

Host–Parasite Coevolution in Primates

Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
| Adaptation | Genetic change that increases fitness (operationally reproductive success) in response to natural selection |
| Co-adaptation | Reciprocal adaptation in two or more ecologically interacting species, such as a host and its parasite (pathogen) or a flowering plant and its pollinator |
| Coevolution | Joint evolution of two or more ecologically interacting species. Often used in the strict sense as an equivalent to co-adaptation |
| Co-diversification | Joint diversification of interacting lineages: (1) early step in the process of co-speciation, and (2) more suitable term than co-speciation in cases where species delimitation is difficult (e.g., bacterial strains, viruses) |
| Co-speciation | Joint speciation of both the host and parasite. Either the host or parasite may speciate slightly after or before the other |
| Evolutionary Arms Race | A struggle between competing sets of coevolving genes, traits, or species, that leads to adaptations and counter-adaptations against each other |
| Fahrenholz’s Rule | Parasite phylogeny mirrors host phylogeny |
| Fitness | Individual reproductive success. Equal to the average contribution of an individual to the gene pool of the next generation(s) |
| Virulence | Parasite (pathogen)-induced reduction in host fitness, and the ability of a parasite to cause disease in and/or death of its hosts (morbidity and mortality) |
2. Coevolution
3. Coevolution and Parasites
4. Virulence
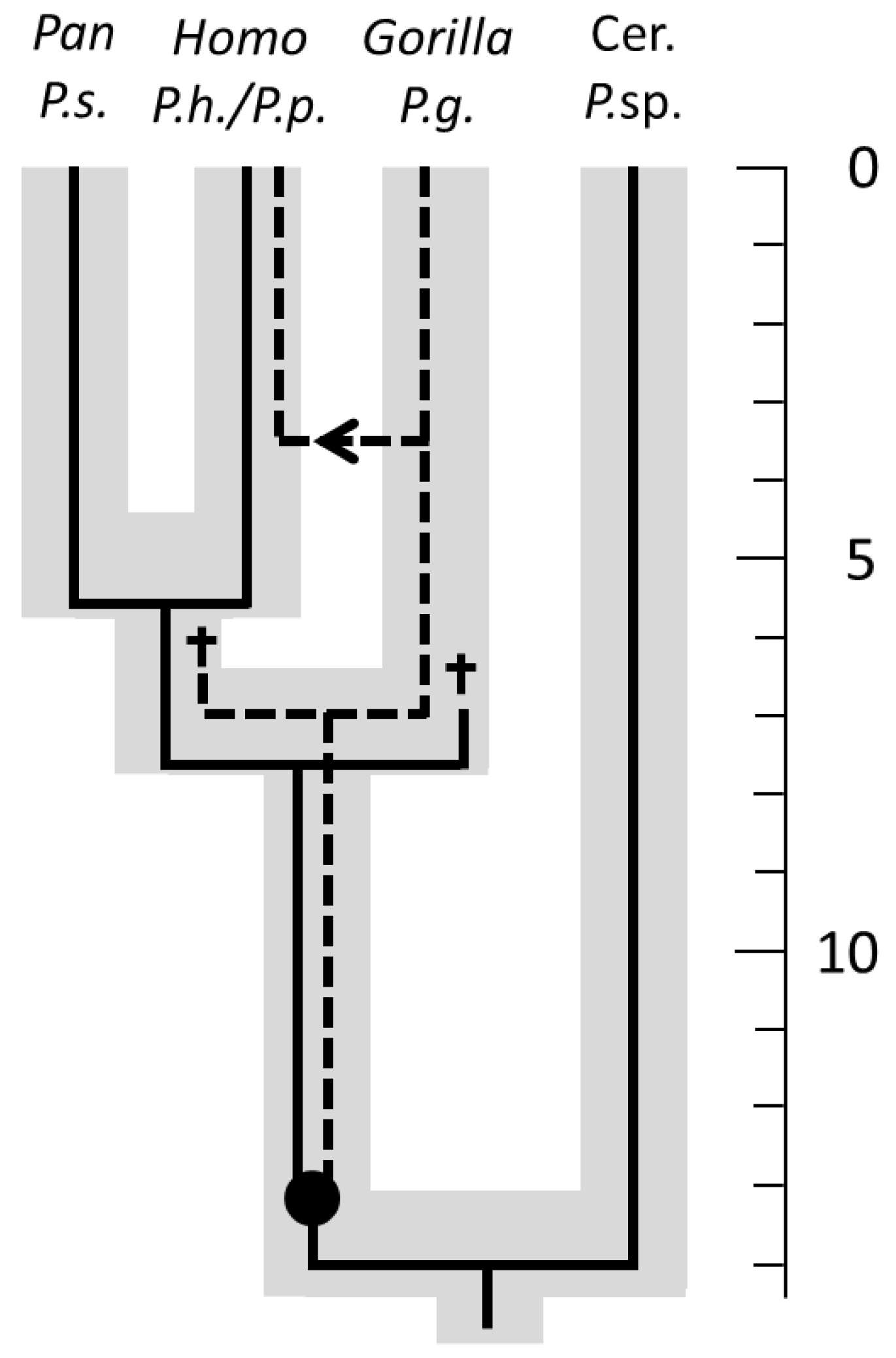
5. Co-Speciation and Phylogenetic Congruence
6. Phylogeny and Coevolution
7. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nunn, C.L.; Altizer, S. Infectious Diseases in Primates: Behavior, Ecology and Evolution; Oxford University Press: New York, NY, USA, 2006. [Google Scholar]
- Wolfe, N.D.; Escalante, A.A.; Karesh, W.B.; Kilbourn, A.; Spielman, A.; Lal, A.A. Wild primate populations in emerging infectious disease research: The missing link? Emerg. Infect. Dis. 1998, 4, 149–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, N.; Griffin, R.; Franz, M.; Omotayo, M.; Nunn, C.L. Phylogenetic host specificity and understanding parasite sharing in primates. Ecol. Lett. 2012, 15, 1370–1377. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, N.D.; Dunavan, C.P.; Diamond, J. Origins of major human infectious diseases. Nature 2007, 447, 279–283. [Google Scholar] [CrossRef]
- Nunn, C.L.; Altizer, S.; Sechrest, W.; Jones, K.E.; Barton, R.A.; Gittleman, J.L. Parasites and the evolutionary diversification of primate clades. Am. Nat. 2004, 164 (Suppl. 5), S90–S103. [Google Scholar] [CrossRef]
- Futuyma, D.J.; Kirkpatrick, M. Evolution, 4th ed.; Sinauer: New York, NY, USA, 2018. [Google Scholar]
- Costello, E.K.; Stagaman, K.; Dethlefsen, L.; Bohannan, B.J.M.; Relman, D.A. The application of ecological theory toward an understanding of the human microbiome. Science 2012, 336, 1255–1262. [Google Scholar] [CrossRef] [Green Version]
