
Bone-Marrow-Derived Mesenchymal Stem Cells Attenuate Behavioral and Cognitive Dysfunction after Subarachnoid Hemorrhage via HMGB1–RAGE Axis Mediation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental SAH Model
2.2. BMSC Culture
2.3. BMSC Administration
2.4. RNA Isolation and Quantitative RT-PCR
2.5. Western Blots
2.6. Brain Water Content
2.7. Behavioral and Cognitive Dysfunction
2.8. Statistical Analysis
3. Results
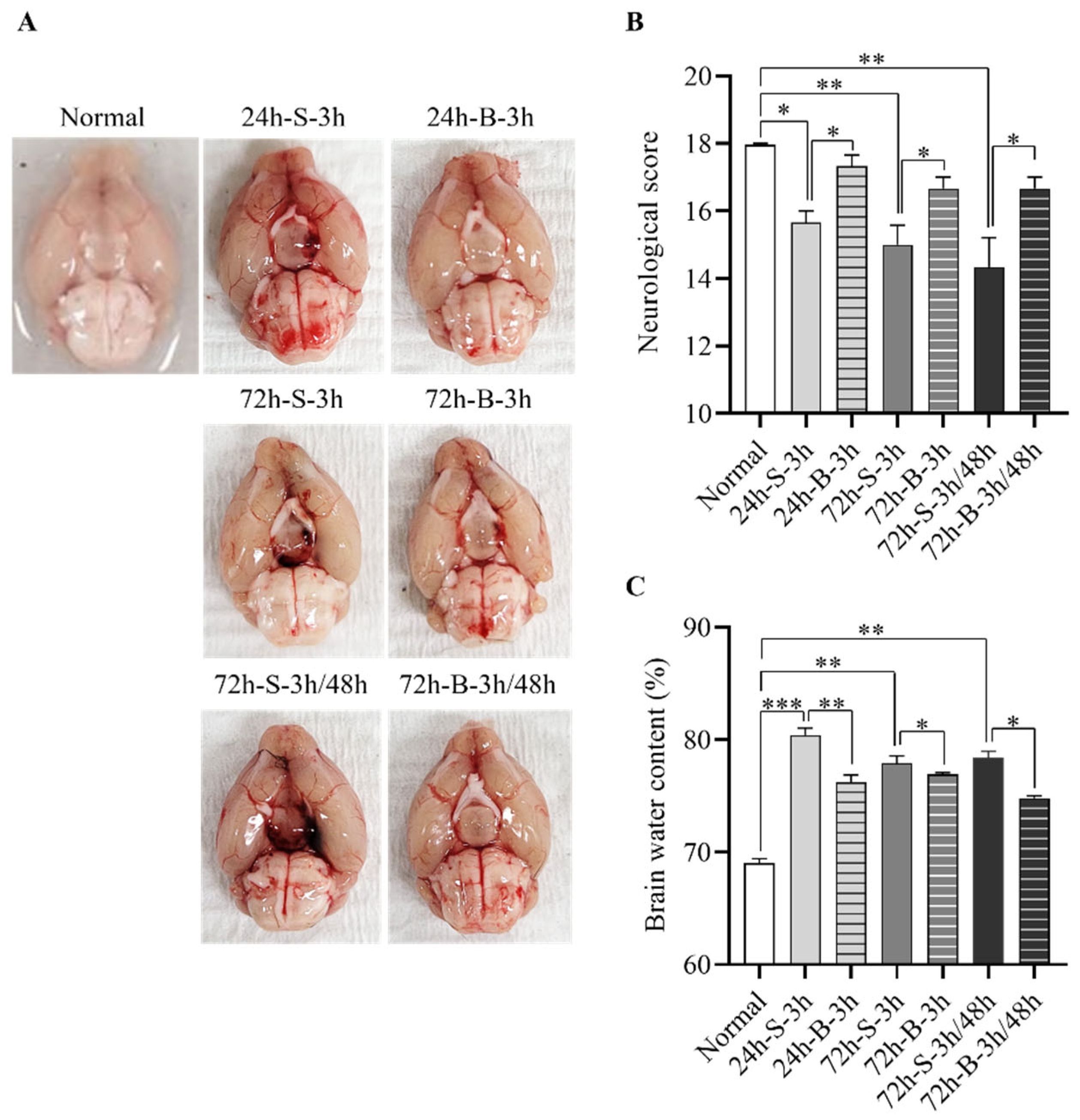
3.1. Effect of BMSC on the In Vivo SAH Model
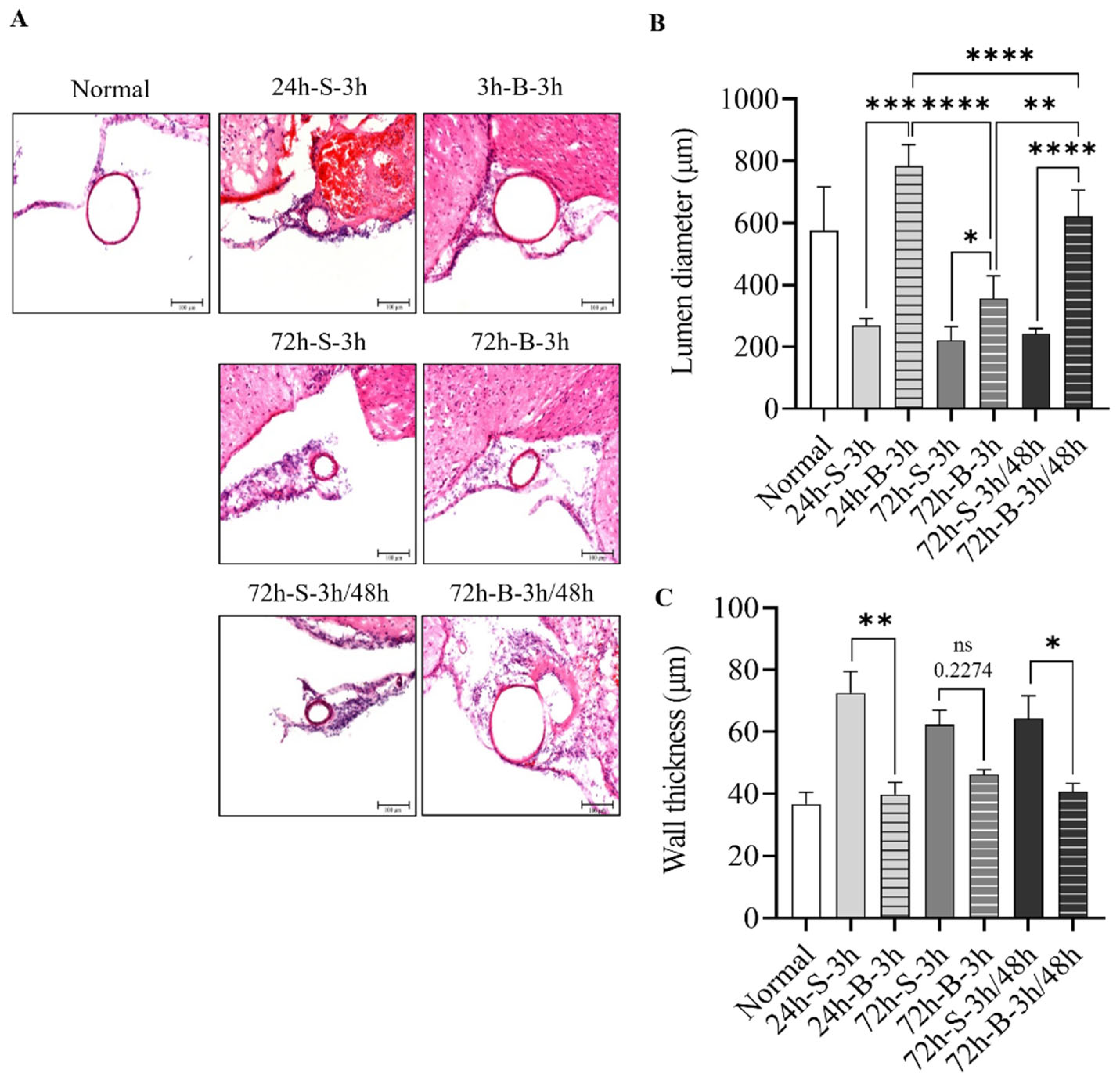
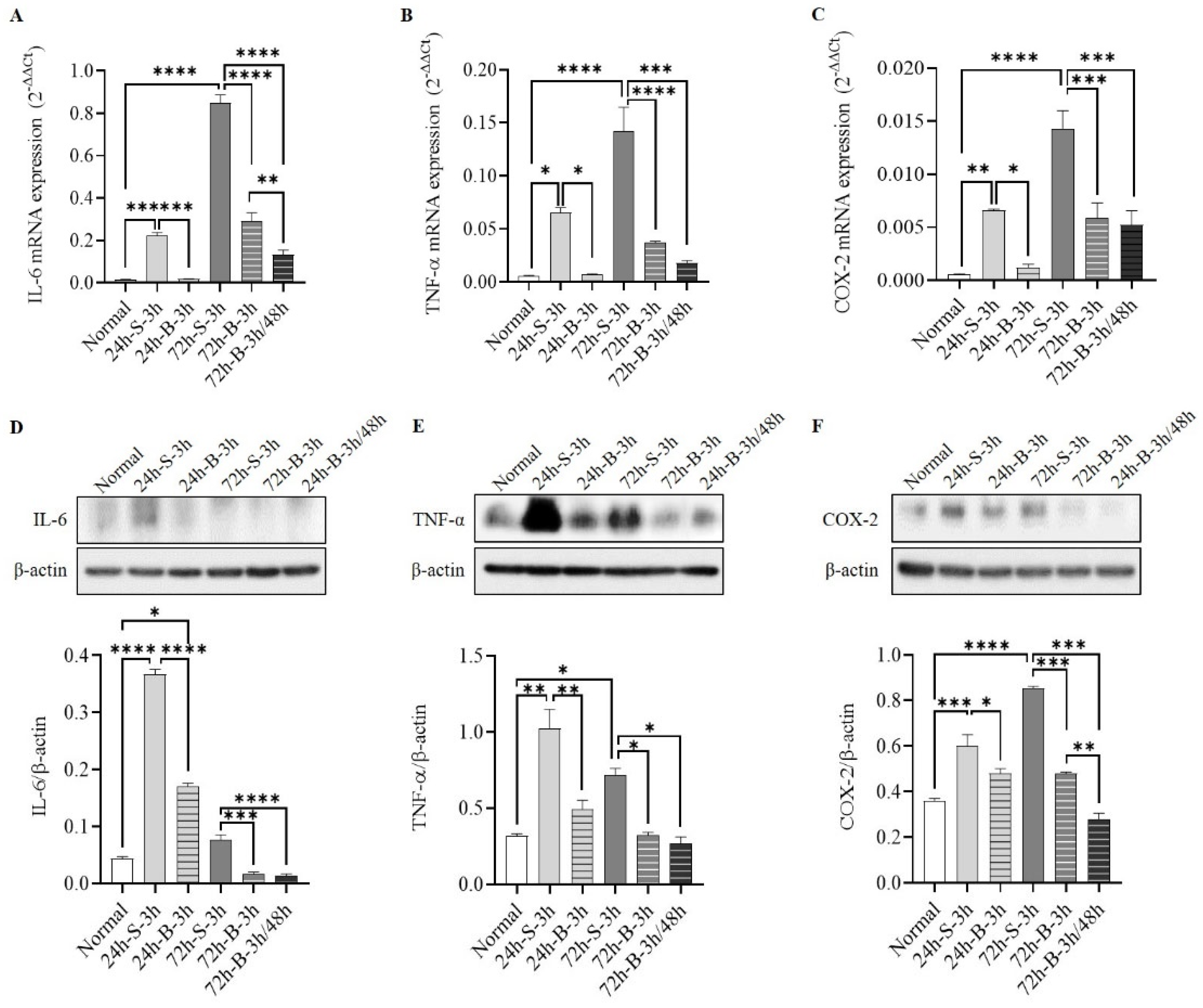
3.2. Histological Examination, Neuronal Apoptosis, and Inflammation
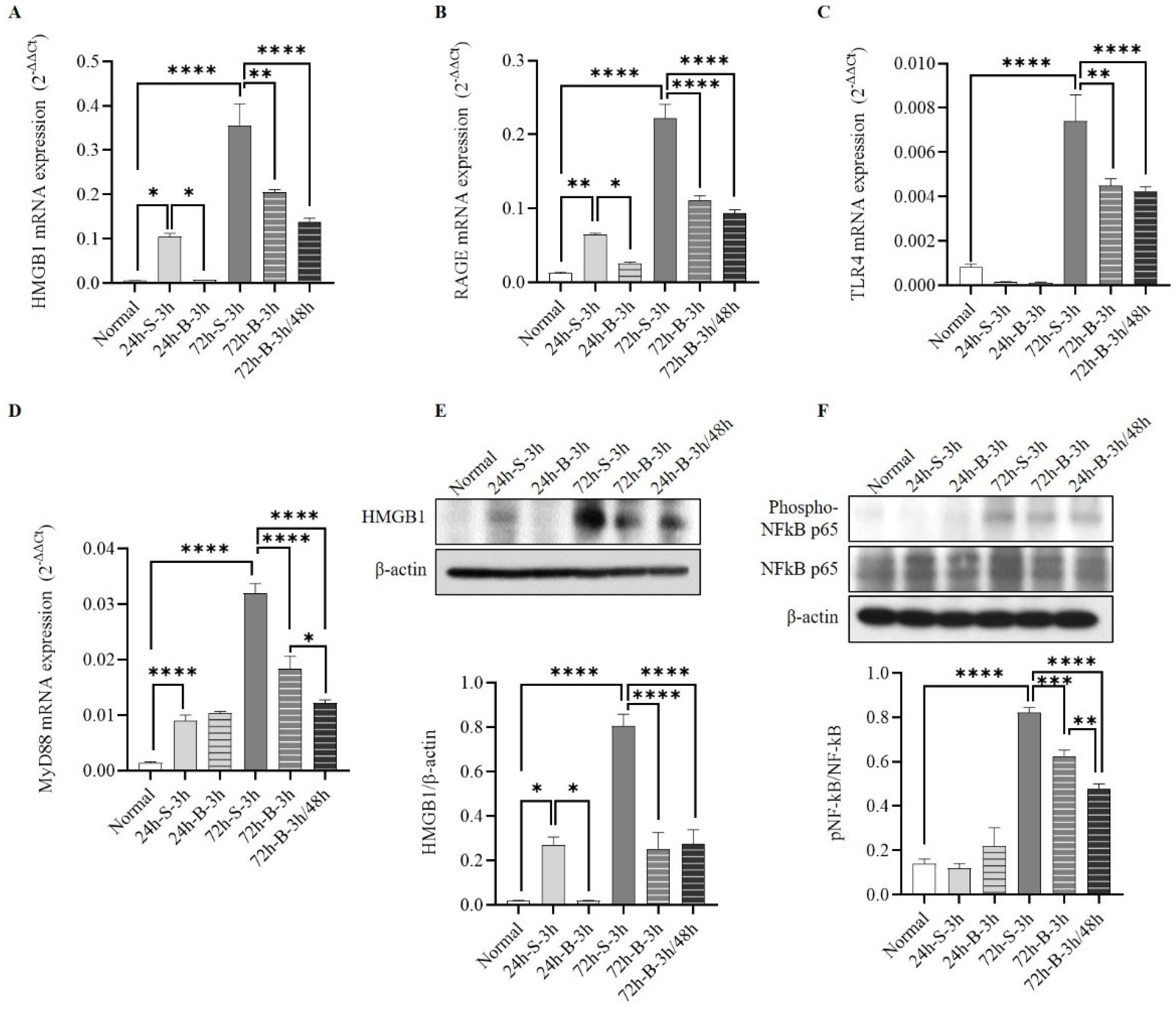
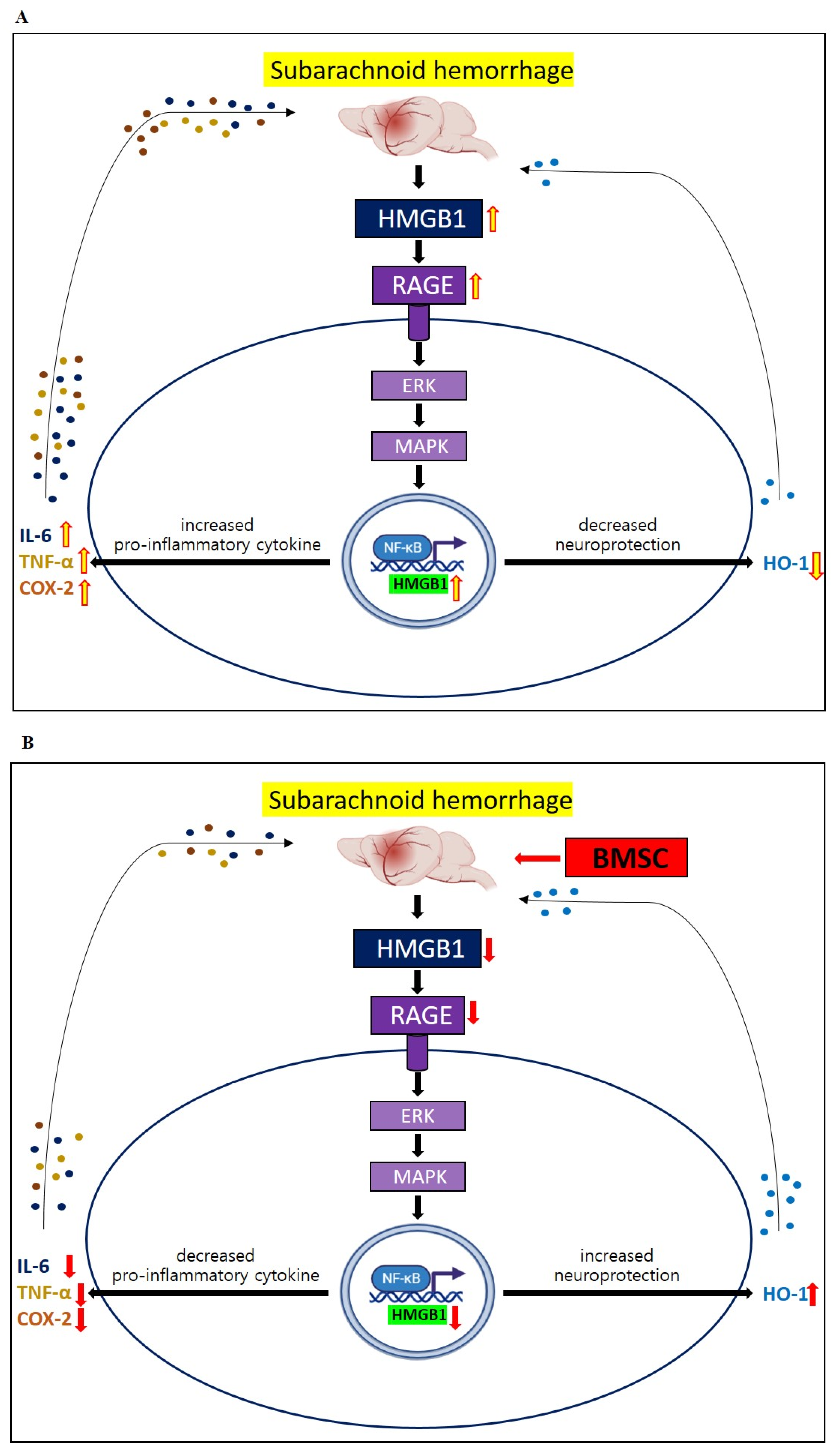
3.3. Changes in HMGB1–RAGE Axis Inflammation
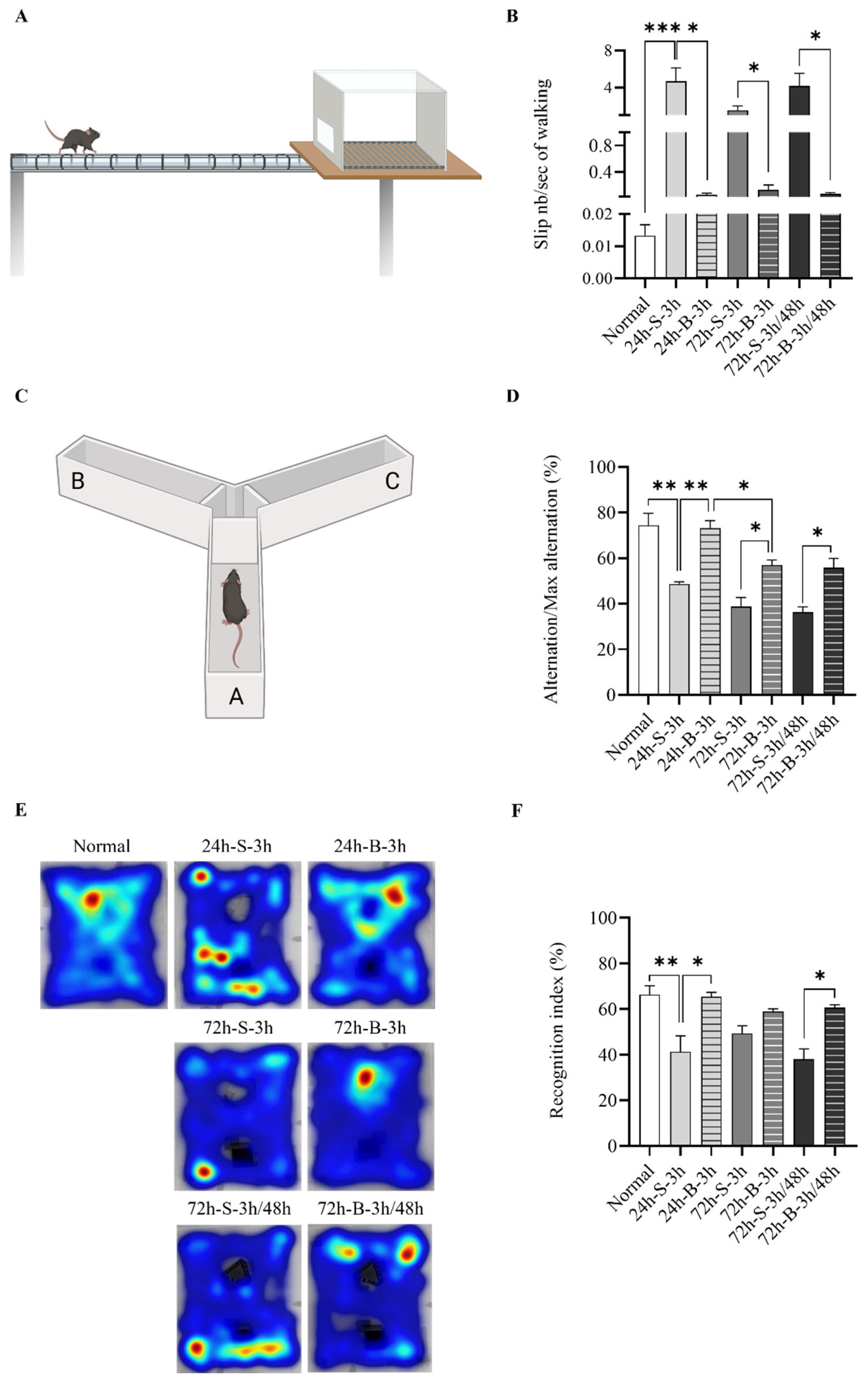
3.4. Behavior and Cognition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Linn, F.H.; Rinkel, G.J.; Algra, A.; van Gijn, J. Incidence of subarachnoid hemorrhage: Role of region, year, and rate of computed tomography: A meta-analysis. Stroke 1996, 27, 625–629. [Google Scholar] [CrossRef]
- Mourelo-Farina, M.; Pertega, S.; Galeiras, R. A Model for Prediction of In-Hospital Mortality in Patients with Subarachnoid Hemorrhage. Neurocrit. Care 2021, 34, 508–518. [Google Scholar] [CrossRef]
- Ravnik, J.; Starovasnik, B.; Sesok, S.; Pirtosek, Z.; Svigelj, V.; Bunc, G.; Bosnjak, R. Long-term cognitive deficits in patients with good outcomes after aneurysmal subarachnoid hemorrhage from anterior communicating artery. Croat. Med. J. 2006, 47, 253–263. [Google Scholar]
