56 | 13.12 | 0.016 | 0.040 | 0.893 |
| Aspartate aminotransferase (U/L) | 46.92 | 64.21 | 54.09 | 65.23 | 2.35 | 0.002 | 0.348 | 0.479 |
| Alanine aminotransferase (U/L) | 0.93 | 0.85 | 0.79 | 0.79 | 0.05 | 0.689 | 0.325 | 0.689 |
| Alkaline phosphatase (U/L) | 4322.9 ab | 4478.5 ab | 5287.9 a | 3825.0 b | 190.71 | 0.078 | 0.670 | 0.030 |
| Glutamyl transpeptidase (U/L) | 19.43 | 21.64 | 18.79 | 20.29 | 0.62 | 0.141 | 0.424 | 0.775 |
| Creatine kinase (U/L) | 2454.5 | 2583.2 | 2017.9 | 2970.4 | 138.68 | 0.050 | 0.927 | 0.132 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, S.; He, L.; Zhang, Y.; Niu, J.; Li, C.; Zhang, Z.; Li, P.; Ding, B. Effects of Vitamin A on Immune Responses and Vitamin A Metabolism in Broiler Chickens Challenged with Necrotic Enteritis. Life 2023, 13, 1122. https://doi.org/10.3390/life13051122
Guo S, He L, Zhang Y, Niu J, Li C, Zhang Z, Li P, Ding B. Effects of Vitamin A on Immune Responses and Vitamin A Metabolism in Broiler Chickens Challenged with Necrotic Enteritis. Life. 2023; 13(5):1122. https://doi.org/10.3390/life13051122
Chicago/Turabian StyleGuo, Shuangshuang, Lai He, Yuanke Zhang, Junlong Niu, Changwu Li, Zhengfan Zhang, Peng Li, and Binying Ding. 2023. "Effects of Vitamin A on Immune Responses and Vitamin A Metabolism in Broiler Chickens Challenged with Necrotic Enteritis" Life 13, no. 5: 1122. https://doi.org/10.3390/life13051122