- Rees, T.; Bosch, T.C.G.; Douglas, A.E. How the microbiome challenges our concept of self. PLoS Biol. 2018, 16, e2005358. [Google Scholar] [CrossRef] [Green Version]
- Clayton, D.; Bush, S.E.; Johnson, K.P. Coevolution of Life on Hosts. Integrating Ecology and History; The University of Chicago Press: Chicago, IL, USA, 2016. [Google Scholar]
- Woolhouse, M.E.J.; Webster, J.P.; Domingo, E.; Charlesworth, B.; Levin, B.R. Biological and biomedical implications of the co-evolution of pathogens and their hosts. Nat. Genet. 2002, 32, 569–577. [Google Scholar] [CrossRef] [Green Version]
- Janzen, D.H. When is it coevolution? Evolution 1980, 34, 611–612. [Google Scholar] [CrossRef]
- Stuart, M.D.; Strier, K.B. Primates and parasites: A case for a multidisciplinary approach. Int. J. Primatol. 1995, 16, 577–593. [Google Scholar] [CrossRef]
- Page, R.D.M. Tangled trees: Phylogeny, cospeciation, and coevolution. Introduction. In Tangled Trees: Phylogeny, Cospeciation, and Coevolution; The University of Chicago Press: Chicago, IL, USA, 2003; pp. 1–21. [Google Scholar]
- Perlman, R.L. Evolution and Medicine; Oxford University Press: Oxford, UK, 2013. [Google Scholar]
- Ehrlich, P.R.; Raven, P.H. Butterflies and plants: A study in coevolution. Evolution 1964, 18, 586–608. [Google Scholar] [CrossRef]
- Darwin, C. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life; John Murray: London, UK, 1859. [Google Scholar]
- Darwin, C. On the Various Contrivances by Which British and Foreign Orchids Are Fertilised by Insects; John Murray: London, UK, 1862. [Google Scholar]
- Kritsky, G. Darwin’s Madagascan hawk moth prediction. Am. Entomol. 1991, 37, 206–210. [Google Scholar] [CrossRef] [Green Version]
- Weiblen, G.D.; Bush, G.L. Speciation in fig pollinators and parasites. Mol. Ecol. 2002, 11, 1573–1578. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.M.; Rasplus, J.Y. Mutualists with attitude: Coevolving fig wasps and figs. Trends Ecol. Evol. 2003, 18, 241–248. [Google Scholar] [CrossRef]
- Hossaert-McKey, M.; Gibernau, M.; Frey, J.E. Chemosensory attraction of fig wasps to substances produced by receptive figs. Entomol. Exp. Appl. 1994, 70, 185–191. [Google Scholar] [CrossRef]
- Ware, A.B.; Compton, S.G. Responses of fig wasps to host plant volatile cues. J. Chem. Ecol. 1994, 20, 785–802. [Google Scholar] [CrossRef]
- Stanley, D.A.; Msweli, S.M.; Johnson, S.D. Native honeybees as flower visitors and pollinators in wild plant communities in a biodiversity hotspot. Ecosphere 2020, 11, e02957. [Google Scholar] [CrossRef]
- Chapman, C.A. Primate seed dispersal: Coevolution and conservation implications. Evol. Anthropol. 1995, 4, 74–82. [Google Scholar] [CrossRef]
- Fox, L.R. Diffuse coevolution within complex communities. Ecology 1988, 69, 906–907. [Google Scholar] [CrossRef]