- Eagles, M.E.; Tso, M.K.; Macdonald, R.L. Cognitive Impairment, Functional Outcome, and Delayed Cerebral Ischemia After Aneurysmal Subarachnoid Hemorrhage. World Neurosurg. 2019, 124, e558–e562. [Google Scholar] [CrossRef]
- Haug Nordenmark, T.; Karic, T.; Sorteberg, W.; Sorteberg, A. Predictors of cognitive function in the acute phase after aneurysmal subarachnoid hemorrhage. Acta Neurochir. 2019, 161, 177–184. [Google Scholar] [CrossRef]
- Chung, D.Y.; Oka, F.; Jin, G.; Harriott, A.; Kura, S.; Aykan, S.A.; Qin, T.; Edmiston, W.J.; Lee, H.; Yassen, M.A.; et al. Subarachnoid hemorrhage leads to early and persistent functional connectivity and behavioral changes in mice. J. Cereb. Blood Flow Metab. 2021, 41, 975–985. [Google Scholar] [CrossRef]
- Xiong, L.; Sun, L.; Zhang, Y.; Peng, J.; Yan, J.; Liu, X. Exosomes from Bone Marrow Mesenchymal Stem Cells Can Alleviate Early Brain Injury After Subarachnoid Hemorrhage Through miRNA129-5p-HMGB1 Pathway. Stem. Cells Dev. 2020, 29, 212–221. [Google Scholar] [CrossRef] [Green Version]
- Shichinohe, H.; Kuroda, S.; Sugiyama, T.; Ito, M.; Kawabori, M. Bone marrow stromal cell transplantation attenuates cognitive dysfunction due to chronic cerebral ischemia in rats. Dement. Geriatr. Cogn. Disord. 2010, 30, 293–301. [Google Scholar] [CrossRef]
- Kim, J.T.; Youn, D.H.; Kim, B.J.; Rhim, J.K.; Jeon, J.P. Recent Stem Cell Research on Hemorrhagic Stroke: An Update. J. Korean Neurosurg. Soc. 2022, 65, 161–172. [Google Scholar] [CrossRef]
- Skok, M. Mesenchymal stem cells as a potential therapeutic tool to cure cognitive impairment caused by neuroinflammation. World J. Stem. Cells. 2021, 13, 1072–1083. [Google Scholar] [CrossRef]
