
WNT16 Regulation of the Articular Chondrocyte Phenotype in Mice

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chondrocyte Culture
2.2. Gene Expression
2.3. Proliferation Assay
2.4. Ex Vivo Joint Culture and Histology
2.5. Ex Vivo Joint Culture and Gene Expression
2.6. Statistical Analysis
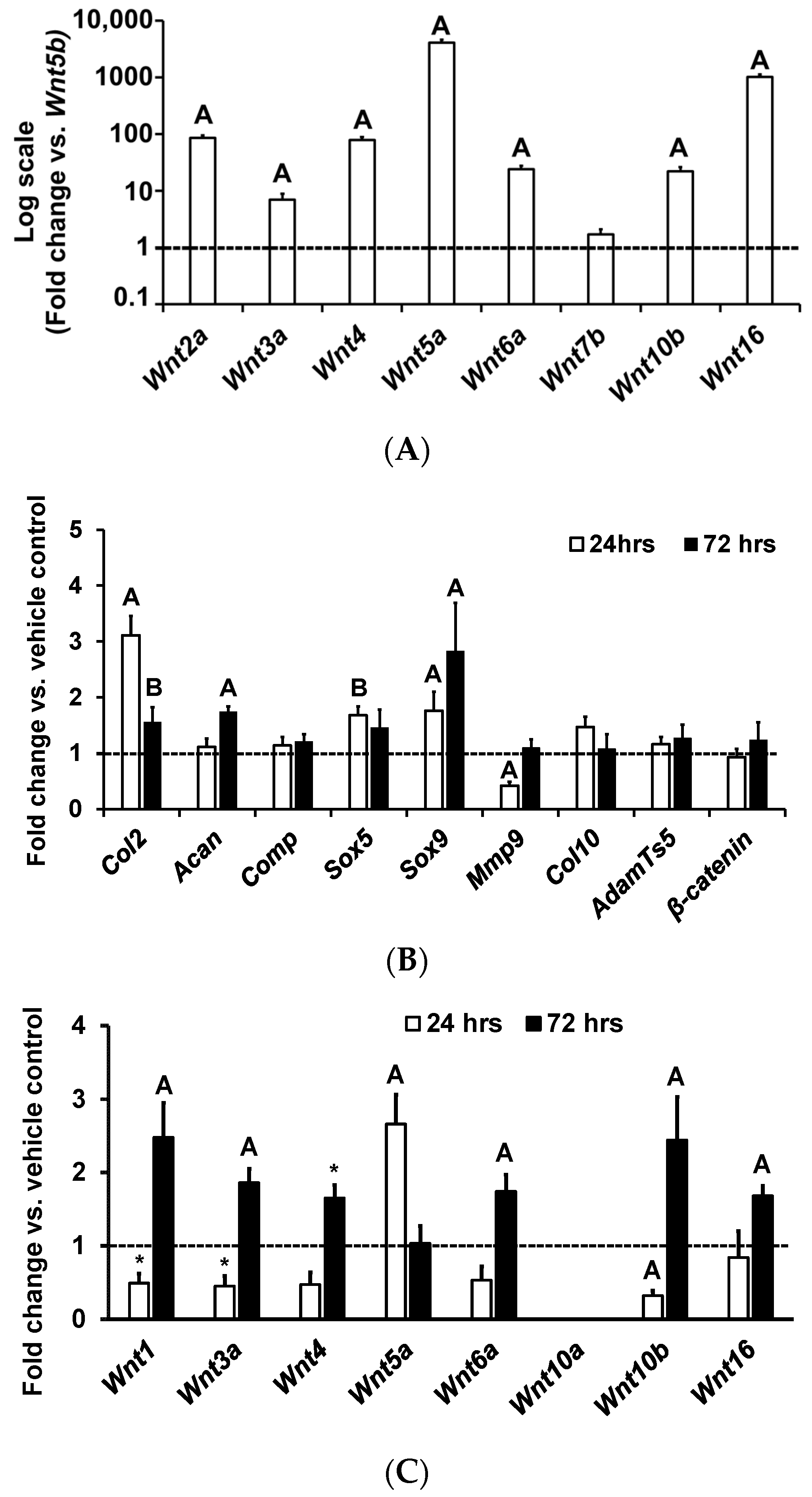
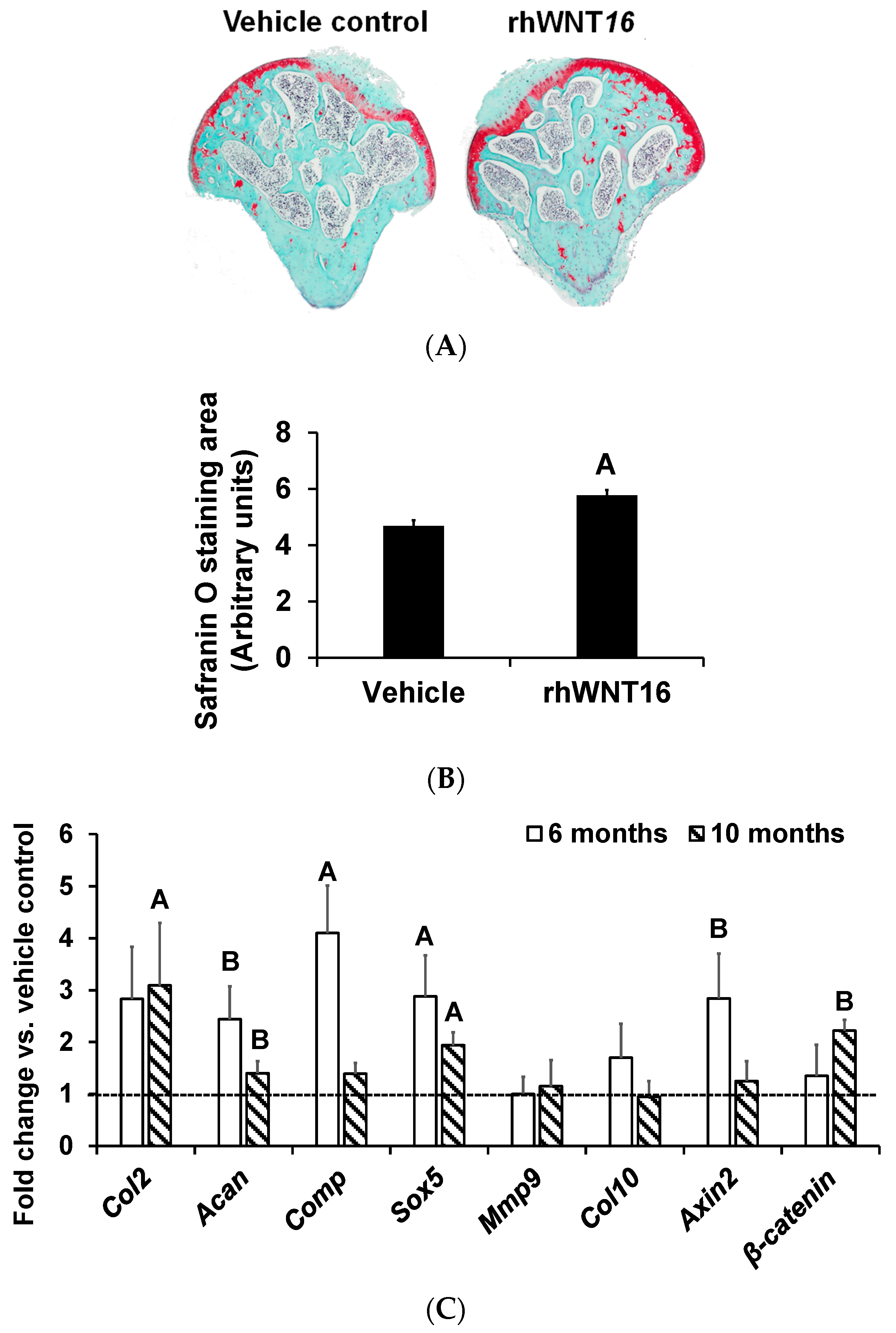
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zichella, L.; Gramolini, R. Autotransfusion during cesarean section. Am. J. Obstet. Gynecol. 1990, 162, 295. [Google Scholar] [CrossRef] [PubMed]
- Buckwalter, J.A.; Brown, T.D. Joint injury, repair, and remodeling: Roles in post-traumatic osteoarthritis. Clin. Orthop. Relat. Res. 2004, 426, 7–16. [Google Scholar] [CrossRef]
- Chen, D.; Shen, J.; Zhao, W.; Wang, T.; Han, L.; Hamilton, J.L.; Im, H.J. Osteoarthritis: Toward a comprehensive understanding of pathological mechanism. Bone Res. 2017, 5, 16044. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.E. Osteoarthritis year in review 2017: Clinical. Osteoarthr. Cartil. 2018, 26, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Xing, D.; Xu, Y.; Liu, Q.; Ke, Y.; Wang, B.; Li, Z.; Lin, J. Osteoarthritis and all-cause mortality in worldwide populations: Grading the evidence from a meta-analysis. Sci. Rep. 2016, 6, 24393. [Google Scholar] [CrossRef]
- Grassel, S.; Muschter, D. Recent advances in the treatment of osteoarthritis. F1000Research 2020, 9, 1–17. [Google Scholar] [CrossRef]
