
Responses to Drought Stress in Poplar: What Do We Know and What Can We Learn?
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
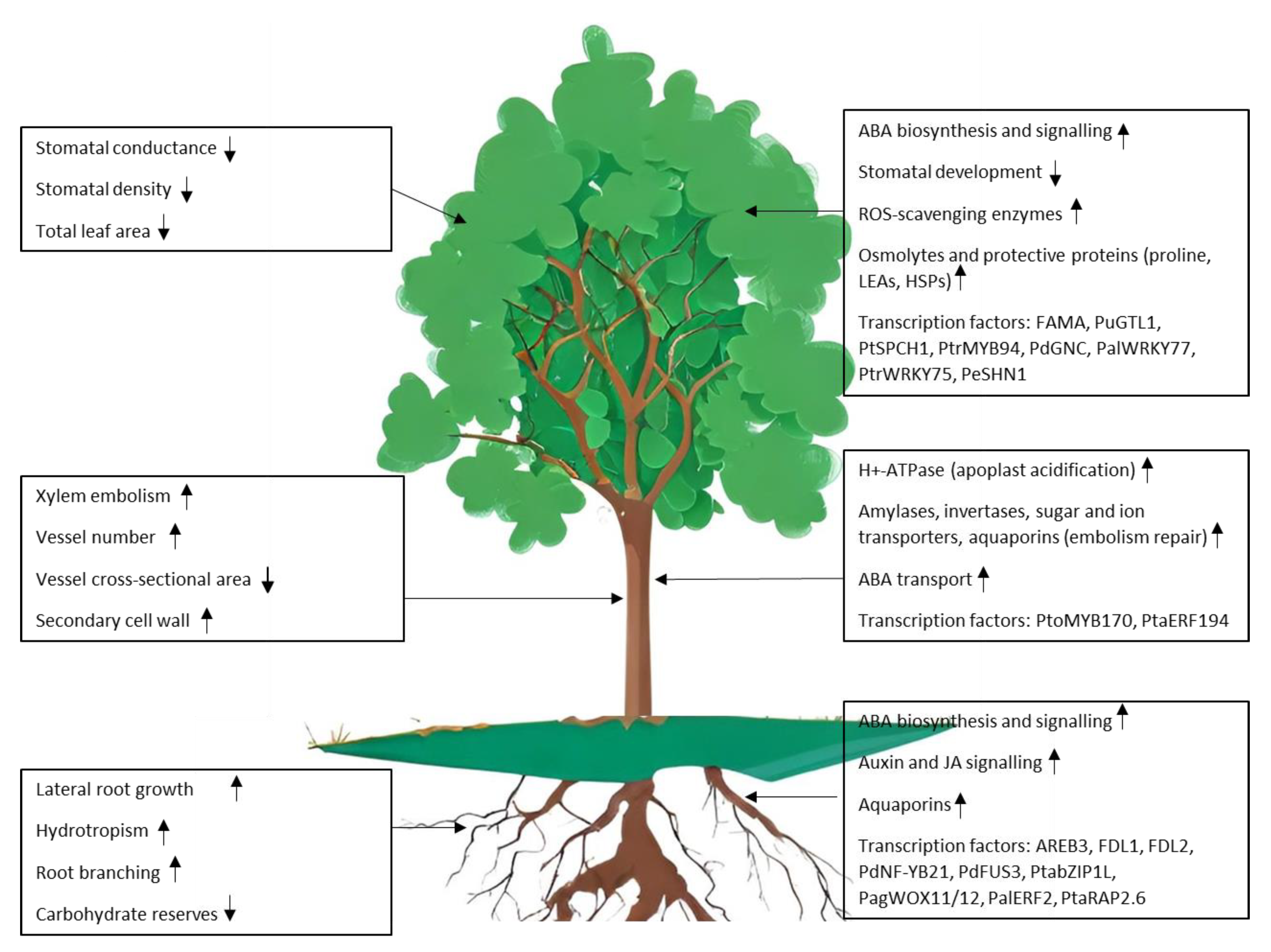
2. Drought Response in Poplar Is a Complex Trait
3. Mining Poplar Transcriptome for Seeking Pathways and Genes Involved in Drought Response
4. Identification of Genes and Pathways Modulating Stomatal Development and Function
5. Control of Non-Stomatal Water Loss by Cuticular Waxes
6. Enhancing Water Uptake and Belowground Traits
7. Epigenetic Regulation of Drought Responses
8. Embolism Formation and Repair Mechanisms
9. Drought and Wood Quality
10. Water Stress Versus Biotic Adversities
- -
- among biochemical changes, a decrease in starch level and a concomitant increase in glucose and fructose levels in various bark tissues; e.g., increased glucose content can enhance attacks by Armillaria mellea (Vahl) P. Kumm., a polyphagous root rot agent [191]; similarly, increased concentrations of amino acids can stimulate hyphal growth of Entoleuca mammata (Wahlenb.) J.D. Rogers & Y.M. Ju, the Hypoxylon canker agent in quaking aspen [192];
- -
- among metabolic changes, the synthesis of antifungal compounds and phytoalexins can be inhibited by water stress, e.g., catechol, salicortin and salicin in quaking aspen versus E. mammata [193];
- -
- among anatomopathological features, water stress may delay the formation of necrophylactic periderm, a barrier tissue mitigating colonization by pathogens;
- -
- among biophysical changes, xylem embolisms associated with low water potential may provide preferential routes for the spread of internal pathogens [194].
11. Phenotyping Drought Tolerance Traits
12. Conclusion and Perspectives

{kind=link}
| Gene | Species | Encoded Product | Note | Reference |
|---|---|---|---|---|
| PtrWRKY75 | P. tremula | WRKY transcription factor | Overexpressing plants display increased salicylic acid and ROS accumulation, higher photosynthetic rate, and increased growth under drought conditions. | [38] |
| PtHMGR | P. × euramericana | Hydroxy-3-methylglutaryl coenzyme A reductase | Overexpression improves drought and salinity tolerance by promoting root development and the expression of genes coding for ROS-scavenging enzymes. | [48] |
| PdEPF1 | P. deltoides | Signaling peptide | Overexpression induces lower stomatal density and transpiration, increased WUE, increased drought tolerance. | [55] |
| PdERECTA | P. nigra × (P. deltoides × P. nigra) | Receptor-like kinase LRR-RLK | Overexpressing plants have reduced stomatal density and display increased drought tolerance and WUE under drought conditions. | [57,58] |
| PdEPFL6 | P. nigra × (P. deltoides × P. nigra) | Signaling peptide | Overexpression induces lower stomatal density and transpiration, increased WUE, increased drought tolerance. | [59] |
| STOMAGEN | P. alba × P. glandulosa | Signaling peptide | Overexpressing plants display increased stomatal density, higher net photosynthetic rate, and higher vegetative growth. | [60] |
| PtaGTL1 | P. tremula × P. alba | Ca2+/Calmodulin-binding transcription factor | Overexpressing plants display reduced stomatal density and transpiration, increased WUE, growth retardation. | [61] |
| Pu-miR172d | P. ussuriensis | MicroRNA | Overexpressing plants display reduced expression of GTL1, reduced stomatal density, increased WUE, and growth retardation. | [62] |
| PePYL6; PePYL9 | P. euphratica | PYR/PYL/RCAR ABA receptors | Overexpression in Arabidopsis confers increased ABA sensitivity, increased WUE, and drought tolerance. | [69] |
| PtPYRL1 | P. trichocarpa | PYR/PYL/RCAR ABA receptor | Overexpression in transgenic poplars improves ABA sensitivity and drought-stress tolerance. | [74] |
| PtPYRL5 | P. trichocarpa | PYR/PYL/RCAR ABA receptor | Overexpression in transgenic poplars improves ABA sensitivity and drought-stress tolerance. | [74] |
| RCAR1/PYL9 | P. tremula × P. tremuloides | PYR/PYL/RCAR ABA receptor | Overexpressing plants display increased biomass accumulation under non-limiting water condition. | [75] |
| AREB3 | P. tremula × P. tremuloides | ABA-Responsive Element Binding protein | Overexpressing plants display increased drought tolerance, biomass re-allocation favouring roots in drought conditions, severely reduced productivity under well-watered conditions. | [75] |
| FDL1, FDL2 | P. tremula × P. tremuloides | bZIP transcription factors | Overexpressing poplar lines show reduced root growth and increased drought sensitivity; RNAi downregulated lines show higher biomass allocation to roots under drought conditions. | [75] |
| PtrMYB94 | P. trichocarpa | MYB transcription factor | Overexpression increases the expression of ABA- and drought-responsive genes and improves drought tolerance. | [77] |
| PdGNC | P. nigra × (P. deltoides × P. nigra) | GATA transcription factor | Overexpression in poplar plants results in increased hexokinase expression in guard cells, production of NO and H2O2, stomatal closure and increased drought resistance. CRISPR/Cas9-mediated mutagenesis results in increased stomatal aperture. | [78] |
| PtXERICO | P. trichocarpa | RING-H2 zinc finger E3 ubiquitin ligase | Overexpression in Arabidopsis confers ABA hypersensitivity and drought tolerance. | [79] |
| PalPUB79 | P. alba | E3 ubiquitin ligase | Overexpressing plants display increased drought tolerance and upregulated ABA signaling due to ubiquitin-mediated degradation of PalWRKY77, a negative regulator of ABA signal transduction; RNAi lines display the opposite phenotype. | [80] |
| PeSHN1 | P. alba × P. glandulosa | AP2/ERF transcription factor | Overexpressing plants display increased cuticular wax accumulation, decreased transpiration, higher photosynthetic activity and WUE in drought conditions. | [84] |
| PdNF-YB21 | P. deltoides | Nuclear factor-Y transcription factor | Overexpression results in increased ABA concentration, promotion of auxin transport to root tips, increased root growth and drought tolerance; CRISPR/Cas9-mediated mutants display opposite phenotype. | [97] |
| PtaSUT4 | P. tremula × P. alba | Sucrose transporter | Under mild drought conditions RNAi downregulated lines display reduced shoot growth compared to wild type and altered gene expression patterns in root tips and stem xylem. | [99] |
| PtabZIP1 | P. tremula × P. alba | bZIP transcription factor | Overexpression in poplar plants results in increased lateral root development under drought and osmotic stress conditions. | [100] |
| PtaJAZ3 | P. tremula × P. alba | Jasmonate ZIM-domain protein | Overexpressing plants display increased lateral root proliferation under drought stress. | [101] |
| PtaRAP2.6 | P. tremula × P. alba | Related to Apetala 2.6 transcription factor | Overexpressing plants display increased lateral root proliferation under drought stress. | [101] |
| PagWOX11/12a | P. alba × P. glandulosa | WUSCHEL-related homeobox (WOX) transcription factor | Overexpression results in increased root biomass and drought tolerance; downregulation results in the opposite phenotype. | [104] |
| PtDDM1 | P. tremula × P. alba | SWI/SNF chromatin remodelling factor | Silenced lines display DNA hypomethylation, increased drought tolerance, increased accumulation of salicylic acid and reduced accumulation of cytokinins in the shoot apex. | [127] |
| PtaPIP1.1 | P. alba × P. tremula | Acquaporin | Downregulated poplars show increased vulnerability to embolism and reduced stomatal control of transpiration. | [169] |
| PtoMYB170 | P. tomentosa | MYB transcription factor | Overexpression results in stronger lignification, thickened secondary wall in xylem and enhanced stomatal closure; downregulation weaken lignin deposition. | [173] |