- Khalili, M.A.; Anvari, M.; Hekmati-Moghadam, S.H.; Sadeghian-Nodoushan, F.; Fesahat, F.; Miresmaeili, S.M. Therapeutic benefit of intravenous transplantation of mesenchymal stem cells after experimental subarachnoid hemorrhage in rats. J. Stroke Cerebrovasc. Dis. 2012, 21, 445–451. [Google Scholar] [CrossRef]
- Liu, W.; Li, R.; Yin, J.; Guo, S.; Chen, Y.; Fan, H.; Li, G.; Li, Z.; Li, X.; Zhang, X.; et al. Mesenchymal stem cells alleviate the early brain injury of subarachnoid hemorrhage partly by suppression of Notch1-dependent neuroinflammation: Involvement of Botch. J. Neuroinflamm. 2019, 16, 8. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A. Editorial: Neuroinflammation and Cognition. Front. Aging Neurosci. 2018, 10, 413. [Google Scholar] [CrossRef] [Green Version]
- Paudel, Y.N.; Shaikh, M.F.; Chakraborti, A.; Kumari, Y.; Aledo-Serrano, A.; Aleksovska, K.; Alvim, M.K.M.; Othman, I. HMGB1: A Common Biomarker and Potential Target for TBI, Neuroinflammation, Epilepsy, and Cognitive Dysfunction. Front. Neurosci. 2018, 12, 628. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Liu, K.; Wake, H.; Teshigawara, K.; Mori, S.; Nishibori, M. Anti-high mobility group box-1 (HMGB1) antibody inhibits hemorrhage-induced brain injury and improved neurological deficits in rats. Sci. Rep. 2017, 7, 46243. [Google Scholar] [CrossRef] [Green Version]
