
Chlorophyll Meter: A Precision Agricultural Decision-Making Tool for Nutrient Supply in Durum Wheat (Triticum turgidum L.) Cultivation under Drought Conditions
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
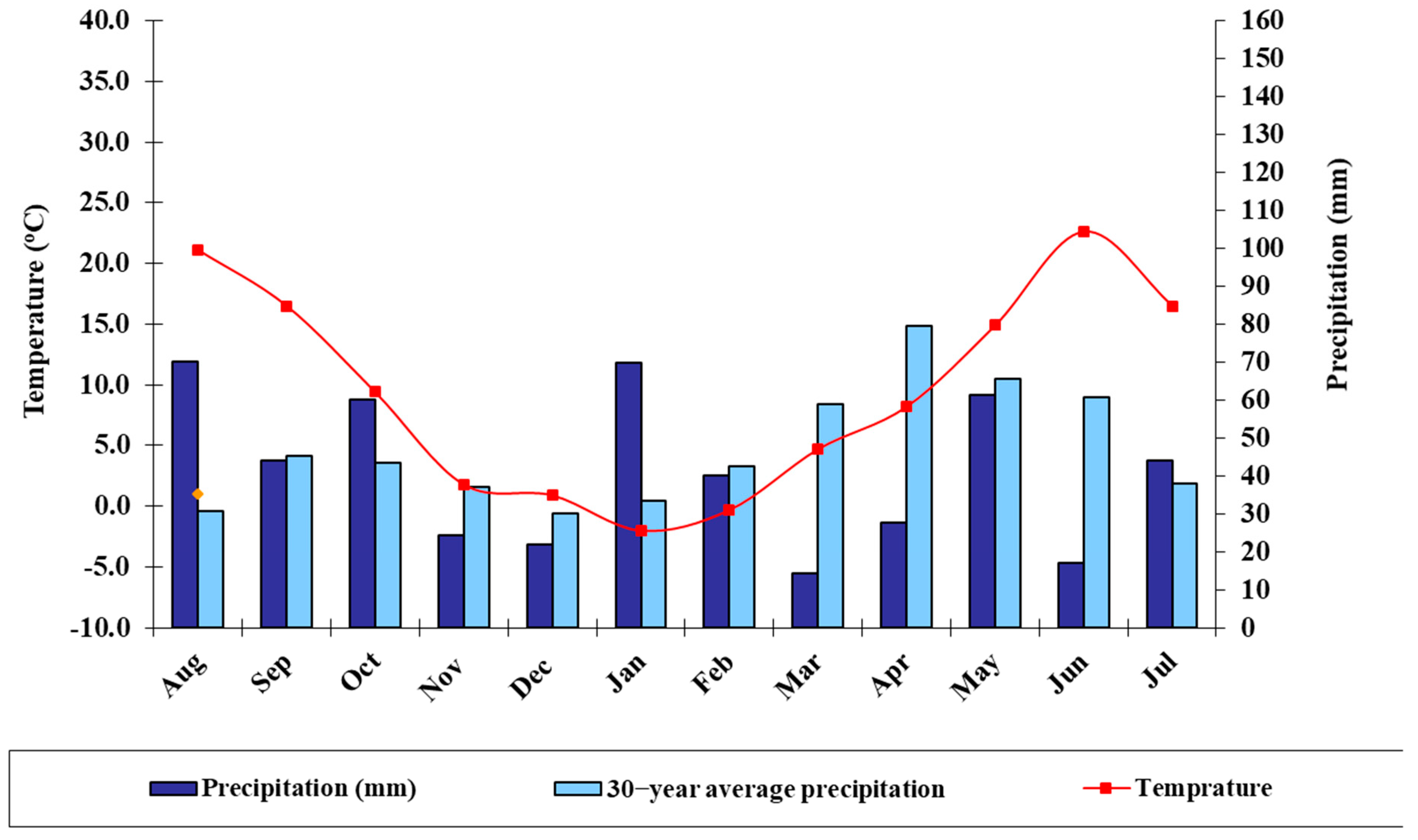
2.1. Genetic Materials and Study Site Description
2.2. Experimental Design and Treatments
2.3. Data Collection
- A = constant;
- B = constant;
- Ior = current from red detectors with sample in place;
- Ir = current from red infrared detectors with sample in place;
- Iof = currents from red detectors with no sample;
- If = currents from infrared detectors with no sample
- AMC = Actual (obtained) Grain Moisture Content (%)
- SMC = Standard Moisture Content
2.4. Statistical Data Analysis
3. Results
3.1. Genetic Regulation and Stability of Chlorophyll Content
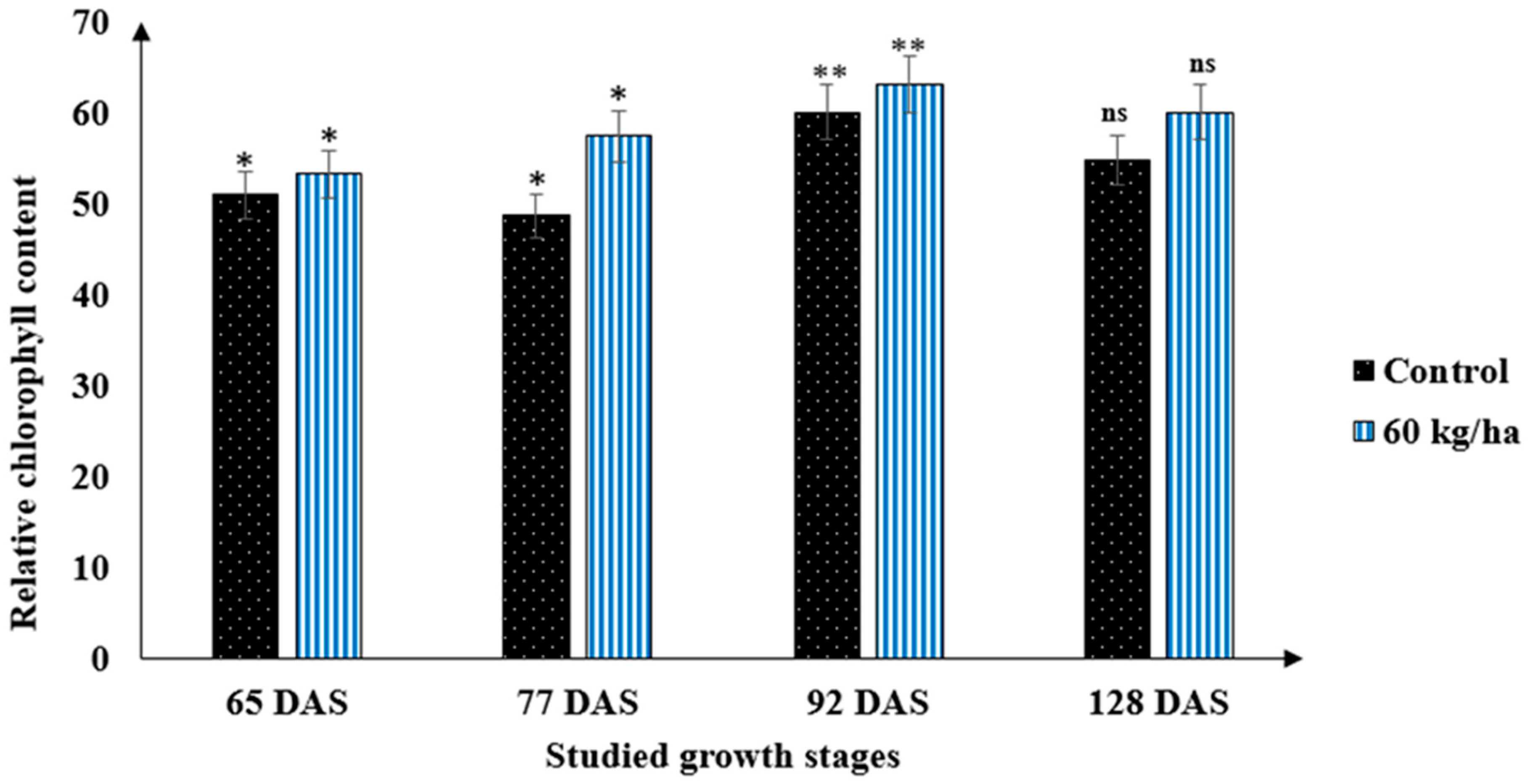
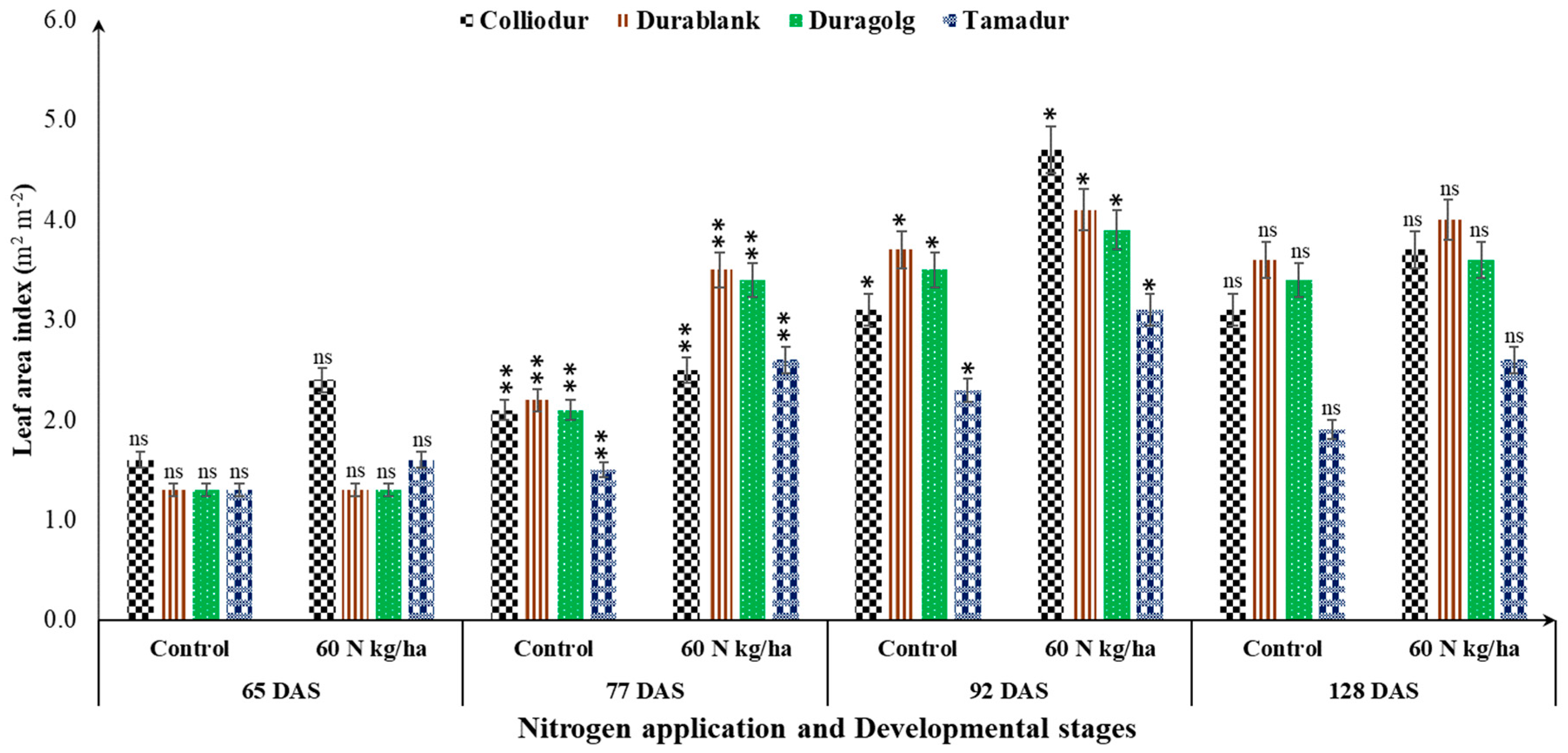
3.2. Interaction of Durum Wheat Varieties and Nutrient Supply under Drought Conditions
3.3. Dynamics in SPAD Values (Chlorophyll) and Its Contribution to Grain Yield
3.4. Evaluation of Durum Wheat Varities Nitrgen Status Using Chlorophyll Meters
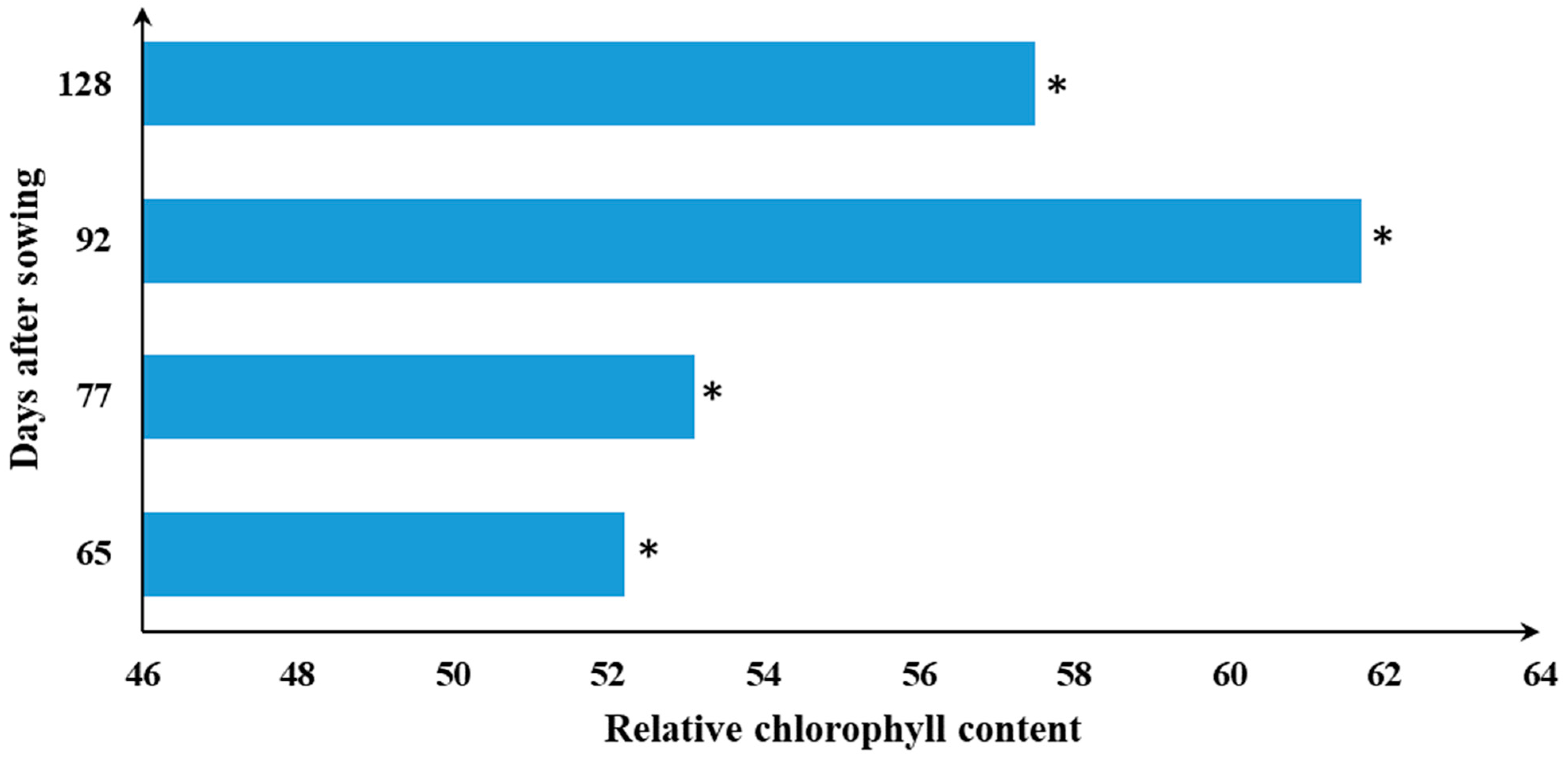
3.5. Developmental Stages Determine the SPAD Readings
3.6. Interaction Effect of Nitrogen × Sulfur × Zinc on Spikes Density (m2)
3.7. Canopy Reflectance Sensor-Based Fertilizer Management
3.8. Nutrient Supply Influences Grain Protein Content (%)
3.9. Relationships between SPAD Readings, Yield, and Morphological Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beres, B.L.; Rahmani, E.; Clarke, J.M.; Grassini, P.; Pozniak, C.J.; Geddes, C.M.; Porker, K.D.; May, W.E.; Ransom, J.K. A Systematic Review of Durum Wheat: Enhancing Production Systems by Exploring Genotype, Environment, and Management (G × E × M) Synergies. Front. Plant Sci. 2020, 11, 568657. [Google Scholar] [CrossRef]
