
Differential Accumulation of sHSPs Isoforms during Desiccation of the Resurrection Plant Haberlea rhodopensis Friv. under Optimal and High Temperature

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material, Desiccation and Rehydration
2.2. Determination of Relative Water Content (RWC)
2.3. Chlorophyll Fluorescence
2.4. Protein Extraction and One-Dimensional Sodium Dodecyl Sulphate Polyacrylamide Gel Electrophoresis (1D SDS-PAGE)
2.5. Two-Dimensional Gel Electrophoresis (2-DE)
2.6. Immunoblot Analysis
2.7. Statistical Analysis
3. Results
3.1. Dehydration-Rehydration Cycle of Haberlea rhodopensis
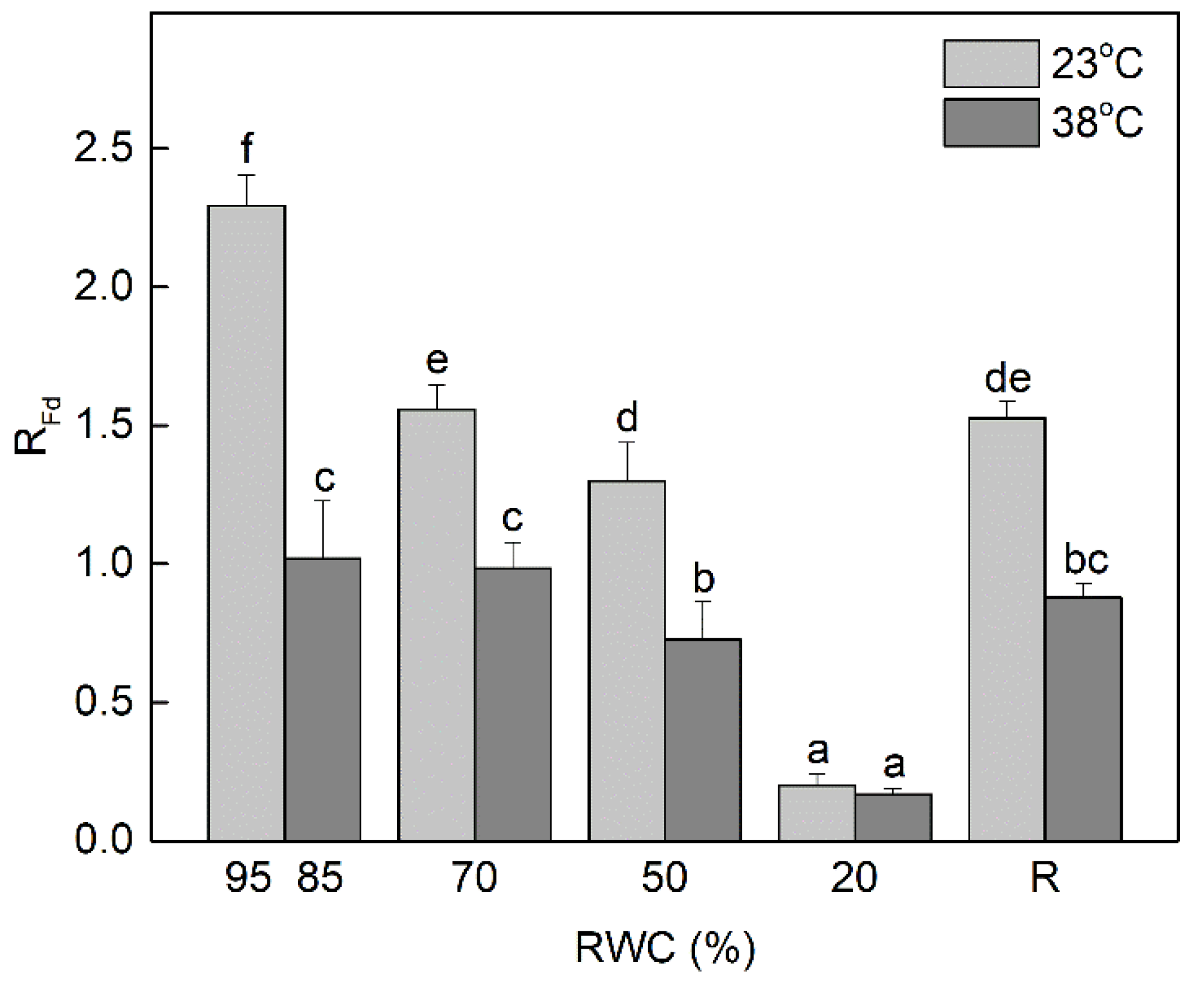
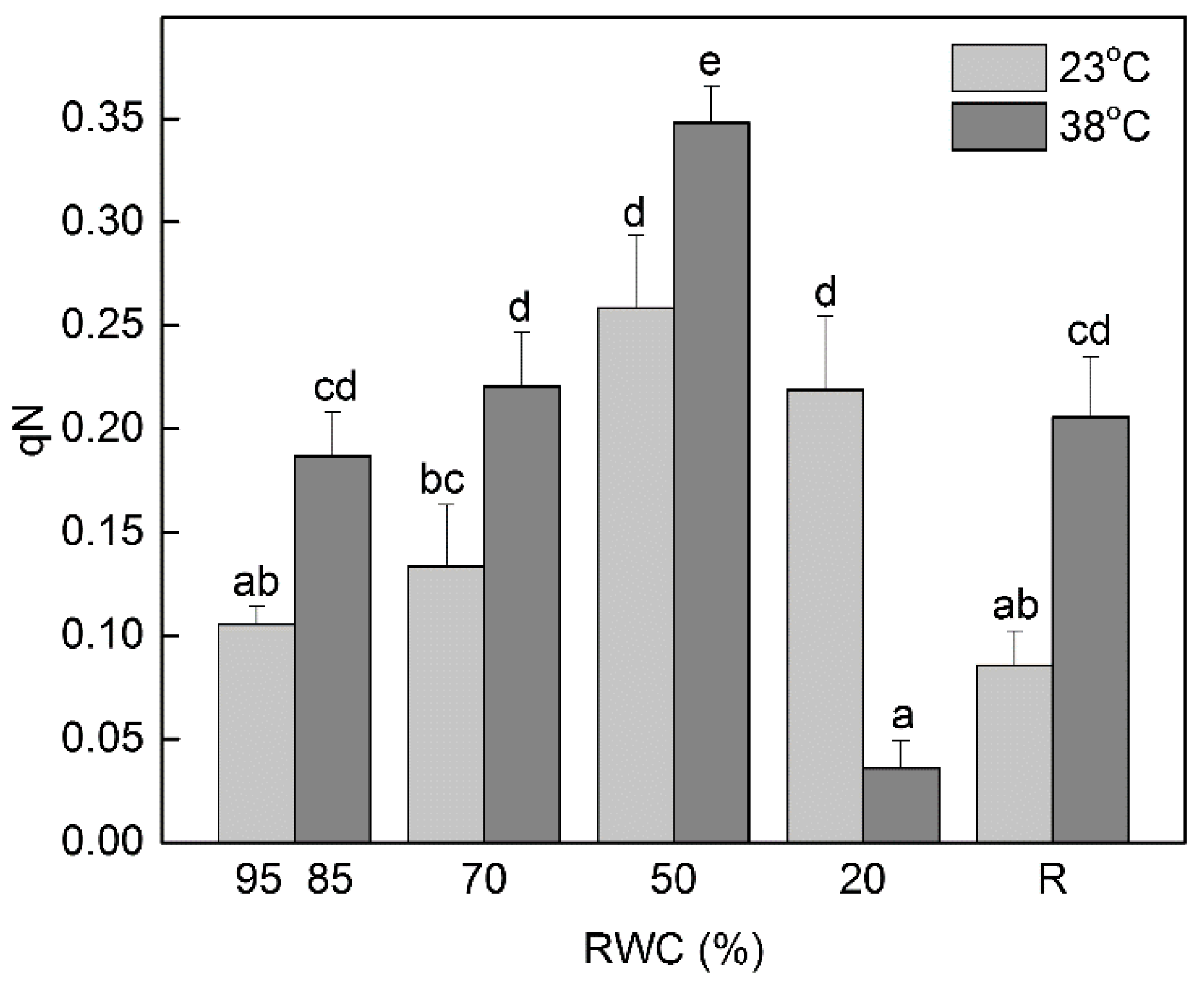
3.2. Changes in the Fluorescence Decrease Ratio (RFd, Vitality Index) and the Extent of Non-Photochemical Quenching under Dehydration at 23 °C and 38 °C
3.3. Differential Accumulation of sHSPs during Dehydration at 23 °C and 38 °C
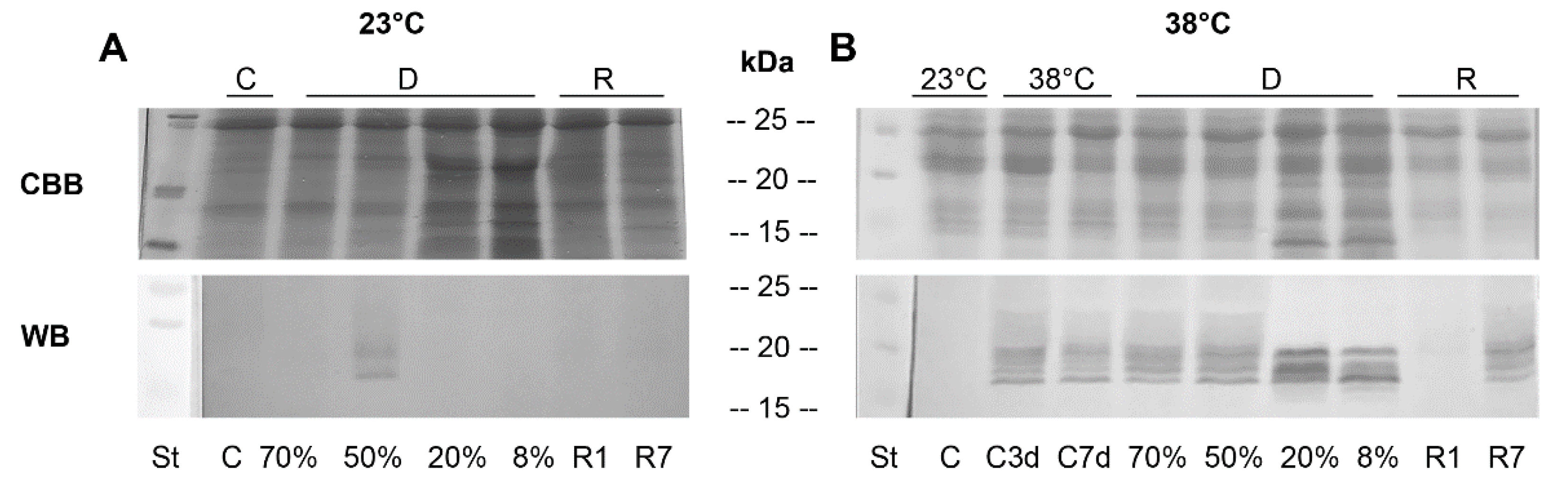
3.3.1. One-Dimensional (1D) SDS-PAGE and Western Blots
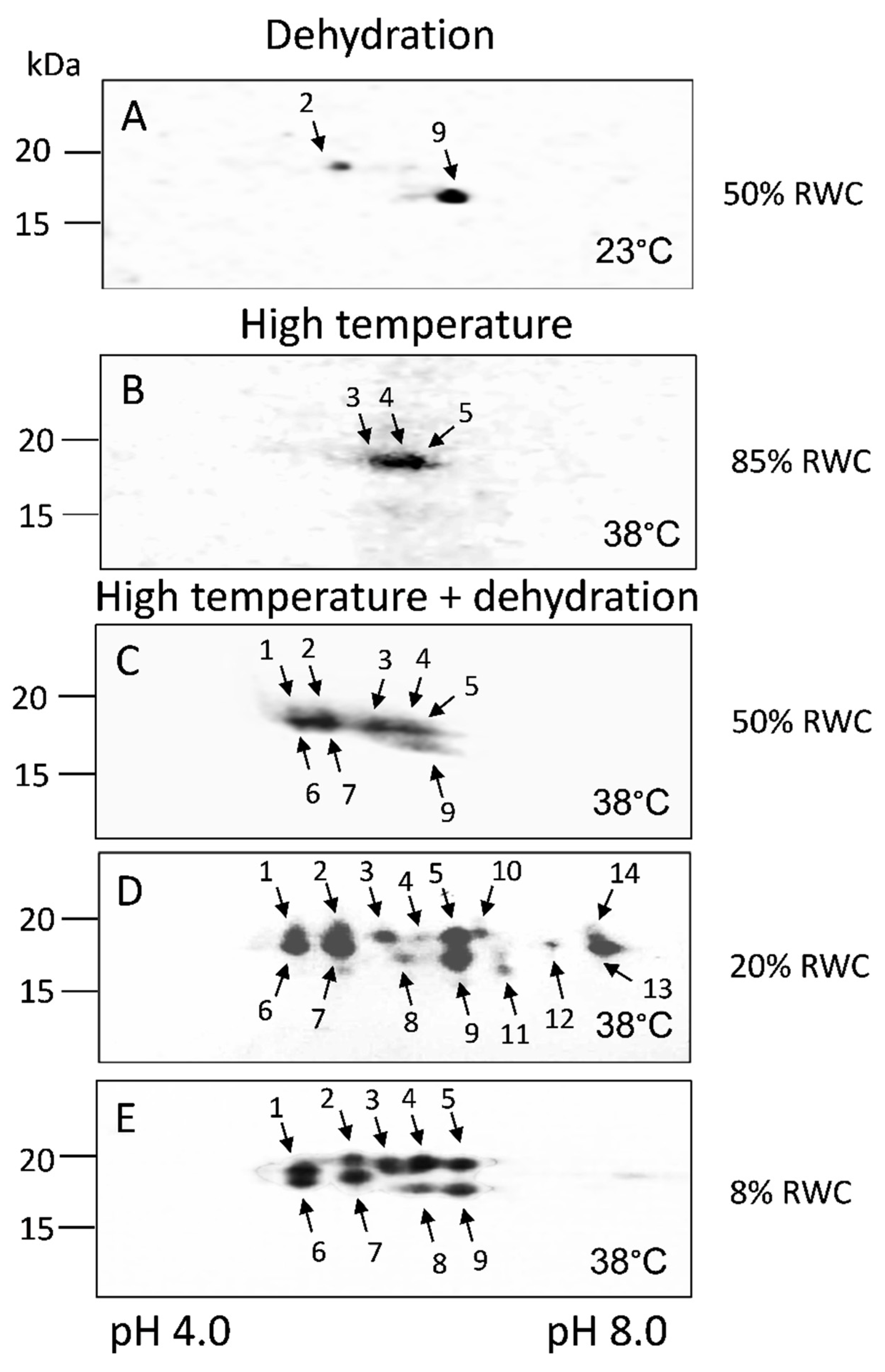
3.3.2. 2D SDS-PAGE and Western Blots
4. Discussion
4.1. Photoprotective Mechanisms under Dehydration at 23 °C and 38 °C
4.2. Accumulation of sHSPs during Dehydration at Optimal and High Temperatures
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farrant, J.M.; Hilhorst, H. Crops for dry environments. Curr. Opin. Biotechn. 2022, 74, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, K.; Maslenkova, L.; Peeva, V.; Markovska, Y.; Stefanov, D.; Tuba, Z. Comparative study on the changes in photosynthetic activity of the homoiochlorophyllous desiccation-tolerant Haberlea rhodopensis and desiccation-sensitive spinach leaves during desiccation and rehydration. Photosynth. Res. 2005, 85, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Petrova, G.; Moyankova, D.; Nishii, K.; Forrest, L.; Tsiripidis, I.; Drouzas, A.D.; Djilianov, D.; Möller, M. The European paleoendemic Haberlea rhodopensis (Gesneriaceae) has an Oligocene origin and a Pleistocene diversification and occurs in a long-persisting refugial area in Southeastern Europe. Int. J. Plant Sci. 2015, 176, 499–514. [Google Scholar] [CrossRef]