| PtaERF194 | P. tremula × P. alba | AP2/ERF transcription factor | Overexpression results in increased number of stem xylem vessels and growth inhibition. Under drought conditions overexpressing plants display lower stomatal conductance and transcriptional activation of genes involved in drought resistance. | [174] |
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2022: Impacts, Adaptation and Vulnerability; Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Pörtner, H.-O., Roberts, D.C., Tignor, M., Poloczanska, E.S., Mintenbeck, K., Alegría, A., Craig, M., Langsdorf, S., Löschke, S., Möller, V., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022; p. 3056. [Google Scholar] [CrossRef]
- Menezes-Silva, P.E.; Loram-Lourenço, L.; Alves, R.D.F.B.; Sousa, L.F.; da Almeida, S.E.; Farnese, F.S. Different ways to die in a changing world: Consequences of climate change for tree species performance and survival through an ecophysiological perspective. Ecol. Evol. 2019, 9, 11979–11999. [Google Scholar] [CrossRef] [Green Version]
- Brodribb, T.J.; Powers, J.; Cochard, H.; Choat, B. Hanging by a thread? Forests and drought. Science 2020, 368, 261–266. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global Convergence in the Vulnerability of Forests to Drought. Nature 2012, 491, 752–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. Global Forest Resources Assessment 2020: Main Report; FAO: Rome, Itlay, 2020. [Google Scholar] [CrossRef]
- Harfouche, A.; Meilan, R.; Altman, A. Molecular and physiological responses to abiotic stress in forest trees and their relevance to tree improvement. Tree Physiol. 2014, 34, 1181–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polle, A.; Chen, S.L.; Eckert, C.; Harfouche, A. Engineering drought resistance in forest trees. Front. Plant Sci. 2019, 9, 1875. [Google Scholar] [CrossRef] [Green Version]
- Alberto, F.J.; Aitken, S.N.; Alía, R.; González-Martínez, S.C.; Hänninen, H.; Kremer, A.; Lefèvre, F.; Lenormand, T.; Yeaman, S.; Whetten, R.; et al. Potential for evolutionary responses to climate change—Evidence from tree populations. Glob. Chang. Biol. 2013, 19, 1645–1661. [Google Scholar] [CrossRef] [Green Version]
- Marchi, M.; Bergante, S.; Ray, D.; Barbetti, R.; Facciotto, G.; Chiarabaglio, P.M.; Hynynen, J.; Nervo, G. Universal Reaction Norms for the Sustainable Cultivation of Hybrid Poplar Clones under Climate Change in Italy. IForest 2022, 15, 47. [Google Scholar] [CrossRef]
- Bradshaw, H.D.; Ceulemans, R.; Davis, J.; Stettler, R. Emerging Model Systems in Plant Biology: Poplar (Populus) as a Model Forest Tree. J. Plant Growth Regul. 2000, 19, 306–313. [Google Scholar] [CrossRef]
- Tuskan, G.A.; DiFazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, M.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanton, B.J.; Neale, D.B.; Li, S. Populus Breeding: From the Classical to the Genomic Approach. In Genetics and Genomics of Populus; Jansson, S., Bhalerao, R., Groover, A., Eds.; Springer: New York, NY, USA, 2010; Volume 8, pp. 309–348. [Google Scholar] [CrossRef]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigues, M.L.; Ricardo, C.P.P.; Osório, M.L.; Carvalho, I.; Faria, T.; Pinheiro, C. How plants cope with water stress in the field. Photosynthesis and growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef] [Green Version]
- Cruz De Carvalho, M.H. Drought stress and reactive oxygen species: Production, scavenging and signaling. Plant Sign. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Attia, Z.; Domec, J.C.; Oren, R.; Way, D.A.; Moshelion, M. Growth and physiological responses of isohydric and anisohydric poplars to drought. J. Exp. Bot. 2015, 66, 4373–4381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, E.J.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Richards, C.L.; Valladares, F.; et al. Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef]
- Viger, M.; Smith, H.K.; Cohen, D.; Dewoody, J.; Trewin, H.; Steenackers, M.; Bastien, C.; Taylor, G. Adaptive mechanisms and genomic plasticity for drought tolerance identified in European black poplar (Populus nigra L.). Tree Physiol. 2016, 36, 909–928. [Google Scholar] [CrossRef] [Green Version]
- Garavillon-Tournayre, M.; Gousset-Dupont, A.; Gautier, F.; Benoit, P.; Conchon, P.; Souchal, R.; Lopez, D.; Petel, G.; Venisse, J.S.; Bastien, C.; et al. Integrated drought responses of black poplar: How important is phenotypic plasticity? Physiol. Plant. 2018, 163, 30–44. [Google Scholar] [CrossRef]
- Traversari, S.; Neri, A.; Traversi, M.L.; Giovannelli, A.; Francini, A.; Sebastiani, L. Daily Osmotic Adjustments in Stem May Be Good Predictors of Water Stress Intensity in Poplar. Plant Physiol. Biochem. 2020, 146, 13–22. [Google Scholar] [CrossRef]
- Galović, V.; Kebert, M.; Popović, B.M.; Kovačević, B.; Vasić, V.; Joseph, M.P.; Orlović, S.; Szabados, L. Biochemical and Gene Expression Analyses in Different Poplar Clones: The Selection Tools for Afforestation of Halomorphic Environments. Forests 2021, 12, 636. [Google Scholar] [CrossRef]
- Jouve, L.; Hoffmann, L.; Hausman, J.F. Polyamine, carbohydrate, and proline content changes during salt stress exposure of aspen (Populus tremula L.): Involvement of oxidation and osmoregulation metabolism. Plant Biol. 2004, 6, 74–80. [Google Scholar] [CrossRef]
- Barchet, G.L.H.; Dauwe, R.; Guy, R.D.; Schroeder, W.R.; Soolanayakanahally, R.Y.; Campbell, M.M.; Mansfield, S.D. Investigating the drought-stress response of hybrid poplar genotypes by metabolite profiling. Tree Physiol. 2014, 34, 1203–1219. [Google Scholar] [CrossRef] [Green Version]
- Monclus, R.; Dreyer, E.; Villar, M.; Delmotte, F.M.; Delay, D.; Petit, J.M.; Barbaroux, C.; Le Thiec, D.; Bréchet, C.; Brignolas, F. Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoides × Populus nigra. New Phytol. 2006, 169, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Monclus, R.; Villar, M.; Barbaroux, C.; Bastien, C.; Fichot, R.; Delmotte, F.M.; Delay, D.; Petit, J.M.; Brchet, C.; Dreyer, E.; et al. Productivity, water-use efficiency and tolerance to moderate water deficit correlate in 33 poplar genotypes from a Populus deltoides × Populus trichocarpa F1 progeny. Tree Physiol. 2009, 29, 1329–1339. [Google Scholar] [CrossRef] [Green Version]
- Larchevêque, M.; Maurel, M.; Desrochers, A.; Larocque, G.R. How does drought tolerance compare between two improved hybrids of balsam poplar and an unimproved native species? Tree Physiol. 2011, 31, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Sun, P.; Cheng, X.; Zhang, L.; Wang, L.; Hu, J. QTL Mapping of drought-related traits in the hybrids of Populus deltoides ‘Danhong’× Populus simonii ‘Tongliao1’. BMC Plant Biol. 2022, 22, 238. [Google Scholar] [CrossRef]
- Street, N.R.; Skogström, O.; Sjödin, A.; Tucker, J.; Rodríguez-Acosta, M.; Nilsson, P.; Jansson, S.; Taylor, G. The genetics and genomics of the drought response in Populus. Plant J. 2006, 48, 321–341. [Google Scholar] [CrossRef] [PubMed]
- Tschaplinski, T.J.; Tuskan, G.A.; Sewell, M.M.; Gebre, G.M.; Todd, D.E.; Pendley, C.D. Phenotypic variation and quantitative trait locus identification for osmotic potential in an interspecific hybrid inbred F2 poplar pedigree grown in contrasting environments. Tree Physiol. 2006, 26, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Biselli, C.; Vietto, L.; Rosso, L.; Cattivelli, L.; Nervo, G.; Fricano, A. Advanced breeding for biotic stress resistance in poplar. Plants 2022, 11, 2032. [Google Scholar] [CrossRef]
