

Probiotic Mechanisms Affecting Glucose Homeostasis: A Scoping Review

Abstract
:1. Introduction
2. Materials and Methods
Search Strategy and Studies’ Selection
3. Effect of Probiotics on Glucose Metabolism and Homeostasis
3.1. General Remarks about the Mechanisms of Probiotics
- The use of oversimplified in vitro models that often fail to reproduce the results in vivo;
- The use of “human” probiotics in animal models in vivo, (e.g., rodents), which do not take into account the functional significance of the species- and strain-specific administration of probiotics and their impact on the host’s immune response and its microbiota;
- Probiotics’ action ultimately requires the involvement of the endogenous microbiota, which is host-specific and often diet-dependent and therefore hardly reproducible;
- Most of the mechanisms of probiotics presented in research studies have suggested two main principles of probiotics’ action [41];
- Direct, contact-dependent principle (binding to different surface molecules);
- Indirect principle via secretory molecules (production of bioactive peptides and metabolites).
3.2. Effect of Probiotics on Blood Parameters
3.2.1. In Vivo Human Studies
3.2.2. In Vivo Animal Models Studies
3.2.3. In Vitro Studies
3.3. Effect of Probiotics on Brain Function
3.4. Effect of Probiotics on Bile Acid Metabolism
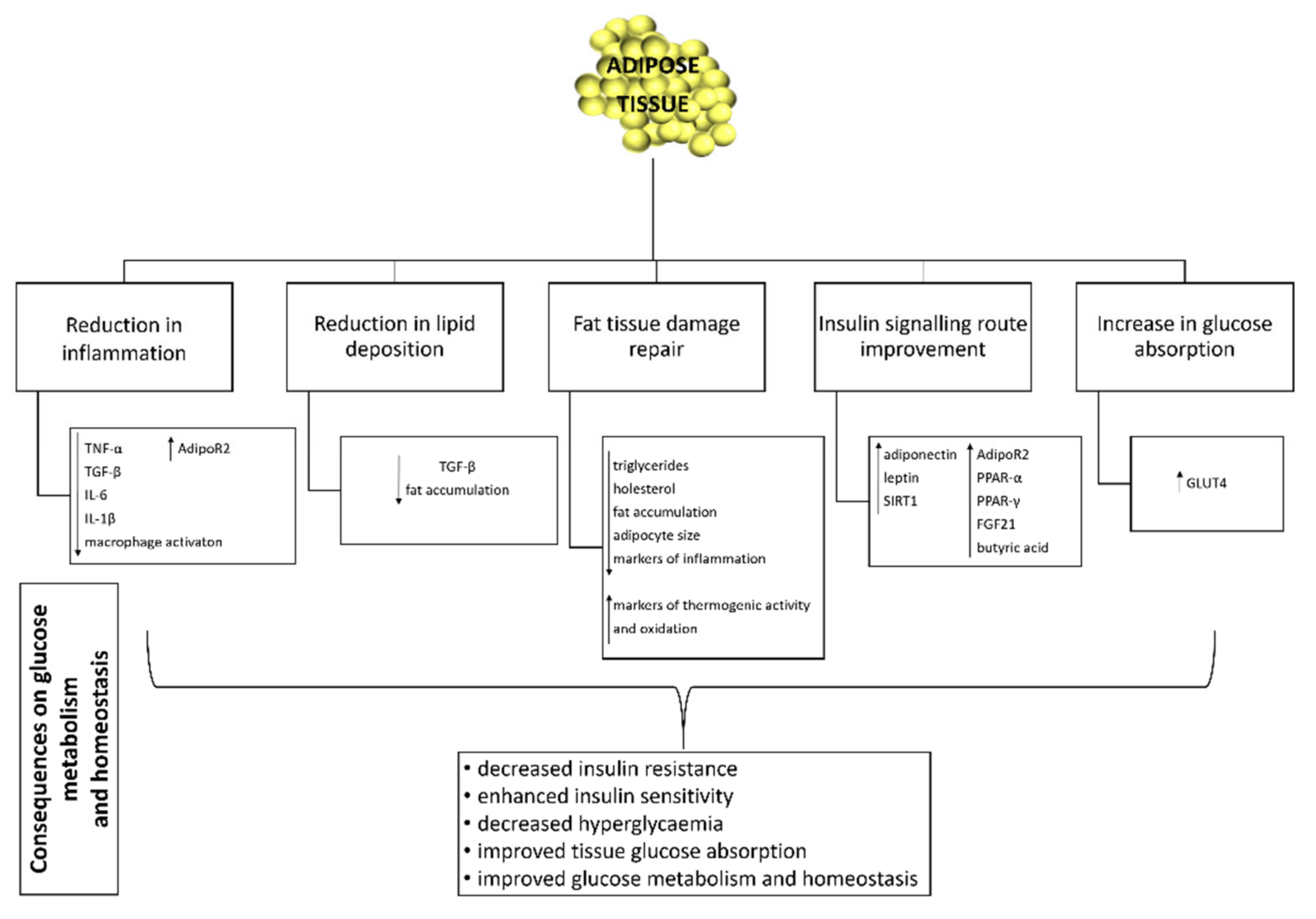
3.5. Effect of Probiotics on Adipose Tissue Function and Inflammation
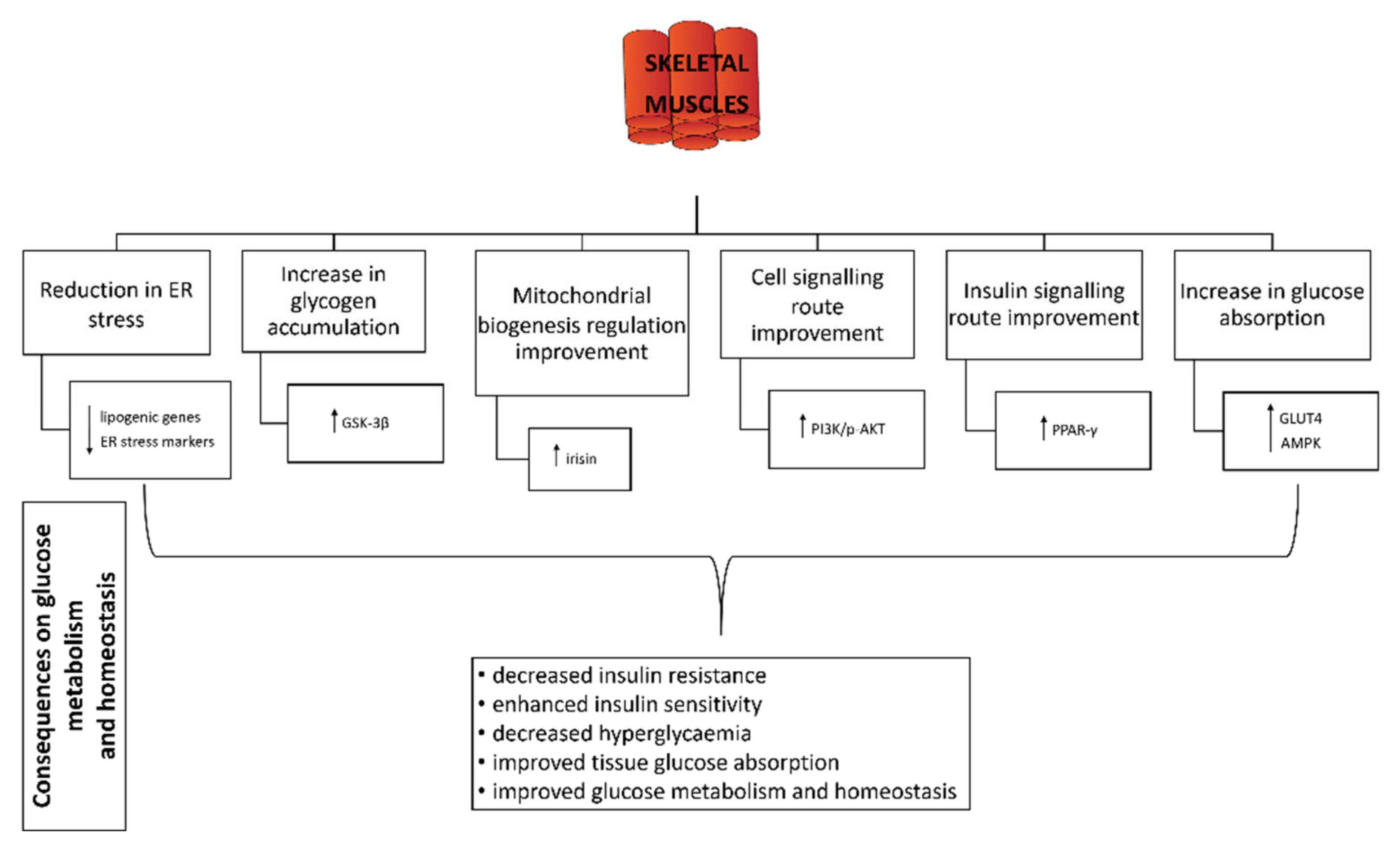
3.6. Effect of Probiotics on Skeletal Muscle
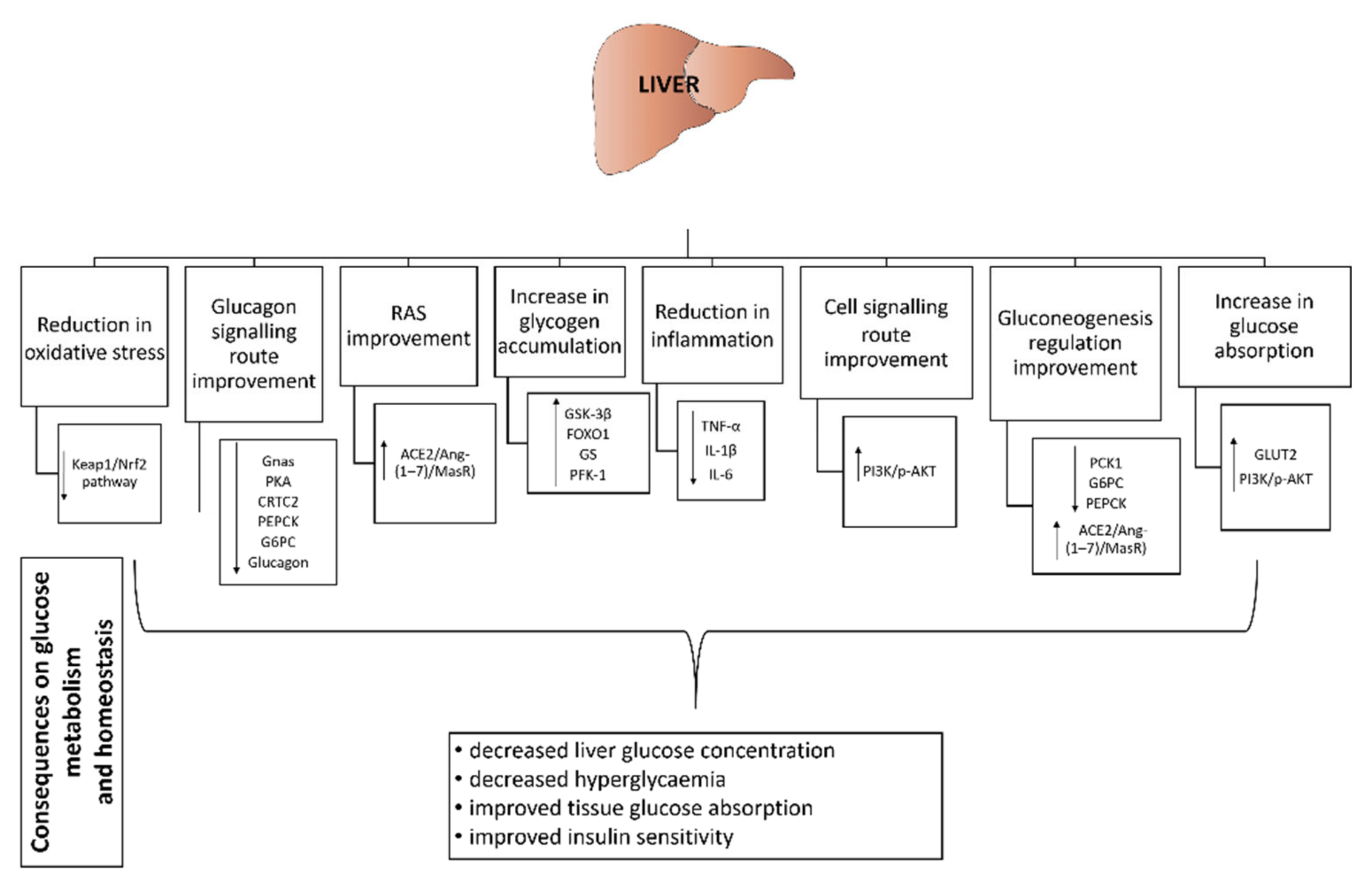
3.7. Effect of Probiotics on Liver, Pancreas and Kidney
3.7.1. Liver
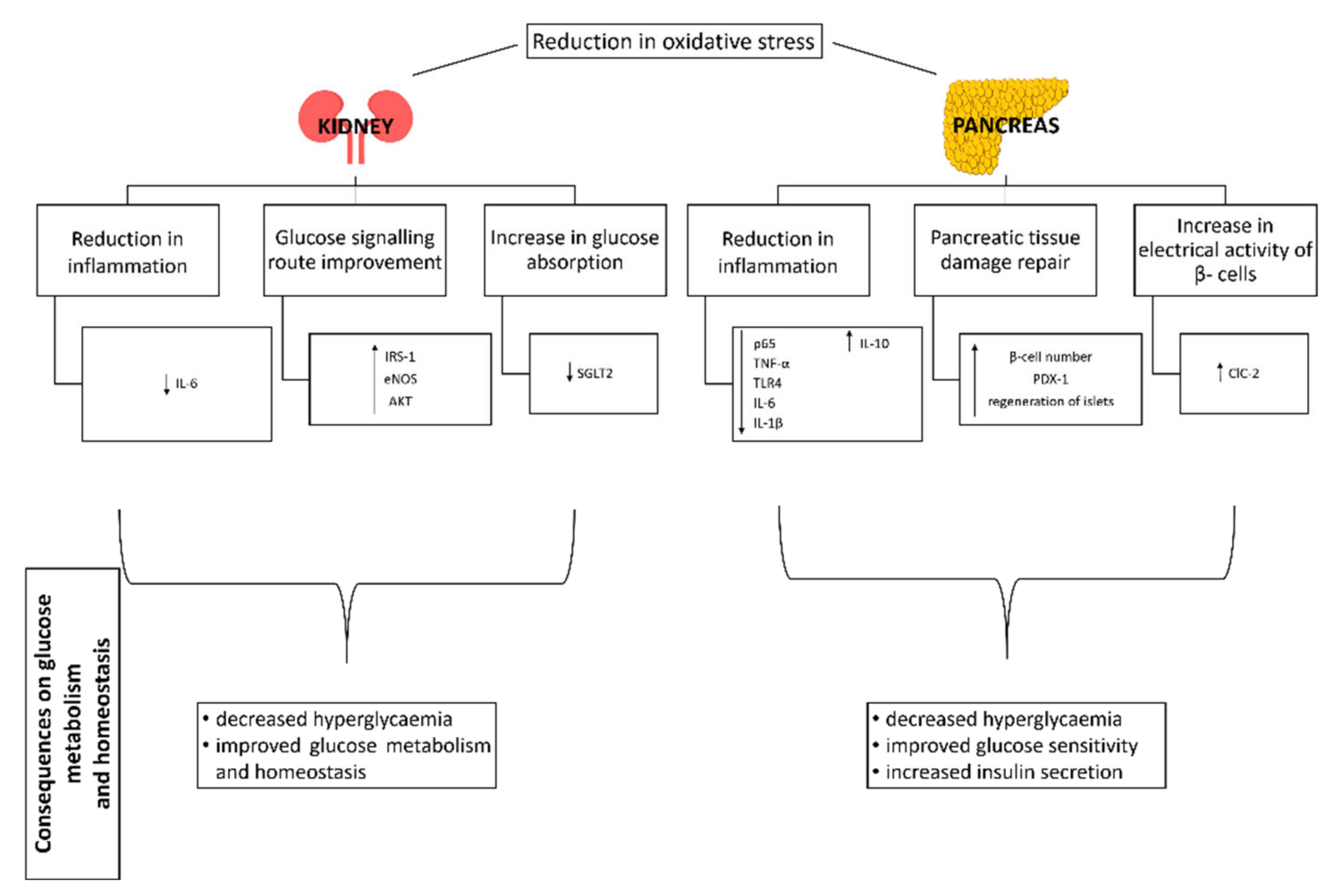
3.7.2. Pancreas
3.7.3. Kidney
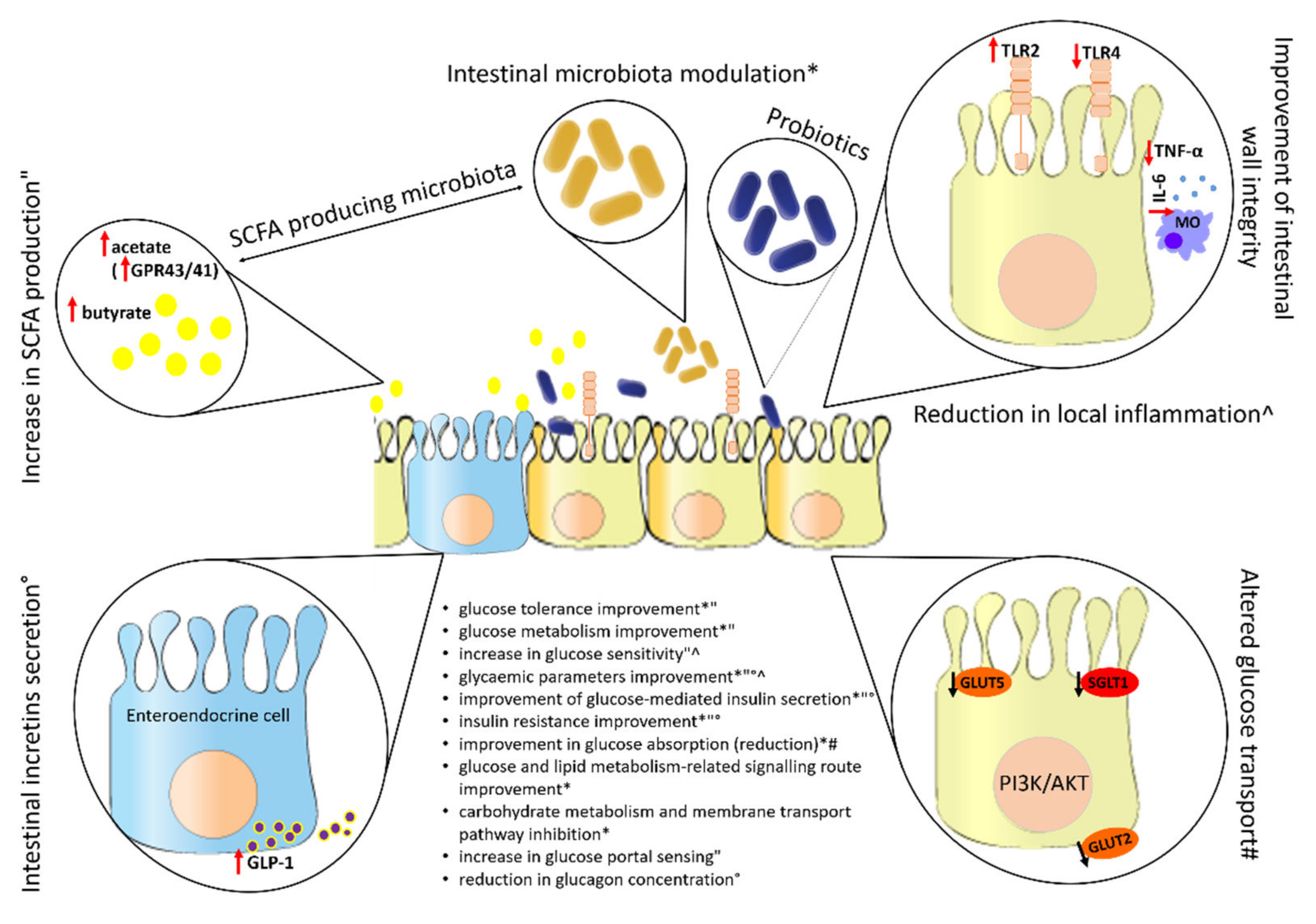
3.8. Effect of Probiotics on the Intestine
3.8.1. Effect of Probiotics on the Intestinal Microbiota
- In vivo human studies
- In vivo animal model studies
3.8.2. Effect of Probiotics on the Formation of SCFA
3.8.3. Effect of Probiotics on the Secretion of Intestinal Incretins
3.8.4. Effect of Probiotics on Permeability and Integrity of the Intestinal Wall
3.8.5. Effect of Probiotics on Glucose Transport in Intestine
4. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Harris, R.A. Carbohydrate metabolism I: Major metabolic pathways and their control. In Textbook of Biochemistry with Clinical Correlations, 4th ed.; Devlin, T.M., Ed.; Wiley, Inc.: New York, NY, USA, 1997; pp. 267–335. [Google Scholar]