- Ayadi, S.; Karmous, C.; Chamekh, Z.; Hammami, Z.; Baraket, M.; Esposito, S.; Rezgui, S.; Trifa, Y. Effects of nitrogen rates on grain yield and nitrogen agronomic efficiency of durum wheat genotypes under different environments. Ann. Appl. Biol. 2015, 168, 264–273. [Google Scholar] [CrossRef] [Green Version]
- Croft, H.; Chen, J.M. Leaf Pigment Content. In Comprehensive Remote Sensing; Liang, S., Ed.; Elsevier: Oxford, UK, 2018; pp. 117–142. [Google Scholar]
- Tang, Y.L.; Wang, J.H.; Huang, J.F.; Wang, R.C.; He, Q.X. Variation law of hyperspectral data and chlorophyll and ca-rotenoid for rice in mature process. Trans. Chin. Soc. Agric. Eng. 2013, 19, 167–173. [Google Scholar]
- Peng, J.; Feng, Y.; Wang, X.; Li, J.; Xu, G.; Phonenasay, S.; Luo, Q.; Han, Z.; Lu, W. Effects of nitrogen application rate on the photosynthetic pigment, leaf fluorescence characteristics, and yield of indica hybrid rice and their interrelations. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liao, Y.; Liu, W. High nitrogen application rate and planting density reduce wheat grain yield by reducing filling rate of inferior grain in middle spikelets. Crop. J. 2020, 9, 412–426. [Google Scholar] [CrossRef]
- Efernández-Marín, B.; Eartetxe, U.; Barrutia, O.; Esteban, R.; Hernandez, A.; Garcia-Plazaola, J.I. Opening Pandora’s box: Cause and impact of errors on plant pigment studies. Front. Plant Sci. 2015, 6, 148. [Google Scholar] [CrossRef]
- Yuan, Z.; Ata-Ul-Karim, S.T.; Cao, Q.; Lu, Z.; Cao, W.; Zhu, Y.; Liu, X. Indicators for diagnosing nitrogen status of rice based on chlorophyll meter readings. Field Crop. Res. 2015, 185, 12–20. [Google Scholar] [CrossRef]