- Cohen, D.; Bogeat-Triboulot, M.B.; Tisserant, E.; Balzergue, S.; Martin-Magniette, M.L.; Lelandais, G.; Ningre, N.; Renou, J.P.; Tamby, J.P.; Le Thiec, D.; et al. Comparative transcriptomics of drought responses in Populus: A meta-analysis of genome-wide expression profiling in mature leaves and root apices across two genotypes. BMC Genom. 2010, 11, 630. [Google Scholar] [CrossRef] [Green Version]
- Yan, D.H.; Fenning, T.; Tang, S.; Xia, X.; Yin, W. Genome-wide transcriptional response of Populus euphratica to long-term drought stress. Plant Sci. 2012, 195, 24–35. [Google Scholar] [CrossRef]
- Takahashi, F.; Kuromori, T.; Urano, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Drought Stress Responses and Resistance in Plants: From Cellular Responses to Long-Distance Intercellular Communication. Front. Plant Sci. 2020, 11, 556972. [Google Scholar] [CrossRef] [PubMed]
- Kopecky, D.; Matušíková, I.; Sziderics, A.H.; Trognitz, F.; Spieß, N.; Stierschneider, M.; Fluch, S. In silico search for drought-responsive genes in plants on the basis of scientific data: Case study on poplar roots. Acta Physiol. Plant. 2013, 35, 1955–1966. [Google Scholar] [CrossRef]
- Secchi, F.; Gilbert, M.E.; Zwieniecki, M.A. Transcriptome response to embolism formation in stems of Populus trichocarpa provides insight into signaling and the biology of refilling. Plant Physiol. 2011, 157, 1419–1429. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.; Liang, H.; Yan, D.; Zhao, Y.; Han, X.; Carlson, J.E.; Xia, X.; Yin, W. Populus euphratica: The transcriptomic response to drought stress. Plant Mol. Biol. 2013, 83, 539–557. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Y.; Zhang, D.; Tang, X.; Li, Z.; Shen, C.; Han, X.; Deng, W.; Yin, W.; Xia, X. PtrWRKY75 overexpression reduces stomatal aperture and improves drought tolerance by salicylic acid-induced reactive oxygen species accumulation in poplar. Environ. Exp. Bot. 2020, 176, 104117. [Google Scholar] [CrossRef]
- Yıldırım, K.; Kaya, Z. Gene regulation network behind drought escape, avoidance and tolerance strategies in black poplar (Populus nigra L.). Plant Physiol. Biochem. 2017, 115, 183–199. [Google Scholar] [CrossRef] [PubMed]
- Hamanishi, E.T.; Barchet, G.L.H.; Dauwe, R.; Mansfield, S.D.; Campbell, M.M. Poplar trees reconfigure the transcriptome and metabolome in response to drought in a genotype- and time-of-day-dependent manner. BMC Genom. 2015, 16, 329. [Google Scholar] [CrossRef]
- Chen, J.; Song, Y.; Zhang, H.; Zhang, D. Genome-wide analysis of gene expression in response to drought stress in Populus simonii. Plant Mol. Biol. Rep. 2013, 31, 946–962. [Google Scholar] [CrossRef]
- Urban, J.; Ingwers, M.W.; McGuire, M.A.; Teskey, R.O. Increase in leaf temperature opens stomata and decouples net photosynthesis from stomatal conductance in Pinus taeda and Populus deltoides x P. nigra. J. Exp. Bot. 2017, 68, 1757–1767. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.K.; Park, E.J.; Choi, Y.I.; Bae, E.K.; Kim, J.H.; Park, S.Y.; Kang, K.S.; Lee, H. Response to drought and salt stress in leaves of poplar (Populus alba × P. glandulosa): Expression profiling by oligonucleotide microarray analysis. Plant Physiol. Biochem. 2014, 84, 158–168. [Google Scholar] [CrossRef]
- Jia, J.; Zhou, J.; Shi, W.; Cao, X.; Luo, J.; Polle, A.; Luo, Z.B. Comparative transcriptomic analysis reveals the roles of overlapping heat-/drought-responsive genes in poplars exposed to high temperature and drought. Sci. Rep. 2017, 7, 43215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cossu, R.M.; Giordani, T.; Cavallini, A.; Natali, L. High-throughput analysis of transcriptome variation during water deficit in a poplar hybrid: A general overview. Tree Genet. Genomes 2014, 10, 53–66. [Google Scholar] [CrossRef]
- Barghini, E.; Cossu, R.M.; Cavallini, A.; Giordani, T. Transcriptome analysis of response to drought in poplar interspecific hybrids. Genom. Data 2015, 3, 143–145. [Google Scholar] [CrossRef]
- Zhang, J.; Gao, F.; Jia, H.; Hu, J.; Feng, Z. Molecular response of poplar to single and combined ozone and drought. Sci. Total Environ. 2019, 655, 1364–1375. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Movahedi, A.; Xu, C.; Sun, W.; Li, L.; Wang, P.; Li, D.; Zhuge, Q. Overexpression of PtHMGR enhances drought and salt tolerance of poplar. Ann. Bot. 2020, 125, 785–803. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Qin, Y.; Duan, H.; Yin, W.; Xia, X. Genome-wide characterization of new and drought stress responsive microRNAs in Populus euphratica. J. Exp. Bot. 2011, 62, 3765–3779. [Google Scholar] [CrossRef] [Green Version]
- Shuai, P.; Liang, D.; Zhang, Z.; Yin, W.; Xia, X. Identification of drought-responsive and novel Populus trichocarpa microRNAs by high-throughput sequencing and their targets using degradome analysis. BMC Genom. 2013, 14, 233. [Google Scholar] [CrossRef]
- Han, S.K.; Kwak, J.M.; Qi, X. Stomatal Lineage Control by Developmental Program and Environmental Cues. Front. Plant. Sci. 2021, 12, 751852. [Google Scholar] [CrossRef]
- Hamanishi, E.T.; Thomas, B.R.; Campbell, M.M. Drought induces alterations in the stomatal development program in Populus. J. Exp. Bot. 2012, 63, 4959–4971. [Google Scholar] [CrossRef] [Green Version]
- Miyazawa, S.I.; Livingston, N.J.; Turpin, D.H. Stomatal development in new leaves is related to the stomatal conductance of mature leaves in poplar (Populus trichocarpa × P. deltoides). J. Exp. Bot. 2006, 57, 373–380. [Google Scholar] [CrossRef]
- Ehonen, S.; Hölttä, T.; Kangasjärvi, J. Systemic signalling in the regulation of stomatal conductance. Plant Physiol. 2020, 182, 1829–1832. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Liu, S.; Dong, Y.; Zhao, Y.; Geng, A.; Xia, X.; Yin, W. PdEPF1 regulates water-use efficiency and drought tolerance by modulating stomatal density in poplar. Plant Biotechnol. J. 2016, 14, 849–860. [Google Scholar] [CrossRef] [Green Version]
- Hatfield, J.L.; Dold, C. Water-Use Efficiency: Advances and Challenges in a Changing Climate. Front. Plant. Sci. 2019, 10, 103. [Google Scholar] [CrossRef] [Green Version]
- Xing, H.T.; Guo, P.; Xia, X.L.; Yin, W.L. PdERECTA, a leucine-rich repeat receptor-like kinase of poplar, confers enhanced water use efficiency in Arabidopsis. Planta 2011, 234, 229–241. [Google Scholar] [CrossRef]
- Li, H.; Yang, Y.; Wang, H.; Liu, S.; Jia, F.; Su, Y.; Li, S.; He, F.; Feng, C.; Niu, M.; et al. The receptor-like kinase ERECTA confers improved water use efficiency and drought tolerance to poplar via modulating stomatal density. Int. J. Mol. Sci. 2021, 22, 7245. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Z.; Han, S.; Li, Z.; Huang, M.; Niu, M.-X.; Yu, X.; Liu, C.; Wang, H.-L.; Yin, W.; Xia, X. PdEPFL6 reduces stomatal density to improve drought tolerance in poplar. Ind. Crops Prod. 2022, 182, 114873. [Google Scholar] [CrossRef]
- Xia, Y.; Du, K.; Ling, A.; Wu, W.; Li, J.; Kang, X. Overexpression of PagSTOMAGEN, a positive regulator of stomatal density, promotes vegetative growth in poplar. Int. J. Mol. Sci. 2022, 23, 10165. [Google Scholar] [CrossRef]
- Weng, H.; Yoo, C.Y.; Gosney, M.J.; Hasegawa, P.M.; Mickelbart, M.V. Poplar GTL1 is a Ca2+/calmodulin-binding transcription factor that functions in plant water use efficiency and drought tolerance. PLoS ONE 2012, 7, e32925. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wang, Z.; Yu, S.; Li, W.; Zhang, M.; Yang, J.; Li, D.; Yang, J.; Li, C. Pu-MiR172d regulates stomatal density and water-use efficiency via targeting PuGTL1 in poplar. J. Exp. Bot. 2021, 72, 1370–1383. [Google Scholar] [CrossRef]