- Daskalova, E.; Dontcheva, S.; Yahoubian, G.; Minkov, I.; Toneva, V. A strategy for conservation and investigation of the protected resurrection plant Haberlea rhodopensis Friv. BioRisk 2011, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mundree, S.G.; Baker, B.; Mowla, S.; Peters, S.; Marais, S.; Vander Willigen, C.; Govender, K.; Maredza, A.; Muyanga, S.; Farrant, J.M.; et al. Physiological and molecular insights into drought tolerance. Afr. J. Biotechnol. 2002, 1, 28–38. [Google Scholar] [CrossRef]
- Lyall, R.; Gechev, T. Multi-omics insights into the evolution of angiosperm resurrection plants. Annu. Plant Rev. Online 2020, 3, 77–110. [Google Scholar] [CrossRef]
- Mihailova, G.; Petkova, S.; Stefanov, D.; Georgieva, K. Light dependence of photosynthetic oxygen evolution of Haberlea rhodopensis desiccated at high temperature. Gen. Appl. Plant Physiol. 2009, 35, 111–116. [Google Scholar]
- Mihailova, G.; Petkova, S.; Georgieva, K. Changes in some antioxidant enzyme activities in Haberlea rhodopensis during desiccation at high temperature. Biotechnol. Biotechnol. Equip. 2009, 23, 561–564. [Google Scholar] [CrossRef]
- Mihailova, G.; Petkova, S.; Büchel, C.; Georgieva, K. Desiccation of the resurrection plant Haberlea rhodopensis at high temperature. Photosynth. Res. 2011, 108, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Mihailova, G.; Petkova, S.; Stefanov, D.; Georgieva, K. Effect of Desiccation of the resurrection plant Haberlea rhodopensis at high temperature on the photochemical activity of PSI and PSII. In Photosynthesis Research for Food, Fuel and the Future; Springer: Berlin/Heidelberg, Germany, 2013; pp. 540–543. [Google Scholar] [CrossRef]
- Velitchkova, M.; Doltchinkova, V.; Lazarova, D.; Mihailova, G.; Doncheva, S.; Georgieva, K. Effect of high temperature on dehydration-induced alterations in photosynthetic characteristics of the resurrection plant Haberlea rhodopensis. Photosynthetica 2013, 51, 630–640. [Google Scholar] [CrossRef]