- Simon, K.; Wittmann, I. Can blood glucose value really be referred to as a metabolic parameter? Rev. Endocr. Metab. Disord. 2019, 20, 151–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Park, S.Y.; Choi, C.S. Insulin Resistance: From Mechanisms to Therapeutic Strategies. Diabetes Metab. J. 2022, 46, 15–37. [Google Scholar] [CrossRef] [PubMed]
- Rohm, T.V.; Meier, D.T.; Olefsky, J.M.; Donath, M.Y. Inflammation in obesity, diabetes, and related disorders. Immunity 2022, 55, 31–55. [Google Scholar] [CrossRef] [PubMed]
- Lima, J.E.B.F.; Moreira, N.C.S.; Sakamoto-Hojo, E.T. Mechanisms underlying the pathophysiology of type 2 diabetes: From risk factors to oxidative stress, metabolic dysfunction, and hyperglycemia. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2022, 874–875, 503437. [Google Scholar] [CrossRef] [PubMed]
- Ceriello, A. Postprandial Hyperglycemia and Diabetes Complications: Is It Time to Treat? Diabetes 2005, 54, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouri, M.; Badireddy, M. Hyperglycemia; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Giri, B.; Dey, S.; Das, T.; Sarkar, M.; Banerjee, J.; Dash, S.K. Chronic hyperglycemia mediated physiological alteration and metabolic distortion leads to organ dysfunction, infection, cancer progression and other pathophysiological consequences: An update on glucose toxicity. Biomed. Pharmacother. 2018, 107, 306–328. [Google Scholar] [CrossRef]
- Nishida, C.; Uauy, R.; Kumanyika, S.; Shetty, P. The joint WHO/FAO expert consultation on diet, nutrition and the prevention of chronic diseases: Process, product and policy implications. Public Health Nutr. 2004, 7, 245–250. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Diet, Nutrition, and the Prevention of Chronic Diseases: Report of a WHO Study Group; World Health Organization: Geneva, Switzerland, 1990; ISBN 9241207973. [Google Scholar]
- Tuomilehto, J.; Lindström, J.; Eriksson, J.G.; Valle, T.T.; Hämäläinen, H.; Ilanne-Parikka, P.; Keinänen-Kiukaanniemi, S.; Laakso, M.; Louheranta, A.; Rastas, M.; et al. Prevention of Type 2 Diabetes Mellitus by Changes in Lifestyle among Subjects with Impaired Glucose Tolerance. N. Engl. J. Med. 2001, 344, 1343–1350. [Google Scholar] [CrossRef]
- Andrade, E.F.; de Oliveira Silva, V.; Orlando, D.R.; Pereira, L.J. Mechanisms Involved in Glycemic Control Promoted by Exercise in Diabetics. Curr. Diabetes Rev. 2019, 15, 105–110. [Google Scholar] [CrossRef]