- Hetherington, A.M.; Woodward, F.I. The role of stomata in sensing and driving environmental change. Nature 2003, 424, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Drake, P.L.; Froend, R.H.; Franks, P.J. Smaller, faster stomata: Scaling of stomatal size, rate of response, and stomatal conductance. J. Exp. Bot. 2013, 64, 495–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearce, D.W.; Millard, S.; Bray, D.F.; Rood, S.B. Stomatal characteristics of riparian poplar species in a semi-arid environment. Tree Physiol. 2006, 26, 211–218. [Google Scholar] [CrossRef]
- Himes, A.; Emerson, P.; McClung, R.; Renninger, H.; Rosenstiel, T.; Stanton, B. Leaf traits indicative of drought resistance in hybrid poplar. Agric. Water Manag. 2021, 246, 106676. [Google Scholar] [CrossRef]
- McKown, A.D.; Guy, R.D.; Quamme, L.; Klápště, J.; La Mantia, J.; Constabel, C.P.; El-Kassaby, Y.A.; Hamelin, R.C.; Zifkin, M.; Azam, M.S. Association genetics, geography and ecophysiology link stomatal patterning in Populus trichocarpa with carbon gain and disease resistance trade-offs. Mol. Ecol. 2014, 23, 5771–5790. [Google Scholar] [CrossRef]
- McKown, A.D.; Klápště, J.; Guy, R.D.; Corea, O.R.A.; Fritsche, S.; Ehlting, J.; El-Kassaby, Y.A.; Mansfield, S.D. A role for SPEECHLESS in the integration of leaf stomatal patterning with the growth vs disease trade-off in poplar. New Phytol. 2019, 223, 1888–1903. [Google Scholar] [CrossRef]
- Chhetri, H.B.; Macaya-Sanz, D.; Kainer, D.; Biswal, A.K.; Evans, L.M.; Chen, J.G.; Collins, C.; Hunt, K.; Mohanty, S.S.; Rosenstiel, T.; et al. Multitrait genome-wide association analysis of Populus trichocarpa identifies key polymorphisms controlling morphological and physiological traits. New Phytol. 2019, 223, 293–309. [Google Scholar] [CrossRef] [PubMed]
- Papacek, M.; Christmann, A.; Grill, E. Interaction network of ABA receptors in grey poplar. Plant J. 2017, 92, 199–210. [Google Scholar] [CrossRef]
- Yu, J.; Li, H.; Peng, Y.; Yang, L.; Zhao, F.; Luan, S.; Lan, W. A survey of the pyrabactin resistance-like abscisic acid receptor gene family in poplar. Plant Signal. Behav. 2017, 12, e1356966. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Yang, L.; Liu, X.; Tang, R.; Wang, Y.; Ge, H.; Wu, M.; Zhang, J.; Zhao, F.; Luan, S.; et al. Overexpression of poplar pyrabactin resistance-like abscisic acid receptors promotes abscisic acid sensitivity and drought resistance in transgenic Arabidopsis. PLoS ONE 2016, 11, e0168040. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Tian, Q.; Zhang, Y.; Niu, M.; Yu, X.; Lian, C.; Liu, C.; Wang, H.L.; Yin, W.; Xia, X. Increased abscisic acid sensitivity and drought tolerance of Arabidopsis by overexpression of poplar abscisic acid receptors. Plant Cell Tissue Organ Cult. 2022, 148, 231–245. [Google Scholar] [CrossRef]
- Yu, J.; Ge, H.; Wang, X.; Tang, R.; Wang, Y.; Zhao, F.; Lan, W.; Luan, S.; Yang, L. Overexpression of pyrabactin resistance-like abscisic acid receptors enhances drought, osmotic, and cold tolerance in transgenic poplars. Front. Plant Sci. 2017, 8, 1752. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Wildhagen, H.; Tylewicz, S.; Miskolczi, P.C.; Bhalerao, R.P.; Polle, A. Abscisic acid signalling mediates biomass trade-off and allocation in poplar. New Phytol. 2019, 223, 1192–1203. [Google Scholar] [CrossRef] [PubMed]
- Arend, M.; Schnitzler, J.P.; Ehlting, B.; Hänsch, R.; Lange, T.; Rennenberg, H.; Himmelbach, A.; Grill, E.; Fromm, J. Expression of the Arabidopsis mutant Abi1 gene alters abscisic acid sensitivity, stomatal development, and growth morphology in grey poplars. Plant Physiol. 2009, 151, 2110–2119. [Google Scholar] [CrossRef] [Green Version]
- Fang, Q.; Wang, X.; Wang, H.; Tang, X.; Liu, C.; Yin, H.; Ye, S.; Jiang, Y.; Duan, Y.; Luo, K. The poplar R2R3 MYB transcription factor PtrMYB94 coordinates with abscisic acid signalling to improve drought tolerance in plants. Tree Physiol. 2019, 40, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Zhang, Y.; Li, Q.; Liu, S.; He, F.; An, Y.; Zhou, Y.; Liu, C.; Yin, W.; Xia, X. PdGNC confers drought tolerance by mediating stomatal closure resulting from NO and H2O2 production via the direct regulation of PdHXK1 expression in Populus. New Phytol. 2021, 230, 1868–1882. [Google Scholar] [CrossRef]
- Kim, M.H.; Cho, J.S.; Park, E.J.; Lee, H.; Choi, Y.I.; Bae, E.K.; Han, K.H.; Ko, J.H. Overexpression of a poplar RING-H2 zinc Finger, Ptxerico, confers enhanced drought tolerance via reduced water loss and ion leakage in Populus. Int. J. Mol. Sci. 2020, 21, 9454. [Google Scholar] [CrossRef]
- Tong, S.; Chen, N.; Wang, D.; Ai, F.; Liu, B.; Ren, L.; Chen, Y.; Zhang, J.; Lou, S.; Liu, H.; et al. The U-box E3 ubiquitin ligase PalPUB79 positively regulates ABA-dependent drought tolerance via ubiquitination of PalWRKY77 in Populus. Plant Biotechnol. J. 2021, 19, 2561–2575. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, M.; Keyl, A.; Feussner, I. Wax biosynthesis in response to danger: Its regulation upon abiotic and biotic stress. New Phytol. 2020, 227, 698–713. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.B.; Suh, M.C. Regulatory mechanisms underlying cuticular wax biosynthesis. J. Exp. Bot. 2022, 73, 2799–2816. [Google Scholar] [CrossRef]
- Aharoni, A.; Dixit, S.; Jetter, R.; Thoenes, E.; van Arkel, G.; Pereira, A. The SHINE clade of AP2 domain transcription factors activates wax biosynthesis, alters cuticle properties, and confers drought tolerance when overexpressed in Arabidopsis. Plant Cell 2004, 16, 2463–2480. [Google Scholar] [CrossRef] [Green Version]
- Meng, S.; Cao, Y.; Li, H.; Bian, Z.; Wang, D.; Lian, C.; Yin, W.; Xia, X. PeSHN1 regulates water-use efficiency and drought tolerance by modulating wax biosynthesis in poplar. Tree Physiol. 2019, 39, 1371–1386. [Google Scholar] [CrossRef]
- Grünhofer, P.; Herzig, L.; Schreiber, L. Leaf morphology, wax composition, and residual (cuticular) transpiration of four poplar clones. Trees 2022, 36, 645–658. [Google Scholar] [CrossRef]
- Grünhofer, P.; Herzig, L.; Sent, S.; Zeisler-Diehl, V.V.; Schreiber, L. Increased cuticular wax deposition does not change residual foliar transpiration. Plant Cell Environ. 2022, 45, 1157–1171. [Google Scholar] [CrossRef]
- Yang, J.; Isabel Ordiz, M.; Jaworski, J.G.; Beachy, R.N. Induced accumulation of cuticular waxes enhances drought tolerance in Arabidopsis by changes in development of stomata. Plant Physiol. Biochem. 2011, 49, 1448–1455. [Google Scholar] [CrossRef] [PubMed]
- Djemal, R.; Khoudi, H. TdSHN1, a WIN1/SHN1-type transcription factor, imparts multiple abiotic stress tolerance in transgenic tobacco. Environ. Exp. Bot. 2016, 131, 89–100. [Google Scholar] [CrossRef]
- Bi, H.; Shi, J.; Kovalchuk, N.; Luang, S.; Bazanova, N.; Chirkova, L.; Zhang, D.; Shavrukov, Y.; Stepanenko, A.; Tricker, P.; et al. Overexpression of the TaSHN1 transcription factor in bread wheat leads to leaf surface modifications, improved drought tolerance, and no yield penalty under controlled growth conditions. Plant Cell Environ. 2018, 41, 2549–2566. [Google Scholar] [CrossRef] [PubMed]
- Schouten, H.J.; Krens, F.A.; Jacobsen, E. Cisgenic plants are similar to traditionally bred plants: International regulations for genetically modified organisms should be altered to exempt cisgenesis. EMBO Rep. 2006, 7, 750–753. [Google Scholar] [CrossRef]
- Brunner, I.; Herzog, C.; Dawes, M.A.; Arend, M.; Sperisen, C. How tree roots respond to drought. Front. Plant Sci. 2015, 6, 547. [Google Scholar] [CrossRef] [Green Version]
- Karlova, R.; Boer, D.; Hayes, S.; Testerink, C. Root plasticity under abiotic stress. Plant Physiol. 2021, 187, 1057–1070. [Google Scholar] [CrossRef]