- Linhart, Y. Plant pollination and dispersal. In Ecology and the Environment; Monson, R.K., Ed.; Springer: New York, NY, USA, 2014; pp. 89–117. [Google Scholar]
- Clayton, J.B.; Gomez, A.; Amato, K.; Knights, D.; Travis, D.A.; Blekhman, R.; Knight, R.; Leigh, S.; Stumpf, R.; Wolf, T.; et al. The gut microbiome of nonhuman primates: Lessons in ecology and evolution. Am. J. Primatol. 2018, 80, e22867. [Google Scholar] [CrossRef]
- Abrams, P.A. The evolution of predator-prey interactions: Theory and evidence. Annu. Rev. Ecol. Syst. 2000, 31, 79–105. [Google Scholar] [CrossRef]
- Dawkins, R.; Krebs, J.R. Arms races between and within species. Proc. R. Soc. B 1979, 205, 489–511. [Google Scholar] [CrossRef]
- Zhao, H.; Luo, H.; Yan, H.; He, G.; Wang, L.; Liang, W. Fatal mobbing and attack of the common cuckoo by its warbler hosts. Ecol. Evol. 2022, 12, e9649. [Google Scholar] [CrossRef] [PubMed]
- Krausová, L.; Veselý, P.; Syrová, M.; Antonová, K.; Fišer, O.; Chlumská, V.; Pátková, M.; Pužej, Š.; Fuchs, R. Red-backed shrike (Lanius collurio) versus common cuckoo (Cuculus canorus): An example of ineffective cuckoo–hawk mimicry. Ecol. Evol. 2022, 12, e9664. [Google Scholar] [CrossRef] [PubMed]
- Van Valen, L. A new evolutionary law. Evol. Theory 1973, 1, 1–30. [Google Scholar]
- Martín-Navarro, I.; Brodie, J.P.; Romanowsky, A.J.; Ruiz-Lara, T.; van de Ven, G. Black-hole-regulated star formation in massive galaxies. Nature 2018, 553, 307–309. [Google Scholar] [CrossRef] [Green Version]
- Nunn, C.L.; Ezenwa, V.O.; Arnold, C.; Koenig, W.D. Mutualism or parasitism? Using a phylogenetic approach to characterize the oxpecker-ungulate relationship. Evolution 2011, 65, 1297–1304. [Google Scholar] [CrossRef]
- Arnqvist, G.; Rowe, L. Sexual conflict and arms races between the sexes: A morphological adaptation for control of mating in a female insect. Proc. R. Soc. B 1995, 261, 123–127. [Google Scholar] [CrossRef]
- Arnqvist, G.; Rowe, L. Sexual Conflict; Princeton University Press: Princeton, NJ, USA, 2005. [Google Scholar]
- Pérez-Barbería, F.J.; Shultz, S.; Dunbar, R.I.M. Evidence for coevolution of sociality and relative brain size in three orders of mammals. Evolution 2007, 61, 2811–2821. [Google Scholar] [CrossRef]
- Fox, K.C.R.; Muthukrishna, M.; Shultz, S. The social and cultural roots of whale and dolphin brains. Nat. Ecol. Evol. 2017, 1, 1699–1705. [Google Scholar] [CrossRef]
- Street, S.E.; Navarrete, A.F.; Reader, S.M.; Laland, K.N. Coevolution of cultural intelligence, extended life history, sociality, and brain size in primates. Proc. Natl. Acad. Sci. USA 2017, 114, 7908–7914. [Google Scholar] [CrossRef] [Green Version]
- Summers, W.C. Virus infection. In Encyclopedia of Microbiology, 3rd ed.; Schaechter, M., Ed.; Academic Press: Oxford, UK, 2009; pp. 546–552. [Google Scholar]
- Dybdahl, M.F.; Lively, C.M. Host-parasite coevolution: Evidence for rare advantage and time-lagged selection in a natural population. Evolution 1998, 52, 1057–1066. [Google Scholar] [CrossRef] [PubMed]