- Paudel, Y.N.; Angelopoulou, E.; Piperi, C.; Othman, I.; Aamir, K.; Shaikh, M.F. Impact of HMGB1, RAGE, and TLR4 in Alzheimer’s Disease (AD): From Risk Factors to Therapeutic Targeting. Cells 2020, 9, 383. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.H.; Lee, D.H.; Song, J. HMGB1 signaling pathway in diabetes-related dementia: Blood-brain barrier breakdown, brain insulin resistance, and Abeta accumulation. Biomed. Pharmacother. 2022, 150, 112933. [Google Scholar] [CrossRef]
- Schuller, K.; Buhler, D.; Plesnila, N. A murine model of subarachnoid hemorrhage. J. Vis. Exp. 2013, 81, e50845. [Google Scholar]
- Sugawara, T.; Ayer, R.; Jadhav, V.; Zhang, J.H. A new grading system evaluating bleeding scale in filament perforation subarachnoid hemorrhage rat model. J. Neurosci. Methods 2008, 167, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Garcia, J.H.; Wagner, S.; Liu, K.F.; Hu, X.J. Neurological deficit and extent of neuronal necrosis attributable to middle cerebral artery occlusion in rats. Statistical validation. Stroke 1995, 26, 627–634. [Google Scholar] [CrossRef]
- Chu, D.T.; Phuong, T.N.T.; Tien, N.L.B.; Tran, D.K.; Thanh, V.V.; Quang, T.L.; Truong, D.T.; Pham, V.H.; Ngoc, V.T.N.; Chu-Dinh, T.; et al. An Update on the Progress of Isolation, Culture, Storage, and Clinical Application of Human Bone Marrow Mesenchymal Stem/Stromal Cells. Int. J. Mol. Sci. 2020, 21, 708. [Google Scholar] [CrossRef] [Green Version]
- Delorme, B.; Charbord, P. Culture and characterization of human bone marrow mesenchymal stem cells. Methods Mol. Med. 2007, 140, 67–81. [Google Scholar]
- Youn, D.H.; Tran, N.M.; Kim, B.J.; Kim, Y.; Jeon, J.P.; Yoo, H. Shape effect of cerium oxide nanoparticles on mild traumatic brain injury. Sci. Rep. 2021, 11, 15571. [Google Scholar] [CrossRef]