- Gérard, C.; Vidal, H. Impact of Gut Microbiota on Host Glycemic Control. Front. Endocrinol. 2019, 10, 29. [Google Scholar] [CrossRef]
- Sivamaruthi, B.S.; Kesika, P.; Suganthy, N.; Chaiyasut, C. A Review on Role of Microbiome in Obesity and Antiobesity Properties of Probiotic Supplements. Biomed Res. Int. 2019, 2019, 3291367. [Google Scholar] [CrossRef]
- Tang, C.; Kong, L.; Shan, M.; Lu, Z.; Lu, Y. Protective and ameliorating effects of probiotics against diet-induced obesity: A review. Food Res. Int. 2021, 147, 110490. [Google Scholar] [CrossRef]
- Torres, S.; Fabersani, E.; Marquez, A.; Gauffin-Cano, P. Adipose tissue inflammation and metabolic syndrome. The proactive role of probiotics. Eur. J. Nutr. 2019, 58, 27–43. [Google Scholar] [CrossRef]
- He, M.; Shi, B. Gut microbiota as a potential target of metabolic syndrome: The role of probiotics and prebiotics. Cell Biosci. 2017, 7, 54. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Liu, J.; Xia, Y.; Ai, L. Probiotics-based interventions for diabetes mellitus: A review. Food Biosci. 2021, 43, 101172. [Google Scholar] [CrossRef]
- Sun, Z.; Sun, X.; Li, J.; Li, Z.; Hu, Q.; Li, L.; Hao, X.; Song, M.; Li, C. Using probiotics for type 2 diabetes mellitus intervention: Advances, questions, and potential. Crit. Rev. Food Sci. Nutr. 2019, 60, 670–683. [Google Scholar] [CrossRef]
- Zhai, L.; Wu, J.; Lam, Y.Y.; Kwan, H.Y.; Bian, Z.X.; Wong, H.L.X. Gut-Microbial Metabolites, Probiotics and Their Roles in Type 2 Diabetes. Int. J. Mol. Sci. 2021, 22, 12846. [Google Scholar] [CrossRef]
- Iatcu, C.O.; Steen, A.; Covasa, M. Gut Microbiota and Complications of Type-2 Diabetes. Nutrients 2021, 14, 166. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [Green Version]
- Salari, A.; Mahdavi-Roshan, M.; Kheirkhah, J.; Ghorbani, Z. Probiotics supplementation and cardiometabolic risk factors: A new insight into recent advances, potential mechanisms, and clinical implications. PharmaNutrition 2021, 16, 100261. [Google Scholar] [CrossRef]
- Kim, Y.A.; Keogh, J.B.; Clifton, P.M. Probiotics, prebiotics, synbiotics and insulin sensitivity. Nutr. Res. Rev. 2018, 31, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Miglioranza Scavuzzi, B.; Miglioranza, L.H.d.S.; Henrique, F.C.; Pitelli Paroschi, T.; Lozovoy, M.A.B.; Simão, A.N.C.; Dichi, I. The role of probiotics on each component of the metabolic syndrome and other cardiovascular risks. Expert Opin. Ther. Targets 2015, 19, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Razmpoosh, E.; Javadi, M.; Ejtahed, H.-S.; Mirmiran, P. Probiotics as beneficial agents in the management of diabetes mellitus: A systematic review. Diabetes. Metab. Res. Rev. 2016, 32, 143–168. [Google Scholar] [CrossRef] [PubMed]