- Howarth, C.J. Genetic improvements of tolerance to high temperature. In Abiotic Stresses: Plant Resistance Through Breeding and Molecular Approaches; Ashraf, M., Harris, P.J.C., Eds.; Howarth Press Inc.: New York, NY, USA, 2005. [Google Scholar]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Goyal, K.; Walton, L.J.; Tunnacliffe, A. LEA proteins prevent protein aggregation due to water stress. Biochem. J. 2005, 388, 151–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahid, A.; Close, T.J. Expression of dehydrins under heat stress and their relationship with water relations of sugarcane leaves. Biol. Plant. 2007, 51, 104–109. [Google Scholar] [CrossRef]
- Iba, K. Acclimative response to temperature stress in higher plants: Approaches of gene engineering for temperature tolerance. Annu. Rev. Plant Biol. 2002, 53, 225–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basha, E.; O’Neill, H.; Vierling, E. Small heat shock proteins and α-crystallins: Dynamic proteins with flexible functions. Trends Biochem. Sci. 2012, 37, 106–117. [Google Scholar] [CrossRef] [Green Version]
- Waters, E.R. The evolution, function, structure, and expression of the plant sHSPs. J. Exp. Bot. 2013, 64, 391–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondino, H.G.; Valle, E.M.; Ten Have, A. Evolution and functional diversification of the small heat shock protein/α-crystallin family in higher plants. Planta 2012, 235, 1299–1313. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Liu, Y.; Kong, X.; Zhang, D.; Pan, J.; Zhou, Y.; Wang, L.; Li, D.; Yang, X. ZmHSP16. 9, a cytosolic class I small heat shock protein in maize (Zea mays), confers heat tolerance in transgenic tobacco. Plant Cell Rep. 2012, 31, 1473–1484. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Sun, C.; Li, Z.; Hu, Q.; Han, L.; Luo, H. AsHSP17, a creeping bentgrass small heat shock protein modulates plant photosynthesis and ABA-dependent and independent signalling to attenuate plant response to abiotic stress. Plant Cell Environ. 2016, 39, 1320–1337. [Google Scholar] [CrossRef]
- Sun, Y.; MacRae, T.H. Small heat shock proteins: Molecular structure and chaperone function. Cell. Mol. Life Sci. 2005, 62, 2460–2476. [Google Scholar] [CrossRef] [PubMed]