- Ata-Ul-Karim, S.T.; Cao, Q.; Zhu, Y.; Tang, L.; Rehmani, M.I.A.; Cao, W. Non-destructive Assessment of Plant Nitrogen Parameters Using Leaf Chlorophyll Measurements in Rice. Front. Plant Sci. 2016, 7, 1829. [Google Scholar] [CrossRef] [Green Version]
- Wan, W.; Zhao, Y.; Xu, J.; Liu, K.; Guan, S.; Chai, Y.; Cui, H.; Wu, P.; Diao, M. Reducing and Delaying Nitrogen Recommended by Leaf Critical SPAD Value Was More Suitable for Nitrogen Utilization of Spring Wheat under a New Type of Drip-Irrigated System. Agronomy 2022, 12, 2331. [Google Scholar] [CrossRef]
- Walsh, O.S.; Torrion, J.A.; Liang, X.; Shafian, S.; Yang, R.; Belmont, K.M.; McClintick-Chess, J.R. Grain yield, quality, and spectral characteristics of wheat grown under varied nitrogen and irrigation. Agrosyst. Geosci. Environ. 2020, 3, e20104. [Google Scholar] [CrossRef]
- Hou, W.; Shen, J.; Xu, W.; Khan, M.R.; Wang, Y.; Zhou, X.; Gao, Q.; Murtaza, B.; Zhang, Z. Recommended nitrogen rates and the verification of effects based on leaf SPAD readings of rice. PeerJ 2021, 9, e12107. [Google Scholar] [CrossRef] [PubMed]
- Kalaji, H.M.; Dąbrowski, P.; Cetner, M.D.; Samborska, I.A.; Łukasik, I.; Brestic, M.; Zivcak, M.; Tomasz, H.; Mojski, J.; Kociel, H.; et al. A comparison between different chlorophyll content meters under nutrient deficiency conditions. J. Plant Nutr. 2016, 40, 1024–1034. [Google Scholar] [CrossRef]
- Ahmad, N.; Malagoli, M.; Wirtz, M.; Hell, R. Drought stress in maize causes differential acclimation responses of glutathione and sulfur metabolism in leaves and roots. BMC Plant Biol. 2016, 16, 247. [Google Scholar] [CrossRef] [Green Version]
- Xiong, D.; Chen, J.; Yu, T.; Gao, W.; Ling, X.; Li, Y.; Peng, S.; Huang, J. SPAD-based leaf nitrogen estimation is impacted by environmental factors and crop leaf characteristics. Sci. Rep. 2015, 5, 13389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorentini, M.; Zenobi, S.; Giorgini, E.; Basili, D.; Conti, C.; Pro, C.; Monaci, E.; Orsini, R. Nitrogen and chlorophyll status determination in durum wheat as influenced by fertilization and soil management: Preliminary results. PLoS ONE 2019, 14, e0225126. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Swain, D.K.; Jha, M.K.; Tewari, V.K.; Bohra, A. Optimizing chlorophyll meter (SPAD) reading to allow efficient nitrogen use in rice and wheat under rice-wheat cropping system in eastern India. Plant Prod. Sci. 2020, 23, 270–285. [Google Scholar] [CrossRef] [Green Version]
- Usmani, M.M.; Nawaz, F.; Majeed, S.; Shehzad, M.A.; Ahmad, K.S.; Akhtar, G.; Aqib, M.; Shabbir, R.N. Sulfate-mediated Drought Tolerance in Maize Involves Regulation at Physiological and Biochemical Levels. Sci. Rep. 2020, 10, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Juhasz, A.; Islam, S.; Diepeveen, D.; Zhang, J.; Wang, P.; Ma, W. Impact of mid-season sulphur deficiency on wheat nitrogen metabolism and biosynthesis of grain protein. Sci. Rep. 2018, 8, 2499. [Google Scholar] [CrossRef] [Green Version]
- Wilson, T.L.; Mary, J.; Guttieri, N.O.; Nelson, A.F.; Michael, T. Nitrogen and sulfur effects on hard winter wheat quality and asparagine concentration. J. Cereal Sci. 2020, 93, 102969. [Google Scholar] [CrossRef]
- Tao, Z.; Chang, X.; Wang, D.; Wang, Y.; Ma, S.; Yang, Y.; Zhao, G. Effects of sulfur fertilization and short-term high temperature on wheat grain production and wheat flour proteins. Crop. J. 2018, 6, 413–425. [Google Scholar] [CrossRef]
- Lee, B.-R.; Zaman, R.; Avice, J.-C.; Ourry, A.; Kim, T.-H. Sulfur Use Efficiency Is a Significant Determinant of Drought Stress Tolerance in Relation to Photosynthetic Activity in Brassica napus Cultivars. Front. Plant Sci. 2016, 7, 459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klikocka, H.; Marks, M.; Marx, M. Sulphur and Nitrogen Fertilization as a Potential Means of Agronomic Biofortification to Improve the Content and Uptake of Microelements in Spring Wheat Grain DM. J. Chem. 2018, 2018, 9326820. [Google Scholar] [CrossRef]