- Gries, D.; Zeng, F.; Foetzki, A.; Arndt, S.K.; Bruelheide, H.; Thomas, F.M.; Zhang, X.; Runge, M. Growth and water relations of Tamarix ramosissima and Populus euphratica on Taklamakan desert dunes in relation to depth to a permanent water table. Plant Cell Environ. 2003, 26, 725–736. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhao, C.; Li, J.; Liu, Z.; Wang, J. Root plasticity of Populus euphratica seedlings in response to different water table depths and contrasting sediment types. PLoS ONE 2015, 10, e0118691. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.; Morgenstern, K.; Heilig, D.; Heil, B.; Kovács, G.; Leibing, C.; Krabel, D. Biomass allocation and root characteristics of early-stage poplars (Populus spp.) for assessing their water-deficit response during SRC establishment. Bioenergy Res. 2021, 14, 385–398. [Google Scholar] [CrossRef]
- Waadt, R.; Seller, C.A.; Hsu, P.-K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell. Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, Y.; Wang, X.; Han, X.; An, Y.; Lin, S.; Shen, C.; Wen, J.L.; Liu, C.; Yin, W.; et al. Root-specific NF-Y family transcription factor, PdNF-YB21, positively regulates root growth and drought resistance by abscisic acid-mediated indoylacetic acid transport in Populus. New Phytol. 2020, 227, 407–426. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Qin, J.; Tong, S.; Wang, W.; Jiang, Y. One AP2/ERF transcription factor positively regulates Pi uptake and drought tolerance in poplar. Int. J. Mol. Sci. 2022, 23, 5241. [Google Scholar] [CrossRef]
- Xue, L.J.; Frost, C.J.; Tsai, C.J.; Harding, S.A. Drought response transcriptomes are altered in poplar with reduced tonoplast sucrose transporter expression. Sci. Rep. 2016, 6, 33655. [Google Scholar] [CrossRef] [Green Version]
- Dash, M.; Yordanov, Y.S.; Georgieva, T.; Tschaplinski, T.J.; Yordanova, E.; Busov, V. Poplar PtabZIP1-like enhances lateral root formation and biomass growth under drought stress. Plant J. 2017, 89, 692–705. [Google Scholar] [CrossRef]
- Dash, M.; Yordanov, Y.S.; Georgieva, T.; Wei, H.; Busov, V. Gene network analysis of poplar root transcriptome in response to drought stress identifies a PtaJAZ3PtaRAP2.6-centered hierarchical network. PLoS ONE 2018, 13, e0208560. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wang, L.; Zhang, J.; Li, J.; Zheng, H.; Chen, J.; Lu, M. WUSCHEL-related Homeobox genes in Populus tomentosa: Diversified expression patterns and a functional similarity in adventitious root formation. BMC Genom. 2014, 15, 296. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Xie, W.; Huang, M. Two WUSCHEL-related Homeobox genes, PeWOX11a and PeWOX11b, are involved in adventitious root formation of poplar. Physiol. Plant. 2015, 155, 446–456. [Google Scholar] [CrossRef]
- Wang, L.Q.; Li, Z.; Wen, S.S.; Wang, J.N.; Zhao, S.T.; Lu, M.Z. WUSCHEL-related Homeobox gene PagWOX11/12a responds to drought stress by enhancing root elongation and biomass growth in poplar. J Exp. Bot. 2020, 71, 1503–1513. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, G.; Guzmán-Guzmán, P.; Parra-Cota, F.I.; de los Santos-Villalobos, S.; Orozco-Mosqueda, M.D.C.; Glick, B.R. Plant Growth Stimulation by Microbial Consortia. Agronomy 2021, 11, 219. [Google Scholar] [CrossRef]
- Caddell, D.F.; Deng, S.; Coleman-Derr, D. Role of the Plant Root Microbiome in Abiotic Stress Tolerance. In Seed Endophytes: Biology and Biotechnology; Verma, S., White, J., Jr., Eds.; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Shakya, M.; Gottel, N.; Castro, H.; Yang, Z.K.; Gunter, L.; Labbé, J.; Muchero, W.; Bonito, G.; Vilgalys, R.; Tuskan, G.; et al. A Multifactor Analysis of Fungal and Bacterial Community Structure in the Root Microbiome of Mature Populus deltoides Trees. PLoS ONE 2013, 8, e76382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cregger, M.A.; Veach, A.M.; Yang, Z.K.; Crouch, M.J.; Vilgalys, R.; Tuskan, G.A.; Schadt, C.W. The Populus Holobiont: Dissecting the Effects of Plant Niches and Genotype on the Microbiome. Microbiome 2018, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Siemens, J.A.; Zwiazek, J.J. Root Hydraulic Properties and Growth of Balsam Poplar (Populus balsamifera) Mycorrhizal with Hebeloma crustuliniforme and Wilcoxina mikolae var. mikolae. Mycorrhiza 2008, 18, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Marjanović, Ž.; Uwe, N.; Hamp, R. Mycorrhiza Formation Enhances Adaptive Response of Hybrid Poplar to Drought. Ann. N. Y. Acad. Sci. 2005, 1048, 496–499. [Google Scholar] [CrossRef]
- Bonito, G.; Benucci, G.M.N.; Hameed, K.; Weighill, D.; Jones, P.; Chen, K.H.; Jacobson, D.; Schadt, C.; Vilgalys, R. Fungal-Bacterial Networks in the Populus Rhizobiome Are Impacted by Soil Properties and Host Genotype. Front. Microbiol. 2019, 10, 481. [Google Scholar] [CrossRef] [Green Version]
- Veach, A.M.; Chen, H.; Yang, Z.K.; Labbe, A.D.; Engle, N.L.; Tschaplinski, T.J.; Schadt, C.W.; Cregger, M.A. Plant Hosts Modify Belowground Microbial Community Response to Extreme Drought. Msystems 2020, 5, e00092-20. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Dawwam, G.E.; Sehim, A.E.; Li, X.; Wu, J.; Chen, S.; Zhang, D. Drought Stress Triggers Shifts in the Root Microbial Community and Alters Functional Categories in the Microbial Gene Pool. Front. Microbiol. 2021, 12, 3066. [Google Scholar] [CrossRef]
- Kristy, B.; Carrell, A.A.; Johnston, E.; Cumming, J.R.; Klingeman, D.M.; Gwinn, K.; Syring, K.C.; Skalla, C.; Emrich, S.; Cregger, M.A. Chronic Drought Differentially Alters the Belowground Microbiome of Drought-Tolerant and Drought-Susceptible Genotypes of Populus Trichocarpa. Phytobiomes J. 2022, 6, 317–330. [Google Scholar] [CrossRef]
- Kurpisz, B.; Pawłowski, T.A. Epigenetic mechanisms of tree responses to climatic changes. Int. J. Mol. Sci. 2022, 23, 13412. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lang, Z.; Zhu, J.K. Dynamics and function of DNA methylation in plants. Nat. Rev. Mol. Cell Biol. 2018, 19, 489–506. [Google Scholar] [CrossRef]
- Crisp, P.A.; Ganguly, D.; Eichten, S.R.; Borevitz, J.O.; Pogson, B.J. Reconsidering Plant Memory: Intersections between Stress Recovery, RNA Turnover, and Epigenetics. Sci. Adv. 2016, 2, e1501340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lämke, J.; Bäurle, I. Epigenetic and Chromatin-Based Mechanisms in Environmental Stress Adaptation and Stress Memory in Plants. Genome Biol. 2017, 18, 124. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Fromm, M.; Avramova, Z. Multiple Exposures to Drought “train” Transcriptional Responses in Arabidopsis. Nat. Commun. 2012, 3, 740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnsen, Ø.; Fossdal, C.G.; Nagy, N.; MØlmann, J.; Dæhlen, O.G.; SkrØppa, T. Climatic Adaptation in Picea abies Progenies Is Affected by the Temperature during Zygotic Embryogenesis and Seed Maturation. Plant Cell Environ. 2005, 28, 1090–1102. [Google Scholar] [CrossRef]
- Sow, M.D.; Allona, I.; Ambroise, C.; Conde, D.; Fichot, R.; Gribkova, S.; Jorge, V.; Le-Provost, G.; Pâques, L.; Plomion, C.; et al. Epigenetics in Forest Trees: State of the Art and Potential Implications for Breeding and Management in a Context of Climate Change. Adv. Bot. Res. 2018, 88, 387–453. [Google Scholar] [CrossRef]
- Raj, S.; Bräutigam, K.; Hamanishi, E.T.; Wilkins, O.; Thomas, B.R.; Schroeder, W.; Mansfield, S.D.; Plant, A.L.; Campbell, M.M. Clone History Shapes Populus Drought Responses. Proc. Natl. Acad. Sci. USA 2011, 108, 12521–12526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bräutigam, K.; Vining, K.J.; Lafon-Placette, C.; Fossdal, C.G.; Mirouze, M.; Marcos, J.G.; Fluch, S.; Fraga, M.F.; Guevara, M.Á.; Abarca, D.; et al. Epigenetic Regulation of Adaptive Responses of Forest Tree Species to the Environment. Ecol. Evol. 2013, 3, 399–415. [Google Scholar] [CrossRef]