- Bezirtzoglou, E.; Stavropoulou, E.; Kantartzi, K.; Tsigalou, C.; Voidarou, C.; Mitropoulou, G.; Prapa, I.; Santarmaki, V.; Kompoura, V.; Yanni, A.E.; et al. Maintaining Digestive Health in Diabetes: The Role of the Gut Microbiome and the Challenge of Functional Foods. Microorganisms 2021, 9, 516. [Google Scholar] [CrossRef]
- Papadopoulos, P.D.; Tsigalou, C.; Valsamaki, P.N.; Konstantinidis, T.G.; Voidarou, C.; Bezirtzoglou, E. The Emerging Role of the Gut Microbiome in Cardiovascular Disease: Current Knowledge and Perspectives. Biomedicines 2022, 10, 948. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- FAO/WHO Guidelines for the Evaluation of Probiotics in Food. Report of a Joint FAO/WHO Working Group on Drafting Guidelines for the Evaluation of Probiotics in Food. Available online: http://www.who.int/foodsafety/fs_management/en/probiotic_guidelines.pdf?ua=1 (accessed on 23 May 2019).
- EFSA Opinion of the Scientific Committee on a request from EFSA related to a generic approach to the safety assessment by EFSA of microorganisms used in food/feed and the production of food/feed additives. EFSA J. 2005, 226, 1–12.
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Voidarou, C.; Antoniadou, M.; Rozos, G.; Tzora, A.; Skoufos, I.; Varzakas, T.; Lagiou, A.; Bezirtzoglou, E. Fermentative Foods: Microbiology, Biochemistry, Potential Human Health Benefits and Public Health Issues. Foods 2020, 10, 69. [Google Scholar] [CrossRef]
- Hampe, C.S.; Roth, C.L. Probiotic strains and mechanistic insights for the treatment of type 2 diabetes. Endocrine 2017, 58, 207–227. [Google Scholar] [CrossRef]
- George Kerry, R.; Patra, J.K.; Gouda, S.; Park, Y.; Shin, H.-S.; Das, G. Benefaction of probiotics for human health: A review. J. Food Drug Anal. 2018, 26, 927–939. [Google Scholar] [CrossRef] [Green Version]
- De Almada, C.N.; Almada, C.N.; Martinez, R.C.R.; Sant’Ana, A.S. Paraprobiotics: Evidences on their ability to modify biological responses, inactivation methods and perspectives on their application in foods. Trends Food Sci. Technol. 2016, 58, 96–114. [Google Scholar] [CrossRef]