- Batista, K.; Monteiro, F.A. Nitrogen and sulphur in Marandu grass: Relationship between supply and concentration in leaf tissues. Sci. Agricola 2007, 64, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Csajbók, J.; Pepó, P.; Kutasy, E. Photosynthetic and Agronomic Traits of Winter Barley (Hordeum vulgare L.) Varieties. Agronomy 2020, 10, 1999. [Google Scholar] [CrossRef]
- Melash, A.A.; Ábrahám, B. Integrated nutrient supply and varietal difference influence grain yield and yield related physio-morphological traits of durum wheat (Triticum turgidum L.) varieties under drought condition. Acta Agrar. Debr. 2022, 1, 111–121. [Google Scholar] [CrossRef]
- Wood, C.W.; Reeves, D.W.; Himelrick, D.G. Relationships between chlorophyll meter readings and leaf chlorophyll concentration, N status, and crop yield: A review. Proc. Agron. Soc. N. Z. 1993, 23, 1–9. [Google Scholar]
- Badu-Apraku, B.; Fakorede, M.A.; Menkir, A.; Sanogo, D. Conduct and Management of Maize Field Trials; IITA: Ibadan, Nigeria, 2012; p. 59. [Google Scholar]
- Panek, E.; Gozdowski, D.; Stępień, M.; Samborski, S.; Ruciński, D.; Buszke, B. Within-Field Relationships between Satellite-Derived Vegetation Indices, Grain Yield and Spike Number of Winter Wheat and Triticale. Agronomy 2020, 10, 1842. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Payne, R.; Murray, D.; Harding, S.; Baird, D.; Soutar, D. Introduction to Genstat for Windows, 18th ed.; VSN International, 2 Amberside, Wood Lane, Hemel Hempstead: Hertfordshire, UK, 2015; pp. 1–144. Available online: http://www.genstat.co.uk/ (accessed on 6 March 2023).