- Gourcilleau, D.; Bogeat-Triboulot, M.B.; Le Thiec, D.; Lafon-Placette, C.; Delaunay, A.; El-Soud, W.A.; Brignolas, F.; Maury, S. DNA Methylation and Histone Acetylation: Genotypic Variations in Hybrid Poplars, Impact of Water Deficit and Relationships with Productivity. Ann. For. Sci. 2010, 67, 208. [Google Scholar] [CrossRef] [Green Version]
- Le Gac, A.L.; Lafon-Placette, C.; Chauveau, D.; Segura, V.; Delaunay, A.; Fichot, R.; Marron, N.; Le Jan, I.; Berthelot, A.; Bodineau, G.; et al. Winter-Dormant Shoot Apical Meristem in Poplar Trees Shows Environmental Epigenetic Memory. J. Exp. Bot. 2018, 69, 4821–4837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafon-Placette, C.; Le Gac, A.L.; Chauveau, D.; Segura, V.; Delaunay, A.; Lesage-Descauses, M.C.; Hummel, I.; Cohen, D.; Jesson, B.; Le Thiec, D.; et al. Changes in the Epigenome and Transcriptome of the Poplar Shoot Apical Meristem in Response to Water Availability Affect Preferentially Hormone Pathways. J. Exp. Bot. 2018, 69, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Sow, M.D.; Segura, V.; Chamaillard, S.; Jorge, V.; Delaunay, A.; Lafon-Placette, C.; Fichot, R.; Faivre-Rampant, P.; Villar, M.; Brignolas, F.; et al. Narrow-Sense Heritability and P ST Estimates of DNA Methylation in Three Populus nigra L. Populations under Contrasting Water Availability. Tree Genet. Genomes 2018, 14, 1–12. [Google Scholar] [CrossRef]
- Liang, D.; Zhang, Z.; Wu, H.; Huang, C.; Shuai, P.; Ye, C.Y.; Tang, S.; Wang, Y.; Yang, L.; Wang, J.; et al. Single-Base-Resolution Methylomes of Populus trichocarpa Reveal the Association between DNA Methylation and Drought Stress. BMC Genet. 2014, 15, S9. [Google Scholar] [CrossRef] [Green Version]
- Vining, K.J.; Pomraning, K.R.; Wilhelm, L.J.; Priest, H.D.; Pellegrini, M.; Mockler, T.C.; Freitag, M.; Strauss, S.H. Dynamic DNA Cytosine Methylation in the Populus trichocarpa Genome: Tissue-Level Variation and Relationship to Gene Expression. BMC Genom. 2012, 13, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Georgii, E.; Kugler, K.; Pfeifer, M.; Vanzo, E.; Block, K.; Domagalska, M.A.; Jud, W.; Elgawad, H.A.; Asard, H.; Reinhardt, R.; et al. The Systems Architecture of Molecular Memory in Poplar after Abiotic Stress. Plant Cell 2019, 31, 346–367. [Google Scholar] [CrossRef]
- Sow, M.D.; Le Gac, A.L.; Fichot, R.; Lanciano, S.; Delaunay, A.; Le Jan, I.; Lesage-Descauses, M.C.; Citerne, S.; Caius, J.; Brunaud, V.; et al. RNAi Suppression of DNA Methylation Affects the Drought Stress Response and Genome Integrity in Transgenic Poplar. New Phytol. 2021, 232, 80–97. [Google Scholar] [CrossRef]
- Holbrook, N.M.; Zwieniecki, M.A. Transporting Water to the Tops of Trees. Phys. Today 2008, 61, 76–77. [Google Scholar] [CrossRef]
- Tyree, M.T.; Sperry, J.S. Vulnerability of Xylem to Cavitation and Embolism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 19–36. [Google Scholar] [CrossRef]
- Davis, S.D.; Ewers, F.W.; Sperry, J.S.; Portwood, K.A.; Crocker, M.C.; Adams, G.C. Shoot Dieback during Prolonged Drought in Ceanothus (Rhamnaceae) Chaparral of California: A Possible Case of Hydraulic Failure. Am. J. Bot. 2002, 89, 820–828. [Google Scholar] [CrossRef]
- Wason, J.; Bouda, M.; Lee, E.F.; McElrone, A.J.; Phillips, R.J.; Shackel, K.A.; Matthews, M.A.; Brodersen, C. Xylem Network Connectivity and Embolism Spread in Grapevine (Vitis vinifera L.). Plant. Physiol. 2021, 186, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Mantova, M.; Menezes-Silva, P.E.; Badel, E.; Cochard, H.; Torres-Ruiz, J.M. The Interplay of Hydraulic Failure and Cell Vitality Explains Tree Capacity to Recover from Drought. Physiol. Plant. 2021, 172, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Adams, H.D.; Zeppel, M.J.B.; Anderegg, W.R.L.; Hartmann, H.; Landhäusser, S.M.; Tissue, D.T.; Huxman, T.E.; Hudson, P.J.; Franz, T.E.; Allen, C.D.; et al. A Multi-Species Synthesis of Physiological Mechanisms in Drought-Induced Tree Mortality. Nat. Ecol. Evol. 2017, 1, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Jansen, S.; Choat, B.; Pletsers, A. Morphological Variation of Intervessel Pit Membranes and Implications to Xylem Function in Angiosperms. Am. J. Bot. 2009, 96, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Hajek, P.; Leuschner, C.; Hertel, D.; Delzon, S.; Schuldt, B. Trade-Offs between Xylem Hydraulic Properties, Wood Anatomy and Yield in Populus. Mol. Hum. Reprod. 2014, 34, 744–756. [Google Scholar] [CrossRef]
- Cai, J.; Tyree, M.T. The Impact of Vessel Size on Vulnerability Curves: Data and Models for within-Species Variability in Saplings of Aspen, Populus tremuloides Michx. Plant Cell Environ. 2010, 33, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Losso, A.; Beikircher, B.; Dämon, B.; Kikuta, S.; Schmid, P.; Mayr, S. Xylem Sap Surface Tension May Be Crucial for Hydraulic Safety. Plant Physiol. 2017, 175, 1135–1143. [Google Scholar] [CrossRef] [Green Version]
- Stiller, V.; Sperry, J.S. Cavitation Fatigue and Its Reversal in Sunflower (Helianthus annuus L.). J. Exp. Bot. 2002, 53, 1155–1161. [Google Scholar] [CrossRef] [Green Version]
- Lemaire, C.; Quilichini, Y.; Brunel-Michac, N.; Santini, J.; Berti, L.; Cartailler, J.; Conchon, P.; Badel, É.; Herbette, S. Plasticity of the Xylem Vulnerability to Embolism in Populus tremula × alba Relies on Pit Quantity Properties Rather than on Pit Structure. Tree Physiol. 2021, 41, 1384–1399. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Klein, T.; Bartlett, M.; Sack, L.; Pellegrini, A.F.A.; Choat, B.; Jansen, S. Meta-Analysis Reveals That Hydraulic Traits Explain Cross-Species Patterns of Drought-Induced Tree Mortality across the Globe. Proc. Natl. Acad. Sci. USA 2016, 113, 5024–5029. [Google Scholar] [CrossRef] [Green Version]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; López, R.; Medlyn, B.E. Triggers of Tree Mortality under Drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef]
- Fichot, R.; Brignolas, F.; Cochard, H.; Ceulemans, R. Vulnerability to Drought-Induced Cavitation in Poplars: Synthesis and Future Opportunities. Plant Cell Environ. 2015, 38, 1233–1251. [Google Scholar] [CrossRef]
- Eckenwalder, J.E. Systematics and Evolution of Populus. In Biology of Populus and Its Implications for Management and Conservation; Stettler, F., Bradshaw, H.D., Jr., Heilman, P.E., Hinckley, T.M., Eds.; NRC Research Press: Ottawa, ON, Canada, 1996; pp. 7–32. [Google Scholar]
- Hukin, D.; Cochard, H.; Dreyer, E.; Le Thiec, D.; Bogeat-Triboulot, M.B. Cavitation Vulnerability in Roots and Shoots: Does Populus euphratica Oliv., a Poplar from Arid Areas of Central Asia, Differ from Other Poplar Species? J. Exp. Bot. 2005, 56, 2003–2010. [Google Scholar] [CrossRef]
- Sperry, J.S.; Perry, A.H.; Sullivan, J.E.M. Pit Membrane Degradation and Air-Embolism Formation in Ageing Xylem Vessels of Populus tremuloides Michx. J. Exp. Bot. 1991, 42, 1399–1406. [Google Scholar] [CrossRef]
- Tomasella, M.; Casolo, V.; Natale, S.; Petruzzellis, F.; Kofler, W.; Beikircher, B.; Mayr, S.; Nardini, A. Shade-Induced Reduction of Stem Nonstructural Carbohydrates Increases Xylem Vulnerability to Embolism and Impedes Hydraulic Recovery in Populus nigra. New Phytol. 2021, 231, 108–121. [Google Scholar] [CrossRef]
- de Baerdemaeker, N.J.F.; Salomón, R.L.; de Roo, L.; Steppe, K. Sugars from Woody Tissue Photosynthesis Reduce Xylem Vulnerability to Cavitation. New Phytol. 2017, 216, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Brodribb, T.J.; Bowman, D.J.M.S.; Nichols, S.; Delzon, S.; Burlett, R. Xylem Function and Growth Rate Interact to Determine Recovery Rates after Exposure to Extreme Water Deficit. New Phytol. 2010, 188, 533–542. [Google Scholar] [CrossRef]