- Pedard, M.; El Amki, M.; Lefevre-Scelles, A.; Compere, V.; Castel, H. Double Direct Injection of Blood into the Cisterna Magna as a Model of Subarachnoid Hemorrhage. J. Vis. Exp. 2020, 162, 975–985. [Google Scholar] [CrossRef]
- Kim, C.K.; Kim, T.; Choi, I.Y.; Soh, M.; Kim, D.; Kim, Y.J.; Park, H.; Park, S.P.; Park, S.; Yu, T.; et al. Ceria nanoparticles that can protect against ischemic stroke. Angew. Chem. Int. Ed. Engl. 2012, 51, 11039–11043. [Google Scholar] [CrossRef]
- Bae, Y.H.; Joo, H.; Bae, J.; Hyeon, S.J.; Her, S.; Ko, E.; Choi, H.G.; Ryu, H.; Hur, E.; Lee, B.D. Brain injury induces HIF-1alpha-dependent transcriptional activation of LRRK2 that exacerbates brain damage. Cell Death Dis. 2018, 9, 1125. [Google Scholar] [CrossRef] [Green Version]
- Han, S.W.; Kim, B.J.; Kim, T.Y.; Lim, S.H.; Youn, D.H.; Hong, E.P.; Rhim, J.K.; Park, J.J.; Lee, J.J.; Cho, Y.J.; et al. Association of Haptoglobin Phenotype With Neurological and Cognitive Outcomes in Patients With Subarachnoid Hemorrhage. Front. Aging Neurosci. 2022, 14, 819628. [Google Scholar] [CrossRef]
- Andrzejewska, A.; Dabrowska, S.; Lukomska, B.; Janowski, M. Mesenchymal Stem Cells for Neurological Disorders. Adv. Sci. 2021, 8, 2002944. [Google Scholar] [CrossRef]
- Nakano, M.; Kubota, K.; Kobayashi, E.; Chikenji, T.S.; Saito, Y.; Konari, N.; Fujimiya, M. Bone Marrow-Derived Mesenchymal Stem Cells Improve Cognitive Impairment in an Alzheimer’s Disease Model by Increasing the Expression of microRNA-146a in hippocampus. Sci. Rep. 2020, 10, 10772. [Google Scholar] [CrossRef]
- Zhang, S.; Lachance, B.B.; Moiz, B.; Jia, X. Optimizing Stem Cell Therapy after Ischemic Brain Injury. J. Stroke 2020, 22, 286–305. [Google Scholar] [CrossRef]