- Liu, Y.; Shah, K.M.; Luo, J. Strategies for Articular Cartilage Repair and Regeneration. Front. Bioeng. Biotechnol. 2021, 9, 770655. [Google Scholar] [CrossRef]
- Blom, A.B.; Brockbank, S.M.; van Lent, P.L.; van Beuningen, H.M.; Geurts, J.; Takahashi, N.; van der Kraan, P.M.; van de Loo, F.A.; Schreurs, B.W.; Clements, K.; et al. Involvement of the Wnt signaling pathway in experimental and human osteoarthritis: Prominent role of Wnt-induced signaling protein 1. Arthritis Rheum. 2009, 60, 501–512. [Google Scholar] [CrossRef]
- Dell’accio, F.; De Bari, C.; Eltawil, N.M.; Vanhummelen, P.; Pitzalis, C. Identification of the molecular response of articular cartilage to injury, by microarray screening: Wnt-16 expression and signaling after injury and in osteoarthritis. Arthritis Rheum. 2008, 58, 1410–1421. [Google Scholar] [CrossRef]
- Tornqvist, A.E.; Nilsson, K.H.; Li, L.; Ohlsson, C.; Moverare-Skrtic, S. Induced inactivation of Wnt16 in young adult mice has no impact on osteoarthritis development. PLoS ONE 2022, 17, e0277495. [Google Scholar] [CrossRef]
- Hua, B.; Qiu, J.; Ye, X.; Liu, X. Intra-articular injection of a novel Wnt pathway inhibitor, SM04690, upregulates Wnt16 expression and reduces disease progression in temporomandibular joint osteoarthritis. Bone 2022, 158, 116372. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Yu, J.; Xu, G.; Bao, G.; Zhang, J.; Xue, P.; Jiang, J.; Chen, J.; Chen, C.; Hong, H.; et al. Wnt16 protects chondrocytes from lumbar facet joint osteoarthritis through the Wnt/beta-catenin pathway in low back pain patients. Somatosens. Mot. Res. 2021, 38, 339–346. [Google Scholar] [CrossRef]