- Sperry, J.S.; Holbrook, N.M.; Zimmermann, M.H.; Tyree, M.T. Spring Filling of Xylem Vessels in Wild Grapevine. Plant Physiol. 1987, 83, 414–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Alejandra Equiza, M.; Deng, X.; Tyree, M.T. Recovery of Populus tremuloides Seedlings Following Severe Drought Causing Total Leaf Mortality and Extreme Stem Embolism. Physiol. Plant. 2010, 140, 246–257. [Google Scholar] [CrossRef]
- Love, D.M.; Sperry, J.S. In Situ Embolism Induction Reveals Vessel Refilling in a Natural Aspen Stand. Tree Physiol. 2018, 38, 1006–1015. [Google Scholar] [CrossRef] [PubMed]
- Brodersen, C.R.; McElrone, A.J.; Choat, B.; Matthews, M.A.; Shackel, K.A. The Dynamics of Embolism Repair in Xylem: In Vivo Visualizations Using High-Resolution Computed Tomography. Plant Physiol. 2010, 154, 1088–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trifilò, P.; Kiorapostolou, N.; Petruzzellis, F.; Vitti, S.; Petit, G.; Lo Gullo, M.A.; Nardini, A.; Casolo, V. Hydraulic Recovery from Xylem Embolism in Excised Branches of Twelve Woody Species: Relationships with Parenchyma Cells and Non-Structural Carbohydrates. Plant Physiol. Biochem. 2019, 139, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Martorell, S.; Diaz-Espejo, A.; Medrano, H.; Ball, M.C.; Choat, B. Rapid Hydraulic Recovery in Eucalyptus Pauciflora after Drought: Linkages between Stem Hydraulics and Leaf Gas Exchange. Plant Cell Environ. 2014, 37, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Salleo, S.; Lo Gullo, M.A.; Trifilò, P.; Nardini, A. New Evidence for a Role of Vessel-Associated Cells and Phloem in the Rapid Xylem Refilling of Cavitated Stems of Laurus nobilis L. Plant Cell Environ. 2004, 27, 1065–1076. [Google Scholar] [CrossRef]
- Zwieniecki, M.A.; Holbrook, N.M. Confronting Maxwell’s Demon: Biophysics of Xylem Embolism Repair. Trends Plant Sci 2009, 14, 530–534. [Google Scholar] [CrossRef]
- Secchi, F.; Zwieniecki, M.A. Accumulation of Sugars in the Xylem Apoplast Observed under Water Stress Conditions Is Controlled by Xylem PH. Plant Cell Environ. 2016, 39, 2350–2360. [Google Scholar] [CrossRef]
- Secchi, F.; Pagliarani, C.; Cavalletto, S.; Petruzzellis, F.; Tonel, G.; Savi, T.; Tromba, G.; Obertino, M.M.; Lovisolo, C.; Nardini, A.; et al. Chemical Inhibition of Xylem Cellular Activity Impedes the Removal of Drought-Induced Embolisms in Poplar Stems—New Insights from Micro-CT Analysis. New Phytol. 2021, 229, 820–830. [Google Scholar] [CrossRef]
- Pagliarani, C.; Casolo, V.; Ashofteh Beiragi, M.; Cavalletto, S.; Siciliano, I.; Schubert, A.; Gullino, M.L.; Zwieniecki, M.A.; Secchi, F. Priming Xylem for Stress Recovery Depends on Coordinated Activity of Sugar Metabolic Pathways and Changes in Xylem Sap PH. Plant Cell Environ. 2019, 42, 1775–1787. [Google Scholar] [CrossRef] [Green Version]
- Secchi, F.; Zwieniecki, M.A. Sensing Embolism in Xylem Vessels: The Role of Sucrose as a Trigger for Refilling. Plant Cell Environ. 2011, 34, 514–524. [Google Scholar] [CrossRef]
- Secchi, F.; Zwieniecki, M.A. Analysis of Xylem Sap from Functional (Nonembolized) and Nonfunctional (Embolized) Vessels of Populus nigra: Chemistry of Refilling. Plant Physiol. 2012, 160, 955–964. [Google Scholar] [CrossRef] [Green Version]
- Secchi, F.; Zwieniecki, M.A. Patterns of PIP Gene Expression in Populus trichocarpa during Recovery from Xylem Embolism Suggest a Major Role for the PIP1 Aquaporin Subfamily as Moderators of Refilling Process. Plant Cell Environ. 2010, 33, 1285–1297. [Google Scholar] [CrossRef]
- Almeida-Rodriguez, A.M.; Cooke, J.E.K.; Yeh, F.; Zwiazek, J.J. Functional Characterization of Drought-Responsive Aquaporins in Populus balsamifera and Populus simonii × balsamifera Clones with Different Drought Resistance Strategies. Physiol. Plant. 2010, 140, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Almeida-Rodriguez, A.M.; Hacke, U.G. Cellular Localization of Aquaporin mRNA in Hybrid Poplar Stems. Am. J. Bot. 2012, 99, 1249–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laur, J.; Hacke, U.G. The Role of Water Channel Proteins in Facilitating Recovery of Leaf Hydraulic Conductance from Water Stress in Populus trichocarpa. PLoS ONE 2014, 9, e111751. [Google Scholar] [CrossRef] [Green Version]
- Secchi, F.; Zwieniecki, M.A. Down-Regulation of Plasma Intrinsic Protein1 Aquaporin in Poplar Trees Is Detrimental to Recovery from Embolism. Plant Physiol. 2014, 164, 1789–1799. [Google Scholar] [CrossRef] [Green Version]
- Leng, H.; Lu, M.; Wan, X. Variation in Embolism Occurrence and Repair along the Stem in Drought-Stressed and Re-Watered Seedlings of a Poplar Clone. Physiol. Plant. 2013, 147, 329–339. [Google Scholar] [CrossRef]
- Yu, D.; Janz, D.; Zienkiewicz, K.; Herrfurth, C.; Feussner, I.; Chen, S.; Polle, A. Wood Formation under Severe Drought Invokes Adjustment of the Hormonal and Transcriptional Landscape in Poplar. Int. J. Mol. Sci. 2021, 22, 9899. [Google Scholar] [CrossRef] [PubMed]
- Popko, J.; Hänsch, R.; Mendel, R.R.; Polle, A.; Teichmann, T. The Role of Abscisic Acid and Auxin in the Response of Poplar to Abiotic Stress. Plant. Biol. 2010, 12, 242–258. [Google Scholar] [CrossRef]
- Xu, C.; Fu, X.; Liu, R.; Guo, L.; Ran, L.; Li, C.; Tian, Q.; Jiao, B.; Wang, B.; Luo, K. PtoMYB170 Positively Regulates Lignin Deposition during Wood Formation in Poplar and Confers Drought Tolerance in Transgenic Arabidopsis. Tree Physiol. 2017, 37, 1713–1726. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Fan, Y.; Du, S.; Zhao, K.; Liu, Q.; Yao, W.; Zheng, T.; Han, Y. PtaERF194 Inhibits Plant Growth and Enhances Drought Tolerance in Poplar. Tree Physiol. 2022, 42, 1–15. [Google Scholar] [CrossRef]
- Xie, Z.; Nolan, T.M.; Jiang, H.; Yin, Y. AP2/ERF Transcription Factor Regulatory Networks in Hormone and Abiotic Stress Responses in Arabidopsis. Front. Plant Sci. 2019, 10, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocozza, C.; Giovannelli, A.; Traversi, M.L.; Castro, G.; Cherubini, P.; Tognetti, R. Do Tree-Ring Traits Reflect Different Water Deficit Responses in Young Poplar Clones (Populus × canadensis Mönch “I-214” and P. Deltoides ’Dvina’)? Trees Struct. Funct. 2011, 25, 975–985. [Google Scholar] [CrossRef]
- Huda, A.S.M.; Koubaa, A.; Cloutier, A.; Hernández, R.E.; Périnet, P.; Fortin, Y. Phenotypic and Genotypic Correlations for Wood Properties of Hybrid Poplar Clones of Southern Quebec. Forests 2018, 9, 140. [Google Scholar] [CrossRef] [Green Version]
- Arend, M.; Fromm, J. Seasonal Change in the Drought Response of Wood Cell Development in Poplar. Tree Physiol. 2007, 27, 985–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steenackers, J.; Steenackers, M.; Steenackers, V.; Stevens, M. Poplar Diseases, consequences on growth and wood quality. Biomass Bioenergy 1996, 10, 267–274. [Google Scholar] [CrossRef]
- Pinon, J.; Poissonnier, M. Etude épidemiologique du Marssonina brunnea (Ell. et Ev.) P. Magn. Eur. J. For. Pathol. 1975, 5, 97–111. [Google Scholar] [CrossRef]
- Castellani, E. Influence des factors climatiques sur les infections des peupliers euraméricains par Marssonina brunnea (Ell. et Ev.) P.Magn. Phytopathol. Mediterr. 1966, 5, 41–52. [Google Scholar]
- Taris, B. Les Maladies des Peupliers; Association Forêt-Cellulose: Champagne sur Seine, France, 1981. [Google Scholar]
- Ullah, C.; Tsai, C.J.; Unsicker, S.B.; Xue, L.; Reichelt, M.; Gershenzon, J.; Hammerbacher, A. Salicylic acid activates poplar defense against the biotrophic rust fungus Melampsora larici-populina via increased biosynthesis of catechin and proanthocyanidins. New Phytol. 2019, 221, 960–975. [Google Scholar] [CrossRef] [Green Version]