- Lee, P.H.; Kim, J.W.; Bang, O.Y.; Ahn, Y.H.; Joo, I.S.; Huh, K. Autologous mesenchymal stem cell therapy delays the progression of neurological deficits in patients with multiple system atrophy. Clin. Pharmacol. Ther. 2008, 83, 723–730. [Google Scholar] [CrossRef]
- Lee, J.K.; Jin, H.K.; Endo, S.; Schuchman, E.H.; Carter, J.E.; Bae, J.S. Intracerebral transplantation of bone marrow-derived mesenchymal stem cells reduces amyloid-beta deposition and rescues memory deficits in Alzheimer’s disease mice by modulation of immune responses. Stem Cells 2010, 28, 329–343. [Google Scholar] [CrossRef]
- Al-Mufti, F.; Amuluru, K.; Smith, B.; Damodara, N.; El-Ghanem, M.; Singh, I.P.; Dangayach, N.; Gandhi, C.D. Emerging Markers of Early Brain Injury and Delayed Cerebral Ischemia in Aneurysmal Subarachnoid Hemorrhage. World Neurosurg. 2017, 107, 148–159. [Google Scholar] [CrossRef]
- Jung, J.W.; Kwon, M.; Choi, J.C.; Shin, J.W.; Park, I.W.; Choi, B.W.; Kim, J.Y. Familial occurrence of pulmonary embolism after intravenous, adipose tissue-derived stem cell therapy. Yonsei Med. J. 2013, 54, 1293–1296. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Deng, H.; Hu, Y.; Pan, C.; Wu, G.; Li, Q.; Tang, Z. Adipose-derived mesenchymal stem cells stereotactic transplantation alleviate brain edema from intracerebral hemorrhage. J. Cell Biochem. 2019, 120, 14372–14382. [Google Scholar] [CrossRef]
- Brunet, M.C.; Chen, S.H.; Khandelwal, P.; Hare, J.M.; Starke, R.M.; Peterson, E.C.; Yavagal, D.R. Intravenous Stem Cell Therapy for High-Grade Aneurysmal Subarachnoid Hemorrhage: Case Report and Literature Review. World Neurosurg. 2019, 128, 573–575. [Google Scholar] [CrossRef]