- Turpin, W.; Humblot, C.; Thomas, M.; Guyot, J.-P. Lactobacilli as multifaceted probiotics with poorly disclosed molecular mechanisms. Int. J. Food Microbiol. 2010, 143, 87–102. [Google Scholar] [CrossRef]
- Reid, G. Probiotics: Definition, scope and mechanisms of action. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 17–25. [Google Scholar] [CrossRef]
- Peterson, C.T.; Sharma, V.; Elmén, L.; Peterson, S.N. Immune homeostasis, dysbiosis and therapeutic modulation of the gut microbiota. Clin. Exp. Immunol. 2015, 179, 363–377. [Google Scholar] [CrossRef] [Green Version]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of Action of Probiotics. Adv. Nutr. 2019, 10, S49–S66. [Google Scholar] [CrossRef] [Green Version]
- Suez, J.; Zmora, N.; Segal, E.; Elinav, E. The pros, cons, and many unknowns of probiotics. Nat. Med. 2019, 25, 716–729. [Google Scholar] [CrossRef]
- Barros, C.P.; Grom, L.C.; Guimarães, J.T.; Balthazar, C.F.; Rocha, R.S.; Silva, R.; Almada, C.N.; Pimentel, T.C.; Venâncio, E.L.; Collopy Junior, I.; et al. Paraprobiotic obtained by ohmic heating added in whey-grape juice drink is effective to control postprandial glycemia in healthy adults. Food Res. Int. 2021, 140, 109905. [Google Scholar] [CrossRef]
- Ejtahed, H.S.; Mohtadi-Nia, J.; Homayouni-Rad, A.; Niafar, M.; Asghari-Jafarabadi, M.; Mofid, V. Probiotic yogurt improves antioxidant status in type 2 diabetic patients. Nutrition 2012, 28, 539–543. [Google Scholar] [CrossRef]
- Rezazadeh, L.; Gargari, B.P.; Jafarabadi, M.A.; Alipour, B. Effects of probiotic yogurt on glycemic indexes and endothelial dysfunction markers in patients with metabolic syndrome. Nutrition 2019, 62, 162–168. [Google Scholar] [CrossRef]
- Schellekens, H.; Torres-Fuentes, C.; van de Wouw, M.; Long-Smith, C.M.; Mitchell, A.; Strain, C.; Berding, K.; Bastiaanssen, T.F.S.; Rea, K.; Golubeva, A.V.; et al. Bifidobacterium longum counters the effects of obesity: Partial successful translation from rodent to human. EBioMedicine 2021, 63, 103176. [Google Scholar] [CrossRef]
- Toshimitsu, T.; Gotou, A.; Furuichi, K.; Hachimura, S.; Asami, Y. Effects of 12-wk Lactobacillus plantarum OLL2712 treatment on glucose metabolism and chronic inflammation in prediabetic individuals: A single-arm pilot study. Nutrition 2019, 58, 175–180. [Google Scholar] [CrossRef]