- Morgan, A.D.; Koskella, B. Coevolution of host and pathogen. In Genetics and Evolution of Infectious Disease; Tibayrenc, M., Ed.; Elsevier: London, UK, 2017; pp. 115–140. [Google Scholar] [CrossRef]
- Kleindorfer, S.; Custance, G.; Peters, K.J.; Sulloway, F. Introduced parasite changes host phenotype, mating signal and hybridization risk: Philornis downsi effects on Darwin’s finch song. Proc. R. Soc. B 2019, 286, 20190461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köndgen, S.; Kühl, H.; N’Goran, P.K.; Walsh, P.D.; Schenk, S.; Ernst, N.; Biek, R.; Formenty, P.; Mätz-Rensing, K.; Schweiger, B.; et al. Pandemic human viruses cause decline of endangered great apes. Curr. Biol. 2008, 18, 260–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patrono, L.V.; Samuni, L.; Corman, V.M.; Nourifar, L.; Röthemeier, C.; Wittig, R.M.; Drosten, C.; Calvignac-Spencer, S.; Leendertz, F. Human coronavirus OC43 outbreak in wild chimpanzees, Côte d’Ivoire, 2016. Emerg. Microbes Infect. 2018, 7, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, R.M.; May, R.M. Coevolution of hosts and parasites. Parasitology 1982, 85, 411–426. [Google Scholar] [CrossRef]
- Thompson, J.N. The Geographic Mosaic of Coevolution; The University of Chicago Press: Chicago, IL, USA, 2005. [Google Scholar]
- Paterson, S.; Vogwill, T.; Buckling, A.; Benmayor, R.; Spiers, A.J.; Thomson, N.R.; Quail, M.; Smith, F.; Walker, D.; Libberton, B.; et al. Antagonistic coevolution accelerates molecular evolution. Nature 2010, 464, 275–278. [Google Scholar] [CrossRef] [Green Version]
- Haldane, J.B.S. Disease and evolution. La Ric. Sci. 1949, 19, 1–11. [Google Scholar]
- Ebert, D. Host–parasite coevolution: Insights from the Daphnia–parasite model system. Curr. Opin. Microbiol. 2008, 11, 290–301. [Google Scholar] [CrossRef]
- Carius, H.J.; Little, T.J.; Ebert, D. Genetic variation in a host-parasite association: Potential for coevolution and frequency-dependent selection. Evolution 2001, 55, 1136–1145. [Google Scholar] [CrossRef]
- Siemens, D.H.; Roy, B.A. Tests for parasite-mediated frequency-dependent selection in natural populations of an asexual plant species. Evol. Ecol. 2005, 19, 321–338. [Google Scholar] [CrossRef]
- Koskella, B.; Lively, C.M. Evidence for negative frequency-dependent selection during experimental coevolution of a freshwater snail and a sterilizing trematode. Evolution 2009, 63, 2213–2221. [Google Scholar] [CrossRef] [PubMed]
- Okada, H.; Kuhn, C.; Feillet, H.; Bach, J.F. The ‘hygiene hypothesis’ for autoimmune and allergic diseases: An update. Clin. Exp. Immunol. 2010, 160, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Scudellari, M. News Feature: Cleaning up the hygiene hypothesis. Proc. Natl. Acad. Sci. USA 2017, 114, 1433–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thrall, P.H.; Antonovics, J. Polymorphism in sexual versus non-sexual disease transmission. Proc. R. Soc. B 1997, 264, 581–587. [Google Scholar] [CrossRef]