- Melash, A.A.; Ábrahám, B. Barriers and levers to enhance end-use functional properties of durum wheat (Triticum turgidum L.) grain: An agronomic implication. Heliyon 2022, 8, e09542. [Google Scholar] [CrossRef]
- Araus, J.L.; Slafer, G.; Reynolds, M.P.; Royo, C. Plant Breeding and Drought in C3 Cereals: What Should We Breed For? Ann. Bot. 2002, 89, 925–940. [Google Scholar] [CrossRef]
- Hamblin, J.; Stefanova, K.; Angessa, T.T. Variation in Chlorophyll Content per Unit Leaf Area in Spring Wheat and Implications for Selection in Segregating Material. PLoS ONE 2014, 9, e92529. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.; Ali, A.M. Using Hand-Held Chlorophyll Meters and Canopy Reflectance Sensors for Fertilizer Nitrogen Management in Cereals in Small Farms in Developing Countries. Sensors 2020, 20, 1127. [Google Scholar] [CrossRef] [Green Version]
- Yasir, T.A.; Wasaya, A.; Hussain, M.; Ijaz, M.; Farooq, M.; Farooq, O.; Nawaz, A.; Hu, Y.-G. Evaluation of physiological markers for assessing drought tolerance and yield potential in bread wheat. Physiol. Mol. Biol. Plants 2019, 25, 1163–1174. [Google Scholar] [CrossRef]
- Ghannoum, O.; Evans, J.R.; Chow, W.S.; Andrews, T.J.; Conroy, J.P.; von Caemmerer, S. Faster Rubisco Is the Key to Superior Nitrogen-Use Efficiency in NADP-Malic Enzyme Relative to NAD-Malic Enzyme C4 Grasses. Plant Physiol. 2005, 137, 638–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanahan, J.; Kitchen, N.; Raun, W.; Schepers, J. Responsive in-season nitrogen management for cereals. Comput. Electron. Agric. 2008, 61, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Yuan, X.; Yan, F.; Xiang, K.; Wu, Y.; Zhang, Q.; Wang, Z.; He, L.; Fan, P.; Yang, Z.; et al. Nitrogen Application Rate Affects the Accumulation of Carbohydrates in Functional Leaves and Grains to Improve Grain Filling and Reduce the Occurrence of Chalkiness. Front. Plant Sci. 2022, 13, 921130. [Google Scholar] [CrossRef]
- Veres, S.; Zsombik, L.; Juhász, C. Regulation Effect of Different Water Supply to the Nitrogen and Carbon Metabolism. In Abiotic and Biotic Stress in Plants; de Oliveira, A.B., Ed.; IntechOpen: London, UK, 2019; p. 261. [Google Scholar]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Naeem, M.; Naeem, M.S.; Ahmad, R.; Ahmad, R. Foliar-applied calcium induces drought stress tolerance in maize by manipulating osmolyte accumulation and antioxidative responses. Pak. J. Bot. 2017, 49, 427–434. [Google Scholar]
- Kizilgeci, F.; Yildirim, M.; Islam, M.; Ratnasekera, D.; Iqbal, M.; Sabagh, A. Normalized Difference Vegetation Index and Chlorophyll Content for Precision Nitrogen Management in Durum Wheat Cultivars under Semi-Arid Conditions. Sustainability 2021, 13, 3725. [Google Scholar] [CrossRef]
- Raza, M.A.; Feng, L.Y.; Iqbal, N.; Manaf, A.; Bin Khalid, M.H.; Rehman, S.U.; Wasaya, A.; Ansar, M.; Billah, M.; Yang, F.; et al. Effect of Sulphur Application on Photosynthesis and Biomass Accumulation of Sesame Varieties under Rainfed Conditions. Agronomy 2018, 8, 149. [Google Scholar] [CrossRef] [Green Version]
- Sohrabi, U.; Ahmadi, A.; Heidari, G.-R.; Siosemarde, A. Efect of sulfur and zinc fertilizers on physiological characteristics and grain yield of corn (Zea mays L.) under drought stress. Environ. Stresses Crop Sci. 2019, 12, 763–779. [Google Scholar] [CrossRef]