- Haruma, J.; Teshigawara, K.; Hishikawa, T.; Wang, D.; Liu, K.; Wake, H.; Mori, S.; Takahashi, H.K.; Sugiu, K.; Date, I.; et al. Anti-high mobility group box-1 (HMGB1) antibody attenuates delayed cerebral vasospasm and brain injury after subarachnoid hemorrhage in rats. Sci. Rep. 2016, 6, 37755. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Hu, L.; Jiang, J.; Li, H.; Wu, Q.; Ooi, K.; Wang, J.; Feng, Y.; Zhu, D.; Xia, C. HMGB1/RAGE axis mediates stress-induced RVLM neuroinflammation in mice via impairing mitophagy flux in microglia. J. Neuroinflamm. 2020, 17, 15. [Google Scholar] [CrossRef]
- Gallyas, F., Jr. Acute changes of pro-inflammatory markers and corticosterone in experimental subarachnoid haemorrhage: A prerequisite for severity assessment. PLoS ONE 2019, 14, e0220467. [Google Scholar]
- Muhammad, S.; Grote, A. Interleukin 6 and Aneurysmal Subarachnoid Hemorrhage. A Narrative Review. Int. J. Mol. Sci. 2021, 22, 4133. [Google Scholar]
- Ayer, R.; Hadhav, V.; Sugawara, T.; Zhang, J.H. The neuroprotective effects of cyclooxygenase-2 inhibition in a mouse model of aneurysmal subarachnoid hemorrhage. Acta Neurochir. Suppl. 2011, 111, 145–149. [Google Scholar]
- Paudel, Y.N.; Angelopoulou, E.; Piperi, C.; Othman, I.; Shaikh, M.F. HMGB1-Mediated Neuroinflammatory Responses in Brain injuries: Potential Mechanisms and Therapeutic Opportunities. Int. J. Mol. Sci. 2020, 21, 4609. [Google Scholar] [CrossRef]