- Sabico, S.; Al-Mashharawi, A.; Al-Daghri, N.M.; Wani, K.; Amer, O.E.; Hussain, D.S.; Ahmed Ansari, M.G.; Masoud, M.S.; Alokail, M.S.; McTernan, P.G. Effects of a 6-month multi-strain probiotics supplementation in endotoxemic, inflammatory and cardiometabolic status of T2DM patients: A randomized, double-blind, placebo-controlled trial. Clin. Nutr. 2019, 38, 1561–1569. [Google Scholar] [CrossRef] [Green Version]
- Khalili, L.; Alipour, B.; Jafar-Abadi, M.A.; Faraji, I.; Hassanalilou, T.; Abbasi, M.M.; Vaghef-Mehrabany, E.; Sani, M.A. The Effects of Lactobacillus casei on Glycemic Response, Serum Sirtuin1 and Fetuin-A Levels in Patients with Type 2 Diabetes Mellitus: A Randomized Controlled Trial. Iran. Biomed. J. 2019, 23, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Khalili, L.; Alipour, B.; Asghari Jafarabadi, M.; Hassanalilou, T.; Mesgari Abbasi, M.; Faraji, I. Probiotic assisted weight management as a main factor for glycemic control in patients with type 2 diabetes: A randomized controlled trial. Diabetol. Metab. Syndr. 2019, 11, 5. [Google Scholar] [CrossRef]
- Tonucci, L.B.; Olbrich dos Santos, K.M.; Licursi de Oliveira, L.; Rocha Ribeiro, S.M.; Duarte Martino, H.S. Clinical application of probiotics in type 2 diabetes mellitus: A randomized, double-blind, placebo-controlled study. Clin. Nutr. 2017, 36, 85–92. [Google Scholar] [CrossRef]
- Hsieh, M.C.; Tsai, W.H.; Jheng, Y.P.; Su, S.L.; Wang, S.Y.; Lin, C.C.; Chen, Y.H.; Chang, W.W. The beneficial effects of Lactobacillus reuteri ADR-1 or ADR-3 consumption on type 2 diabetes mellitus: A randomized, double-blinded, placebo-controlled trial. Sci. Rep. 2018, 8, 16791. [Google Scholar] [CrossRef]
- Kobyliak, N.; Falalyeyeva, T.; Mykhalchyshyn, G.; Kyriienko, D.; Komissarenko, I. Effect of alive probiotic on insulin resistance in type 2 diabetes patients: Randomized clinical trial. Diabetes Metab. Syndr. Clin. Res. Rev. 2018, 12, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Mohamadshahi, M.; Veissi, M.; Haidari, F.; Javid, A.Z.; Mohammadi, F.; Shirbeigi, E. Effects of probiotic yogurt consumption on lipid profile in type 2 diabetic patients: A randomized controlled clinical trial. J. Res. Med. Sci. 2014, 19, 531–536. [Google Scholar]
- Mobini, R.; Tremaroli, V.; Ståhlman, M.; Karlsson, F.; Levin, M.; Ljungberg, M.; Sohlin, M.; Bertéus Forslund, H.; Perkins, R.; Bäckhed, F.; et al. Metabolic effects of Lactobacillus reuteri DSM 17938 in people with type 2 diabetes: A randomized controlled trial. Diabetes Obes. Metab. 2017, 19, 579–589. [Google Scholar] [CrossRef]