- Noah, D.W.; Mohammed, G.; Edwin, L.; Paul, M.; David, M.; Mohamed, M.; Jerry, J.; James, B.H.; Gina, B.G. Genetic var-iation for plant growth traits in a common wheat population is dominated by known variants and novel QTL. BioRxiv 2020, 12, 16, 422696. [Google Scholar] [CrossRef]
- Panek, E.; Gozdowski, D. Analysis of relationship between cereal yield and NDVI for selected regions of Central Europe based on MODIS satellite data. Remote Sens. Appl. Soc. Environ. 2019, 17, 100286. [Google Scholar] [CrossRef]
- Bożek, K.S.; Żuk-Gołaszewska, K.; Bochenek, A.; Gołaszewski, J.; Kalaji, H.M. Modelling the growth, development and yield of Triticum durum Desf under the changes of climatic conditions in north-eastern Europe. Sci. Rep. 2021, 11, 21753. [Google Scholar] [CrossRef]
- Din, M.; Zheng, W.; Rashid, M.; Wang, S.; Shi, Z. Evaluating Hyperspectral Vegetation Indices for Leaf Area Index Estimation of Oryza sativa L. at Diverse Phenological Stages. Front. Plant Sci. 2017, 8, 820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abid, M.; Tian, Z.; Ata-Ul-Karim, S.T.; Cui, Y.; Liu, Y.; Zahoor, R.; Jiang, D.; Dai, T. Nitrogen Nutrition Improves the Potential of Wheat (Triticum aestivum L.) to Alleviate the Effects of Drought Stress during Vegetative Growth Periods. Front. Plant Sci. 2016, 7, 981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Würschum, T.; Leiser, W.L.; Langer, S.M.; Tucker, M.R.; Longin, C.F.H. Phenotypic and genetic analysis of spike and kernel characteristics in wheat reveals long-term genetic trends of grain yield components. Theor. Appl. Genet. 2018, 131, 2071–2084. [Google Scholar] [CrossRef]
- Lin, X.; Li, P.; Shang, Y.; Liu, S.; Wang, S.; Hu, X.; Wang, D. Spike formation and seed setting of the main stem and tillers under post-jointing drought in winter wheat. J. Agron. Crop. Sci. 2020, 206, 694–710. [Google Scholar] [CrossRef]
- Kadkol, G.P.; Sissons, M. Durum Wheat Overview, Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Sissons, M. Role of durum wheat composition on the quality of pasta and bread. Food 2008, 2, 75–90. [Google Scholar]
- Gaines, C.S. Prediction of Sugar-Snap Cookie Diameter Using Sucrose Solvent Retention Capacity, Milling Softness, and Flour Protein Content. Cereal Chem. 2004, 81, 549–552. [Google Scholar] [CrossRef]
- Tipples, K.; Kilborn, R.; Preston, K. Bread wheat quality defined. In Wheat Production, Properties and Quality, 1st ed.; Bushuk, W., Rasper, V.F., Eds.; Blackie Academic and Professional: Glasgow, Scotland, 1994; pp. 25–36. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N Rate (kg/ha) | Varieties | Chlorophyll Measured at Different Stages | SD (m2) | GPC (%) | GY (t/ha) | |||
|---|---|---|---|---|---|---|---|---|
| 65 DAS | 77 DAS | 92 DAS | 128 DAS | |||||
| Control | Colliodur | 49.0 | 49.0 | 56.8 | 52.3 | 216 | 14.6 | 6.61 |
| Durablank | 54.3 | 56.6 | 65.3 | 62.7 | 216 | 16.6 | 6.85 | |
| Duragold | 53.6 | 55.0 | 65.9 | 61.6 | 206 | 16.3 | 7.00 | |
| Tamadur | 51.8 | 51.8 | 58.8 | 53.5 | 154 | 15.3 | 4.95 | |
| LSD (0.05) | 1.19 | 1.03 | 1.36 | 1.93 | 10.3 | 0.61 | 0.24 | |
| CV (%) | 3.40 | 2.90 | 3.30 | 5.0 | 7.7 | 2.8 | 3.0 | |