- Nalesso, G.; Thomas, B.L.; Sherwood, J.C.; Yu, J.; Addimanda, O.; Eldridge, S.E.; Thorup, A.S.; Dale, L.; Schett, G.; Zwerina, J.; et al. WNT16 antagonises excessive canonical WNT activation and protects cartilage in osteoarthritis. Ann. Rheum. Dis. 2017, 76, 218–226. [Google Scholar] [CrossRef]
- Cheng, S.; Pourteymoor, S.; Alarcon, C.; Mohan, S. Conditional Deletion of the Phd2 Gene in Articular Chondrocytes Accelerates Differentiation and Reduces Articular Cartilage Thickness. Sci. Rep. 2017, 7, 45408. [Google Scholar] [CrossRef] [PubMed]
- Al-Sabah, A.; Stadnik, P.; Gilbert, S.J.; Duance, V.C.; Blain, E.J. Importance of reference gene selection for articular cartilage mechanobiology studies. Osteoarthr. Cartil. 2016, 24, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.X.; Zhao, R.L.; Song, W.; Chu, H.R.; Li, M.; Song, S.Y.; Li, G.Z.; Liang, D.C. Selection of suitable reference genes for normalization of quantitative real-time PCR in cartilage tissue injury and repair in rabbits. Int. J. Mol. Sci. 2012, 13, 14344–14355. [Google Scholar] [CrossRef]
- Mohan, S.; Wergedal, J.E.; Das, S.; Kesavan, C. Conditional disruption of miR17-92 cluster in collagen type I-producing osteoblasts results in reduced periosteal bone formation and bone anabolic response to exercise. Physiol. Genom. 2015, 47, 33–43. [Google Scholar] [CrossRef]
- Tong, W.; Zeng, Y.; Chow, D.H.K.; Yeung, W.; Xu, J.; Deng, Y.; Chen, S.; Zhao, H.; Zhang, X.; Ho, K.K.; et al. Wnt16 attenuates osteoarthritis progression through a PCP/JNK-mTORC1-PTHrP cascade. Ann. Rheum. Dis. 2019, 78, 551–561. [Google Scholar] [CrossRef]