- Santhanagopalan, I.; Basha, E.; Ballard, K.N.; Bopp, N.E.; Vierling, E. Model chaperones: Small heat shock proteins from plants. In The Big Book on Small Heat Shock Proteins; Tanguay, R., Hightower, L., Eds.; Springer: Cham, Switzerland, 2015; Volume 8, pp. 119–153. [Google Scholar] [CrossRef]
- Sun, W.; Van Montagu, M.; Verbruggen, N. Small heat shock proteins and stress tolerance in plants. Biochim. Biophys. Acta Gene Struct. Expr. 2002, 1577, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Siddique, M.; Gernhard, S.; von Koskull-Döring, P.; Vierling, E.; Scharf, K.D. The plant sHSP superfamily: Five new members in Arabidopsis thaliana with unexpected properties. Cell Stress and Chaperones 2008, 13, 183–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vierling, R.A.; Nguyen, H.T. Heat-shock protein gene expression in diploid wheat genotypes differing in thermal tolerance. Crop Sci. 1992, 32, 370–377. [Google Scholar] [CrossRef]
- Long, T.A. Many needles in a haystack: Cell-type specific abiotic stress responses. Curr. Opin. Plant Biol. 2011, 14, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Wang, Y.; Zhang, K.; Gao, C. Expression analysis of nine small heat shock protein genes from Tamarix hispida in response to different abiotic stresses and abscisic acid treatment. Mol. Biol. Rep. 2014, 41, 1279–1289. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, Z.; Jing, Y.; Wang, L.; Liu, X.; Liu, Y.; Deng, X. Ectopic over-expression of BhHsf1, a heat shock factor from the resurrection plant Boea hygrometrica, leads to increased thermotolerance and retarded growth in transgenic Arabidopsis and tobacco. Plant Mol. Biol. 2009, 71, 451. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, B.; Sun, D.; Deng, X. Molecular cloning and differential expression of sHSP gene family members from the resurrection plant Boea hygrometrica in response to abiotic stresses. Biologia 2013, 68, 651–661. [Google Scholar] [CrossRef]