- Andreasen, A.S.; Larsen, N.; Pedersen-Skovsgaard, T.; Berg, R.M.G.; Møller, K.; Svendsen, K.D.; Jakobsen, M.; Pedersen, B.K. Effects of Lactobacillus acidophilus NCFM on insulin sensitivity and the systemic inflammatory response in human subjects. Br. J. Nutr. 2010, 104, 1831–1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulston, C.J.; Churnside, A.A.; Venables, M.C. Probiotic supplementation prevents high-fat, overfeeding-induced insulin resistance in human subjects. Br. J. Nutr. 2015, 113, 596–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, C.M.C.; Gibson, G.R.; Rowland, I. Health benefits of probiotics: Are mixtures more effective than single strains? Eur. J. Nutr. 2011, 50, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kurl, S.; Zaccardi, F.; Onaemo, V.N.; Jae, S.Y.; Kauhanen, J.; Ronkainen, K.; Laukkanen, J.A. Association between HOMA-IR, fasting insulin and fasting glucose with coronary heart disease mortality in nondiabetic men: A 20-year observational study. Acta Diabetol. 2015, 52, 183–186. [Google Scholar] [CrossRef]
- Holmes, D. Physiologic role of IL-1β in glucose homeostasis. Nat. Rev. Endocrinol. 2017, 13, 128. [Google Scholar] [CrossRef]
- Ibfelt, T.; Fischer, C.P.; Plomgaard, P.; Van Hall, G.; Pedersen, B.K. The acute effects of low-dose TNF- on glucose metabolism and β -cell function in humans. Mediators Inflamm. 2014, 2014, 295478. [Google Scholar] [CrossRef]
- Gonzalez, Y.; Herrera, M.T.; Soldevila, G.; Garcia-Garcia, L.; Fabián, G.; Pérez-Armendariz, E.M.; Bobadilla, K.; Guzmán-Beltrán, S.; Sada, E.; Torres, M. High glucose concentrations induce TNF-α production through the down-regulation of CD33 in primary human monocytes. BMC Immunol. 2012, 13, 19. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, P.-S.; An, Y.; Tsai, Y.-C.; Chen, Y.-C.; Chuang, C.-J.; Zeng, C.-T.; Wang, C.-T.; An-Erl King, V. Potential of probiotic strains to modulate the inflammatory responses of epithelial and immune cells in vitro. New Microbiol. 2013, 36, 167–179. [Google Scholar]
- Asemi, Z.; Zare, Z.; Shakeri, H.; Sabihi, S.; Esmaillzadeh, A. Effect of Multispecies Probiotic Supplements on Metabolic Profiles, hs-CRP, and Oxidative Stress in Patients with Type 2 Diabetes. Ann. Nutr. Metab. 2013, 63, 1–9. [Google Scholar] [CrossRef]
- Ismail, N.A.; Ragab, S.; Abd El Dayem, S.M.; Abd ElBaky, A.; Salah, N.; Hamed, M.; Assal, H.; Koura, H. Fetuin-A levels in obesity: Differences in relation to metabolic syndrome and correlation with clinical and laboratory variables. Arch. Med. Sci. 2012, 8, 826–833. [Google Scholar] [CrossRef]