- Wang, L.; Geng, G.; Zhu, T.; Chen, W.; Li, X.; Gu, J.; Jiang, E. Progress in Research on TLR4-Mediated Inflammatory Response Mechanisms in Brain Injury after Subarachnoid Hemorrhage. Cells 2022, 23, 3781. [Google Scholar] [CrossRef]
- Ahmed, H.; Khan, M.A.; Kahlert, U.D.; Niemelä, M.; Hänggi, D.; Chaudhry, S.R.; Muhammad, S. Role of Adaptor Protein Myeloid Differentiation 88 (MyD88) in Post-Subarachnoid Hemorrhage: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 4185. [Google Scholar] [CrossRef]
- Shao, A.; Wu, H.; Hong, Y.; Tu, S.; Sun, X.; Wu, Q.; Zhao, Q.; Zhang, J.; Sheng, J. Hydrogen-Rich Saline Attenuated Subarachnoid Hemorrhage-Induced Early Brain Injury in Rats by Suppressing Inflammatory Response: Possible Involvement of NF-kB Pathway and NLRP3 Inflammasome. Mol. Neurobiol. 2016, 53, 3462–3467. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, H.; Youn, D.H.; Park, J.J.; Jeon, J.P. Bone-Marrow-Derived Mesenchymal Stem Cells Attenuate Behavioral and Cognitive Dysfunction after Subarachnoid Hemorrhage via HMGB1–RAGE Axis Mediation. Life 2023, 13, 881. https://doi.org/10.3390/life13040881
Jung H, Youn DH, Park JJ, Jeon JP. Bone-Marrow-Derived Mesenchymal Stem Cells Attenuate Behavioral and Cognitive Dysfunction after Subarachnoid Hemorrhage via HMGB1–RAGE Axis Mediation. Life. 2023; 13(4):881. https://doi.org/10.3390/life13040881
Chicago/Turabian StyleJung, Harry, Dong Hyuk Youn, Jeong Jin Park, and Jin Pyeong Jeon. 2023. "Bone-Marrow-Derived Mesenchymal Stem Cells Attenuate Behavioral and Cognitive Dysfunction after Subarachnoid Hemorrhage via HMGB1–RAGE Axis Mediation" Life 13, no. 4: 881. https://doi.org/10.3390/life13040881