- Cooper, M.D.; Alder, M.N. The evolution of adaptive immune systems. Cell 2006, 124, 815–822. [Google Scholar] [CrossRef] [Green Version]
- Boehm, T.; Hirano, M.; Holland, S.J.; Das, S.; Schorpp, M.; Cooper, M.D. Evolution of alternative adaptive immune systems in vertebrates. Annu. Rev. Immunol. 2018, 36, 19–42. [Google Scholar] [CrossRef]
- Morran, L.T.; Schmidt, O.G.; Gelarden, I.A.; Parrish, R.C.; Lively, C.M. Running with the Red Queen: Host-parasite coevolution selects for biparental sex. Science 2011, 333, 216–218. [Google Scholar] [CrossRef] [Green Version]
- Ashby, B.; King, K.C. Diversity and the maintenance of sex by parasites. J. Evol. Biol. 2015, 28, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Bosch, T.C.; McFall-Ngai, M.J. Metaorganisms as the new frontier. Zoology 2011, 114, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Vorburger, C.; Perlman, S.J. The role of defensive symbionts in host–parasite coevolution. Biol. Rev. 2018, 93, 1747–1764. [Google Scholar] [CrossRef] [Green Version]
- Betts, A.; Gray, C.; Zelek, M.; MacLean, R.C.; King, K.C. High parasite diversity accelerates host adaptation and diversification. Science 2018, 360, 907–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunn, C.L.; Gillespie, T.R. Infectious disease and primate conservation. In An Introduction to Primate Conservation; Wich, S.A., Marshall, A.J., Eds.; Oxford University Press: Oxford, UK, 2016; pp. 151–173. [Google Scholar] [CrossRef] [Green Version]
- Huff, J.L.; Barry, P.A. B-virus (cercopithecine herpesvirus 1) infection in humans and macaques: Potential for zoonotic disease. Emerg. Infect. Dis. 2003, 9, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Keele, B.F.; Jones, J.H.; Terio, K.A.; Estes, J.D.; Rudicell, R.S.; Wilson, M.L.; Li, Y.; Learn, G.H.; Beasley, T.M.; Schumacher-Stankey, J.; et al. Increased mortality and AIDS-like immunopathology in wild chimpanzees infected with SIVcpz. Nature 2009, 460, 515–519. [Google Scholar] [CrossRef] [Green Version]
- Silvestri, G. Immunity in natural SIV infections. J. Intern. Med. 2009, 265, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Joas, S.; Parrish, E.H.; Gnanadurai, C.W.; Lump, E.; Stürzel, C.M.; Parrish, N.F.; Learn, G.H.; Sauermann, U.; Neumann, B.; Mätz-Rensing, K.; et al. Species-specific host factors rather than virus-intrinsic virulence determine primate lentiviral pathogenicity. Nat. Commun. 2018, 9, 1371. [Google Scholar] [CrossRef] [Green Version]
- Barreiro, L.B.; Quintana-Murci, L. From evolutionary genetics to human immunology: How selection shapes host defence genes. Nat. Rev. Genet. 2010, 11, 17. [Google Scholar] [CrossRef]
- Knell, R.J. Sexually transmitted disease and parasite-mediated sexual selection. Evolution 1999, 53, 957–961. [Google Scholar] [CrossRef]