- Gechev, T.S.; Benina, M.; Obata, T.; Tohge, T.; Sujeeth, N.; Minkov, I.; Hille, J.; Temanni, M.-R.; Marriott, A.S.; Bergström, E.; et al. Molecular mechanisms of desiccation tolerance in the resurrection glacial relic Haberlea rhodopensis. Cell Mol. Life Sci. 2013, 70, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, M.; Li, X.; Cao, B.; Ma, X. Identification of differentially expressed genes in leaf of Reaumuria soongorica under PEG-induced drought stress by digital gene expression profiling. PLoS ONE 2014, 9, e104142. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Moyankova, D.; Lin, C.T.; Mladenov, P.; Sun, R.Z.; Djilianov, D.; Deng, X. Transcriptome reprogramming during severe dehydration contributes to physiological and metabolic changes in the resurrection plant Haberlea rhodopensis. BMC Plant Biol. 2018, 18, 351. [Google Scholar] [CrossRef] [PubMed]
- Alamillo, J.; Almoguera, C.; Bartels, D.; Jordano, J. Constitutive expression of small heat shock proteins in vegetative tissues of the resurrection plant Craterostigma plantagineum. Plant Mol. Biol. 1995, 29, 1093–1099. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, U.; Schliwa, U.; Bilger, W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Tyystjärvi, E.; Karunen, J. A microcomputer program and fast analog to digital converter card for the analysis of fluorescence induction transients. Photosynth. Res. 1990, 26, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Rinderle, U. The role of chlorophyll fluorescence in the detection of stress conditions in plants. Crit. Rev. Anal. Chem. 1988, 19, S29–S85. [Google Scholar] [CrossRef]
- Van Kooten, O.; Snel, J.F. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosynth. Res. 1990, 25, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Okadjima, T.; Tanabe, T.; Yasuda, T. Nonurea sodium dodecyl sulfate-polyacrylamide gel electrophoresis with high-molarity buffers for the separation of protein and peptides. Anal. Biochem. 1993, 211, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Robertson, E.; Dannelly, H.; Malloy, P.; Reevs, H. Rapid isoelectric focusing in a vertical polyacrylamide minigel system. Anal. Biochem. 