- Yan, H.; Hu, Y.; Akk, A.; Rai, M.F.; Pan, H.; Wickline, S.A.; Pham, C.T.N. Induction of WNT16 via Peptide-mRNA Nanoparticle-Based Delivery Maintains Cartilage Homeostasis. Pharmaceutics 2020, 12, 73. [Google Scholar] [CrossRef]
- Ozeki, N.; Mogi, M.; Hase, N.; Hiyama, T.; Yamaguchi, H.; Kawai, R.; Kondo, A.; Nakata, K. Wnt16 Signaling Is Required for IL-1beta-Induced Matrix Metalloproteinase-13-Regulated Proliferation of Human Stem Cell-Derived Osteoblastic Cells. Int. J. Mol. Sci. 2016, 17, 221. [Google Scholar] [CrossRef]
- Dreier, R. Hypertrophic differentiation of chondrocytes in osteoarthritis: The developmental aspect of degenerative joint disorders. Arthritis Res. Ther. 2010, 12, 216. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Dong, S. The Signaling Pathways Involved in Chondrocyte Differentiation and Hypertrophic Differentiation. Stem Cells Int. 2016, 2016, 2470351. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohan, S.; Pourteymoor, S.; Kesavan, C. WNT16 Regulation of the Articular Chondrocyte Phenotype in Mice. Life 2023, 13, 878. https://doi.org/10.3390/life13040878
Mohan S, Pourteymoor S, Kesavan C. WNT16 Regulation of the Articular Chondrocyte Phenotype in Mice. Life. 2023; 13(4):878. https://doi.org/10.3390/life13040878
Chicago/Turabian StyleMohan, Subburaman, Shelia Pourteymoor, and Chandrasekhar Kesavan. 2023. "WNT16 Regulation of the Articular Chondrocyte Phenotype in Mice" Life 13, no. 4: 878. https://doi.org/10.3390/life13040878