- Tybur, J.M.; Gangestad, S.W. Mate preferences and infectious disease: Theoretical considerations and evidence in humans. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 3375–3388. [Google Scholar] [CrossRef] [Green Version]
- Gluckman, P.; Beedle, A.; Buklijas, T.; Low, F.; Hanson, M. Coevolution, infection, and immunity. In Principles of Evolutionary Medicine, 2nd ed.; Oxford University Press: Oxford, UK, 2016; pp. 237–258. [Google Scholar]
- Fraser, C.; Hollingsworth, T.D.; Chapman, R.; de Wolf, F.; Hanage, W.P. Variation in HIV-1 set-point viral load: Epidemiological analysis and an evolutionary hypothesis. Proc. Natl. Acad. Sci. USA 2007, 104, 17441–17446. [Google Scholar] [CrossRef] [Green Version]
- Fraser, C.; Lythgoe, K.; Leventhal, G.E.; Shirreff, G.; Hollingsworth, T.D.; Alizon, S.; Bonhoeffer, S. Virulence and pathogenesis of HIV-1 infection: An evolutionary perspective. Science 2014, 343, 1243727. [Google Scholar] [CrossRef] [Green Version]
- Paciência, F.M.D.; Rushmore, J.; Chuma, I.S.; Lipende, I.F.; Caillaud, D.; Knauf, S.; Zinner, D. Mating avoidance in female olive baboons (Papio anubis) infected by Treponema pallidum. Sci. Adv. 2019, 5, eaaw9724. [Google Scholar] [CrossRef] [Green Version]
- Paciência, F.M.D.; Chuma, I.S.; Lipende, I.F.; Knauf, S.; Zinner, D. Female post-copulatory behavior in a group of olive baboons (Papio anubis) infected by Treponema pallidum. PLoS ONE 2022, 17, e0261894. [Google Scholar] [CrossRef] [PubMed]
- Dieckmann, U. Adaptive dynamics of pathogen–host interactions. In Adaptive Dynamics of Infectious Diseases; Dieckmann, U., Metz, J.A.J., Sabelis, M.W., Sigmund, K., Eds.; Cambridge University Press: Cambridge, UK, 2002; pp. 39–59. [Google Scholar]
- Hawley, D.M.; Osnas, E.E.; Dobson, A.P.; Hochachka, W.M.; Ley, D.H.; Dhondt, A.A. Parallel patterns of increased virulence in a recently emerged wildlife pathogen. PLoS Biol. 2013, 11, e1001570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cressler, C.E.; McLeod, D.V.; Rozins, C.; Van Den Hoogen, J.; Day, T. The adaptive evolution of virulence: A review of theoretical predictions and empirical tests. Parasitology 2016, 143, 915–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telenti, A.; Hodcroft, E.B.; Robertson, D.L. The evolution and biology of SARS-CoV-2 variants. Cold Spring Harb. Perspect. Med. 2022, 12, a041390. [Google Scholar] [CrossRef]
- Fenner, F.J. The Florey Lecture, 1983—Biological control, as exemplified by smallpox eradication and myxomatosis. Proc. R. Soc. B 1983, 218, 259–285. [Google Scholar] [CrossRef]
- Buckling, A.; Maclean, R.C.; Brockhurst, M.A.; Colegrave, N. The Beagle in a bottle. Nature 2009, 457, 824–829. [Google Scholar] [CrossRef]
- Martínez-Aquino, A. Phylogenetic framework for coevolutionary studies: A compass for exploring jungles of tangled trees. Curr. Zool. 2016, 62, 393–403. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F. Phylogenetic approaches in coevolution and biogeography. Zool. Scr. 1998, 26, 313–322. [Google Scholar] [CrossRef]