1987, 167, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, K.; Szigeti, Z.; Sarvari, E.; Gaspar, L.; Maslenkova, L.; Peeva, V.; Peli, E.; Tuba, Z. Photosynthetic activity of homoiochlorophyllous desiccation tolerant plant Haberlea rhodopensis during dehydration and rehydration. Planta 2007, 225, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Babani, F. Light adaptation and senescence of the photosynthetic apparatus. Changes in pigment composition, chlorophyll fluorescence parameters and photosynthetic activity. In Chlorophyll a Fluorescence. Advances in Photosynthesis and Respiration; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; Volume 19, pp. 713–736. [Google Scholar] [CrossRef]
- Niyogi, K.K. Safety valves for photosynthesis. Curr. Opin. Plant Biol. 2000, 3, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Krause, G.H.; Weis, E. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev. Plant Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Augusti, A.; Scartazza, A.; Navari-Izzo, F.; Sgherri, C.L.M.; Stevanovic, B.; Brugnoli, E. Photosystem II photochemical efficiency, zeaxanthin and antioxidant contents in the poikilohydric Ramonda serbica during dehydration and rehydration. Photosynth. Res. 2001, 67, 79–88. [Google Scholar] [CrossRef]
- Huang, W.; Yang, S.J.; Zhang, S.B.; Zhang, J.L.; Cao, K.F. Cyclic electron flow plays an important role in photoprotection for the resurrection plant Paraboea rufescens under drought stress. Planta 2012, 235, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Heckathorn, S.A.; Downs, C.A.; Sharkey, T.D.; Coleman, J.S. The small, methionine-rich chloroplast heat-shock protein protects photosystem II electron transport during heat stress. Plant Physiol. 1998, 116, 439–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preczewski, P.J.; Heckathorn, S.A.; Downs, C.A.; Coleman, J.S. Photosynthetic thermotolerance is quantitatively and positively correlated with production of specific heat-shock proteins among nine genotypes of Lycopersicon (tomato). Photosynthetica 2000, 38, 127–134. [Google Scholar] [CrossRef]
- Guo, L.; Chen, S.; Liu, K.; Liu, Y.; Ni, L.; Zhang, K.; Zhang, L. Isolation of heat shock factor HsfA1a-binding sites in vivo revealed variations of heat shock elements in Arabidopsis thaliana. Plant Cell Physiol. 2008, 49, 1306–1315. [Google Scholar] [CrossRef] [Green Version]