- Yarwood, C.E. Predisposition. In Plant Pathology; Horsfall, J.G., Dimond, A.E., Eds.; Academic Press: New York, NY, USA; London, UK, 1959; pp. 521–562. [Google Scholar]
- Oliva, J.; Redondo, M.A.; Stenlid, J. Functional Ecology of Forest Disease. Annu. Rev. Phytopathol. 2020, 58, 343–361. [Google Scholar] [CrossRef]
- Brader, G.; Compant, S.; Vescio, K.; Mitter, B.; Trognitz, F.; Ma, L.J.; Sessitsch, A. Ecology and Genomic Insights into Plant-Pathogenic and Plant-Nonpathogenic Endophytes. Annu. Rev. Phytopathol. 2017, 55, 61–83. [Google Scholar] [CrossRef]
- Vayssier-Taussat, M.; Albina, E.; Citti, C.; Cosson, J.F.; Jacques, M.A.; Lebrun, M.H.; Le Loir, Y.; Ogliastro, M.; Petit, M.A.; Roumagnac, P.; et al. Shifting the paradigm from pathogens to pathobiome new concepts in the light of meta-omics. Front. Cell. Infect. Microbiol. 2014, 5, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guyon, J.C.; Jacobi, W.R.; McIntyre, G.A. Effects of environmental stress on the development of Cytospora canker of aspen. Plant Dis. 1996, 80, 1320–1326. [Google Scholar] [CrossRef]
- Cellerino, G.P.; Gennaro, M. Drought as predisposing factor in oak decline. In Decline of Oak Species in Italy: Problems and Perspectives; Ragazzi, A., Dellavalle, I., Moricca, S., Capretti, P., Raddi, P., Eds.; Accademia Italiana di Scienze Forestali: Firenze, Italy, 2000; pp. 157–175. [Google Scholar]
- Desprez-Loustau, M.L.; Marçais, B.; Nageleisen, L.M.; Piou, D.; Vannini, A. Interactive effects of drought and pathogens in forest trees. Ann. For. Sci. 2006, 63, 597–612. [Google Scholar] [CrossRef] [Green Version]
- Bélanger, R.R.; Manion, P.D.; Griffin, D.H. Amino acid content of water-stressed plantlets of Populus tremuloides clones in relation to clonal susceptibility to Hypoxylon mammatum in vitro. Can. J. Bot. 1990, 68, 26–29. [Google Scholar] [CrossRef]
- Kruger, B.M.; Manion, P.D. Antifungal compounds in aspen: Effect of water stress. Can. J. Bot. 1994, 72, 454–460. [Google Scholar] [CrossRef]
- Vannini, A.; Valentini, R. Influence of water relations on Quercus cerris–Hypoxylon mediterraneum interaction: A model of drought-induced susceptibility to a weakness parasite. Tree Physiol. 1994, 14, 129–139. [Google Scholar] [CrossRef]
- Manion, P.D. Tree Disease Concepts; Prentice Hall: Englewood Cliffs, NJ, USA, 1991. [Google Scholar]
- Charles, J.G.; Nef, L.; Allegro, G.; Collins, C.M.; Delplanque, A.; Gimenez, R.; Höglund, S.; Jiafu, H.; Larsson, S.; Luo, Y. Insect and other pests of poplars and willows. In Poplars and willows—Trees for Society and Environment; Isebrands, J.G., Richardson, J., Eds.; CAB International and FAO: Roma, Italy, 2014; pp. 459–526. [Google Scholar]
- Cornelissen, T.; Wilson Fernandes, G.; Vasconcellos-Neto, J. Size does matter: Variation in herbivory between and within plants and the plant vigor hypothesis. Oikos 2008, 117, 1121–1130. [Google Scholar] [CrossRef]
- Leybourne, D.J.; Preedy, K.F.; Valentine, T.A.; Bos, J.I.B.; Karley, A.J. Drought has negative consequences on aphid fitness and plant vigor: Insights from a meta-analysis. Ecol. Evol. 2021, 11, 11915–11929. [Google Scholar] [CrossRef]
- Dardeau, F.; Berthier, A.; Feinard-Duranceau, M.; Brignolas, F.; Laurans, F.; Lieutier, F.; Sallé, A. Tree genotype modulates the effects of water deficit on a plant-manipulating aphid. For. Ecol. Manag. 2015, 353, 118–125. [Google Scholar] [CrossRef]
- Gely, C.; Laurance, S.G.W.; Stork, N.E. How do herbivorous insects respond to drought stress in trees? Biol. Rev. 2020, 95, 434–448. [Google Scholar] [CrossRef]
- La Spina, S.; Gregoire, J.C.; Mertens, P.; De-Canniere, C. Impact of poplar water status on leaf-beetle (Chrysomela populi) survival and feeding. Ann. For. Sci. 2010, 67, 209. [Google Scholar] [CrossRef]
- Canelles, Q.; Aquilué, N.; James, P.M.A.; Lawler, J.; Brotons, L. Global review on interactions between insect pests and other forest disturbances. Landsc. Ecol. 2021, 36, 945–972. [Google Scholar] [CrossRef]
- Simonetto, A.; Sperandio, G.; Battisti, A.; Mori, N.; Ciampitti, M.; Cavagna, B.; Bianchi, A.; Gilioli, G. Exploring the main factors influencing habitat preference of Popillia japonica in an area of recent introduction. Ecol. Inform. 2022, 70, 101749. [Google Scholar] [CrossRef]
- Lobos, G.A.; Camargo, A.V.; del Pozo, A.; Araus, J.L.; Ortiz, R.; Doonan, J.H. Editorial: Plant phenotyping and phenomics for plant breeding. Front. Plant. Sci. 2017, 8, 2181. [Google Scholar] [CrossRef] [Green Version]
- Marron, N.; Brignolas, F.; Gao, J.; Johnson, J.D.; Karnosky, D.F.; Polle, A.; Scarascia-Mugnozza, G.; Schroeder, W.R.; Ceulemans, R. Abiotic Stresses. In Poplars and Willows: Trees for Society and the Environment; Richardson, J., Isebrands, J.G., Eds.; FAO-CABI: Wallingford, UK; Boston, MA, USA, 2014; pp. 337–454. [Google Scholar]
- Ucar, Z.; Değermenci, A.S.; Zengin, H.; Bettinger, P. Evaluating the accuracy of remote dendrometers in tree diameter measurements at breast height. Croat. J. For. Eng. 2022, 43, 185–197. [Google Scholar] [CrossRef]
- Rocha, E.; Holzkämper, S. Assessing urban climate effects on Pinus sylvestris with point dendrometers: A case study from Stockholm, Sweden. Trees 2021, 1–10. [Google Scholar] [CrossRef]
- Giovannelli, A.; Deslauriers, A.; Fragnelli, G.; Scaletti, L.; Castro, G.; Rossi, S.; Crivellaro, A. Evaluation of drought response of two poplar clones (Populus × canadensis Mönch ‘I-214′ and P. deltoides Marsh. ’Dvina’) through high resolution analysis of stem growth. J. Exp. Bot. 2007, 58, 2673–2683. [Google Scholar] [CrossRef]
- Chianucci, F.; Giannetti, F.; Tattoni, C.; Puletti, N.; Giorcelli, A.; Bisaglia, C.; Romano, E.; Brambilla, M.; Chiarabaglio, P.; Gennaro, M.; et al. PRECISIONPOP: A multi-scale monitoring system for poplar plantations integrating field, aerial and satellite remote sensing. In Proceedings of the EGU General Assembly 2022, Vienna, Austria, 23–27 May 2022. [Google Scholar]
- Chianucci, F.; Marchino, L.; Bidini, C.; Giorcelli, A.; Coaloa, D.; Chiarabaglio, P.M.; Giannetti, F.; Chirici, G.; Tattoni, C. Dataset of tree inventory and canopy structure in poplar plantations in northern Italy. Ann. Silvicult. Res. 2021, 46, 93–96. [Google Scholar] [CrossRef]
- Ludovisi, R.; Tauro, F.; Salvati, R.; Khoury, S.; Mugnozza, G.S.; Harfouche, A. UAV-based thermal imaging for high-throughput field phenotyping of black poplar response to drought. Front. Plant Sci. 2017, 8, 1681. [Google Scholar] [CrossRef]
- Tauro, F.; Maltese, A.; Giannini, R.; Harfouche, A. Latent heat flux variability and response to drought stress of black poplar: A multi-platform multi-sensor remote and proximal sensing approach to relieve the data scarcity bottleneck. Remote Sens. Environ. 2022, 268, 112771. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosso, L.; Cantamessa, S.; Bergante, S.; Biselli, C.; Fricano, A.; Chiarabaglio, P.M.; Gennaro, M.; Nervo, G.; Secchi, F.; Carra, A. Responses to Drought Stress in Poplar: What Do We Know and What Can We Learn? Life 2023, 13, 533. https://doi.org/10.3390/life13020533
Rosso L, Cantamessa S, Bergante S, Biselli C, Fricano A, Chiarabaglio PM, Gennaro M, Nervo G, Secchi F, Carra A. Responses to Drought Stress in Poplar: What Do We Know and What Can We Learn? Life. 2023; 13(2):533. https://doi.org/10.3390/life13020533
Chicago/Turabian StyleRosso, Laura, Simone Cantamessa, Sara Bergante, Chiara Biselli, Agostino Fricano, Pier Mario Chiarabaglio, Massimo Gennaro, Giuseppe Nervo, Francesca Secchi, and Andrea Carra. 2023. "Responses to Drought Stress in Poplar: What Do We Know and What Can We Learn?" Life 13, no. 2: 533. https://doi.org/10.3390/life13020533