- Hafner, M.S.; Nadler, S.A. Phylogenetic trees support the coevolution of parasites and their hosts. Nature 1988, 332, 258–259. [Google Scholar] [CrossRef]
- Hafner, M.S.; Nadler, S.A. Cospeciation in host-parasite assemblages: Comparative analysis of rates of evolution and timing of cospeciation events. Syst. Biol. 1990, 39, 192–204. [Google Scholar] [CrossRef]
- Whelan, S.; Liò, P.; Goldman, N. Molecular phylogenetics: State-of-the-art methods for looking into the past. Trends Genet. 2001, 17, 262–272. [Google Scholar] [CrossRef]
- Haeckel, E. Generelle Morphologie der Organismen: Allgemeine Grundzüge der organischen Formenwissenschaft, Mechanisch Begründet Durch die von Charles Darwin Reformirte Descendenztheorie. Band 2: Allgemeine Entwickelungsgeschichte der Organismen; Georg Reimer: Berlin, Germany, 1866. [Google Scholar]
- Edwards, A.W.F.; Cavalli-Sforza, L.L. Reconstruction of evolutionary trees. In Phenetic and Phylogenetic Classification; Heywood, V.H., McNeill, J., Eds.; Systematics Association: London, UK, 1964; pp. 67–76. [Google Scholar]
- Stearns, S.C.; Hoekstra, R.F. Evolution. An Introduction; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Raupach, M.J.; Amann, R.; Wheeler, Q.; Roos, C. The application of “omics”-technologies in the classification and identification of animals. Org. Divers. Evol. 2016, 16, 1–12. [Google Scholar] [CrossRef]
- Reed, D.L.; Light, J.E.; Allen, J.M.; Kirchman, J.J. Pair of lice lost or parasites regained: The evolutionary history of anthropoid primate lice. BMC Biol. 2007, 5, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svardal, H.; Jasinska, A.J.; Apetrei, C.; Coppola, G.; Huang, Y.; Schmitt, C.A.; Jacquelin, B.; Ramensky, V.; Müller-Trutwin, M.; Antonio, M.; et al. Ancient hybridization and strong adaptation to viruses across African vervet monkey populations. Nat. Genet. 2017, 49, 1705–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geoghegan, J.L.; Duchêne, S.; Holmes, E.C. Comparative analysis estimates the relative frequencies of co-divergence and cross-species transmission within viral families. PLoS Pathog. 2017, 13, e1006215. [Google Scholar] [CrossRef] [Green Version]
- Althoff, D.M.; Segraves, K.A.; Johnson, M.T.J. Testing for coevolutionary diversification: Linking pattern with process. Trends Ecol. Evol. 2014, 29, 82–89. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zinner, D.; Paciência, F.M.D.; Roos, C. Host–Parasite Coevolution in Primates. Life 2023, 13, 823. https://doi.org/10.3390/life13030823
Zinner D, Paciência FMD, Roos C. Host–Parasite Coevolution in Primates. Life. 2023; 13(3):823. https://doi.org/10.3390/life13030823
Chicago/Turabian StyleZinner, Dietmar, Filipa M. D. Paciência, and Christian Roos. 2023. "Host–Parasite Coevolution in Primates" Life 13, no. 3: 823. https://doi.org/10.3390/life13030823