- Shakeel, S.; Haq, N.U.; Heckathorn, S.A.; Hamilton, E.W.; Luthe, D. Ecotypic variation in chloroplast small heat-shock proteins and related thermotolerance in Chenopodium album. Plant Physiol. Biochem. 2011, 49, 898–908. [Google Scholar] [CrossRef]
- Ledesma, N.A.; Kawabata, S.; Sugiyama, N. Effect of high temperature on protein expression in strawberry plants. Biol. Plant. 2004, 48, 73–79. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shang, H.; Liu, Y.; Zheng, M.; Wu, R.; Phillips, J.; Bartels, D.; Deng, X. A role for a cell wall localized glycine-rich protein in dehydration and rehydration of the resurrection plant Boea hygrometrica. Plant Biol. 2009, 11, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Röehrig, H.; Schmidt, J.; Colby, T.; Braeutigam, A.; Hufnagel, P.; Bartels, D. Desiccation of the resurrection plant Craterostigma plantagineum induces dynamic changes in protein phosphorylation. Plant Cell Environ. 2006, 29, 1606–1617. [Google Scholar] [CrossRef] [Green Version]
- Mladenov, P.; Zasheva, D.; Planchon, S.; Leclercq, C.C.; Falconet, D.; Moyet, L.; Brugière, S.; Moyankova, D.; Tchorbadjieva, M.; Ferro, M.; et al. Proteomics evidence of a systemic response to desiccation in the resurrection plant Haberlea rhodopensis. Int. J. Mol. Sci. 2022, 23, 8520. [Google Scholar] [CrossRef] [PubMed]
- Haq, N.U.; Raza, S.; Luthe, D.S.; Heckathorn, S.A.; Shakeel, S.N. A dual role for the chloroplast small heat shock protein of Chenopodium album including protection from both heat and metal stress. Plant Mol. Biol. Rep. 2013, 31, 398–408. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Mittler, R. The combined effect of drought stress and heat shock on gene expression in tobacco. Plant Physiol. 2002, 130, 1143–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Downs, C.A.; Ryan, S.L.; Heckathorn, S.A. The chloroplast small heat-shock protein: Evidence for a general role in protecting photosystem II against oxidative stress and photoinhibition. J. Plant Physiol. 1999, 155, 488–496. [Google Scholar] [CrossRef]
- Hu, X.; Li, Y.; Li, C.; Yang, H.; Wang, W.; Lu, M. Characterization of small heat shock proteins associated with maize tolerance to combined drought and heat stress. J. Plant Growth Regul. 2010, 29, 455–464. [Google Scholar] [CrossRef]