| N Rate (kg/ha) | Varieties | Chlorophyll Measured at Different Stages | SD (m2) | GPC (%) | GY (t/ha) | |||
|---|---|---|---|---|---|---|---|---|
| 65 DAS | 77 DAS | 92 DAS | 128 DAS | |||||
| Control | Colliodur | 47.2 | 43.0 | 54.9 | 48.6 | 190.3 | 13.1 | 6.51 |
| Durablank | 53.8 | 52.8 | 64.7 | 61.3 | 211.2 | 15.8 | 6.80 | |
| Duragold | 53.3 | 51.7 | 64.9 | 59.8 | 200.2 | 15.4 | 6.53 | |
| Tamadur | 49.8 | 47.3 | 56.0 | 50.1 | 137.9 | 15.2 | 4.01 | |
| 60 kg/ha | Colliodur | 50.7 | 55.0 | 58.7 | 55.9 | 241.9 | 16.2 | 6.70 |
| Durablank | 54.9 | 60.4 | 65.8 | 64.2 | 220.8 | 17.4 | 6.90 | |
| Duragold | 54.0 | 58.3 | 66.9 | 63.3 | 212.7 | 17.3 | 7.45 | |
| Tamadur | 53.8 | 56.4 | 61.6 | 56.9 | 170.3 | 15.3 | 5.89 | |
| LSD (0.05) | 2.34 | 1.36 | 3.69 | 2.99 | 14.52 | 0.72 | 0.96 | |
| CV (%) | 3.40 | 2.90 | 3.30 | 5.00 | 7.70 | 2.80 | 3.00 | |
| Varieties | Chlorophyll Measured at Different Stages | SD (m2) | GPC (%) | GY (t/ha) | |||
|---|---|---|---|---|---|---|---|
| 65 DAS | 77 DAS | 92 DAS | 128 DAS | ||||
| Colliodur | 49.0 | 49.0 | 56.8 | 52.3 | 216 | 14.6 | 6.61 |
| Durablank | 54.3 | 56.6 | 65.3 | 62.7 | 216 | 16.6 | 6.85 |
| Duragold | 53.6 | 55.0 | 65.9 | 61.6 | 206 | 16.3 | 7.00 |
| Tamadur | 51.8 | 51.8 | 58.8 | 53.5 | 154 | 15.3 | 4.95 |
| LSD (0.05) | 1.19 | 1.03 | 1.36 | 1.93 | 10.3 | 0.61 | 0.24 |
| CV (%) | 3.40 | 2.90 | 3.30 | 5.0 | 7.7 | 2.8 | 3.0 |
| Nutrients | Varieties | Chlorophyll Measured at Different Stages | GPC (%) | GY (t/ha) | |||
|---|---|---|---|---|---|---|---|
| SPAD 65 | SPAD 77 | SPAD 95 | SPAD 128 | ||||
| Control | Colliodur | 49.8 | 49.4 | 57.4 | 52.6 | 14.3 | 6.9 |
| Durablank | 53.4 | 55.8 | 65.6 | 62.1 | 15.3 | 6.9 | |
| Duragold | 53.3 | 54.1 | 66.8 | 62.3 | 14.3 | 7.1 | |
| Tamadur | 51.0 | 52.8 | 56.9 | 52.7 | 16.6 | 4.8 | |
| Colliodur | 47.9 | 47.7 | 56.4 | 52.1 | 16.5 | 6.4 | |
| Durablank | 54.3 | 56.4 | 64.3 | 62.5 | 16.8 | 6.9 | |
| Sulfur | Duragold | 55.0 | 55.3 | 65.9 | 61.1 | 16.4 | 6.9 |
| Tamadur | 52.5 | 51.2 | 59.4 | 54.9 | 16.3 | 5.4 | |
| Zinc | Colliodur | 49.2 | 50.0 | 56.6 | 52.1 | 16.4 | 6.5 |
| Durablank | 55.3 | 57.5 | 66.0 | 63.6 | 15.2 | 6.7 | |
| Duragold | 52.7 | 55.6 | 65.0 | 61.4 | 15.1 | 7.0 | |
| Tamadur | 52.0 | 51.6 | 60.1 | 52.9 | 15.7 | 4.7 | |
| LSD (0.05) | 2.1 | ns | 2.5 | 3.12 | 0.7 | 0.4 | |
| CV (%) | 3.4 | 2.9 | 3.3 | 5.0 | 3.1 | 5.5 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melash, A.A.; Bytyqi, B.; Nyandi, M.S.; Vad, A.M.; Ábrahám, É.B. Chlorophyll Meter: A Precision Agricultural Decision-Making Tool for Nutrient Supply in Durum Wheat (Triticum turgidum L.) Cultivation under Drought Conditions. Life 2023, 13, 824. https://doi.org/10.3390/life13030824
Melash AA, Bytyqi B, Nyandi MS, Vad AM, Ábrahám ÉB. Chlorophyll Meter: A Precision Agricultural Decision-Making Tool for Nutrient Supply in Durum Wheat (Triticum turgidum L.) Cultivation under Drought Conditions. Life. 2023; 13(3):824. https://doi.org/10.3390/life13030824
Chicago/Turabian StyleMelash, Anteneh Agezew, Bekir Bytyqi, Muhoja Sylivester Nyandi, Attila Miklós Vad, and Éva Babett Ábrahám. 2023. "Chlorophyll Meter: A Precision Agricultural Decision-Making Tool for Nutrient Supply in Durum Wheat (Triticum turgidum L.) Cultivation under Drought Conditions" Life 13, no. 3: 824. https://doi.org/10.3390/life13030824