- Downs, C.A.; Coleman, J.S.; Heckathorn, S.A. The chloroplast 22-Ku heat-shock protein: A lumenal protein that associates with the oxygen evolving complex and protects photosystem II during heat stress. J. Plant Physiol. 1999, 155, 477–487. [Google Scholar] [CrossRef]
- Heckathorn, S.A.; Downs, C.A.; Coleman, J.S. Small heat shock proteins protect electron transport in chloroplasts and mitochondria during stress. Am. Zool. 1999, 39, 865–876. [Google Scholar] [CrossRef]
- Török, Z.; Goloubinoff, P.; Horváth, I.; Tsvetkova, N.M.; Glatz, A.; Balogh, G.; Varvasovszki, V.; Los, D.A.; Vierling, E.; Crowe, J.H.; et al. Synechocystis HSP17 is an amphitropic protein that stabilizes heat-stressed membranes and binds denatured proteins for subsequent chaperone-mediated refolding. Proc. Natl. Acad. Sci. USA 2001, 98, 3098–3103. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specificity | Protein, No. |
|---|---|
| Induced by dehydration (D) | 2, 9 |
| Induced by high temperature (H) | 3, 4, 5 |
| Induced by dehydration at 38 °C (D + H) | 1, 6, 7, 8, 10–14 |
| kDa | Protein, No. |
|---|---|
| 19.0 | 1, 2 |
| 18.5 | 3, 4, 5, 10 |
| 18.0 | 6, 7, 12, 13 |
| 17.5 | 14 |
| 17.0 | 8, 9 |
| 16.5 | 11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mihailova, G.; Tchorbadjieva, M.; Rakleova, G.; Georgieva, K. Differential Accumulation of sHSPs Isoforms during Desiccation of the Resurrection Plant Haberlea rhodopensis Friv. under Optimal and High Temperature. Life 2023, 13, 238. https://doi.org/10.3390/life13010238
Mihailova G, Tchorbadjieva M, Rakleova G, Georgieva K. Differential Accumulation of sHSPs Isoforms during Desiccation of the Resurrection Plant Haberlea rhodopensis Friv. under Optimal and High Temperature. Life. 2023; 13(1):238. https://doi.org/10.3390/life13010238
Chicago/Turabian StyleMihailova, Gergana, Magdalena Tchorbadjieva, Goritsa Rakleova, and Katya Georgieva. 2023. "Differential Accumulation of sHSPs Isoforms during Desiccation of the Resurrection Plant Haberlea rhodopensis Friv. under Optimal and High Temperature" Life 13, no. 1: 238. https://doi.org/10.3390/life13010238