- Srinivas, P.R.; Wagner, A.S.; Reddy, L.V.; Deutsch, D.D.; Leon, M.A.; Goustin, A.S.; Grunberger, G. Serum alpha 2-HS-glycoprotein is an inhibitor of the human insulin receptor at the tyrosine kinase level. Mol. Endocrinol. 1993, 7, 1445–1455. [Google Scholar] [CrossRef] [Green Version]
- Turkmen, K.; Karagoz, A.; Kucuk, A. Sirtuins as novel players in the pathogenesis of diabetes mellitus. World J. Diabetes 2014, 5, 894. [Google Scholar] [CrossRef]
- Sun, J.; Buys, N.J. Glucose- and glycaemic factor-lowering effects of probiotics on diabetes: A meta-analysis of randomised placebo-controlled trials. Br. J. Nutr. 2016, 115, 1167–1177. [Google Scholar] [CrossRef]
- Zhang, Q.; Wu, Y.; Fei, X. Effect of probiotics on glucose metabolism in patients with type 2 diabetes mellitus: A meta-analysis of randomized controlled trials. Medicina 2016, 52, 28–34. [Google Scholar] [CrossRef]
- Samah, S.; Ramasamy, K.; Lim, S.M.; Neoh, C.F. Probiotics for the management of type 2 diabetes mellitus: A systematic review and meta-analysis. Diabetes Res. Clin. Pract. 2016, 118, 172–182. [Google Scholar] [CrossRef]
- Nikbakht, E.; Khalesi, S.; Singh, I.; Williams, L.T.; West, N.P.; Colson, N. Effect of probiotics and synbiotics on blood glucose: A systematic review and meta-analysis of controlled trials. Eur. J. Nutr. 2018, 57, 95–106. [Google Scholar] [CrossRef]
- Ruan, Y.; Sun, J.; He, J.; Chen, F.; Chen, R.; Chen, H. Effect of Probiotics on Glycemic Control: A Systematic Review and Meta-Analysis of Randomized, Controlled Trials. PLoS ONE 2015, 10, e0132121. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Li, X.; Han, H.; Cui, H.; Peng, M.; Wang, G.; Wang, Z. Effect of probiotics on metabolic profiles in type 2 diabetes mellitus. Medicine 2016, 95, e4088. [Google Scholar] [CrossRef]
- He, J.; Zhang, F.; Han, Y. Effect of probiotics on lipid profiles and blood pressure in patients with type 2 diabetes: A meta-analysis of RCTs. Medicine 2017, 96, e9166. [Google Scholar] [CrossRef]
- Kasińska, M.A.; Drzewoski, J. Effectiveness of probiotics in type 2 diabetes: A meta-analysis. Pol. Arch. Med. Wewn. 2015, 125, 803–813. [Google Scholar] [CrossRef] [Green Version]
- Ardeshirlarijani, E.; Tabatabaei-Malazy, O.; Mohseni, S.; Qorbani, M.; Larijani, B.; Baradar Jalili, R. Effect of probiotics supplementation on glucose and oxidative stress in type 2 diabetes mellitus: A meta-analysis of randomized trials. DARU J. Pharm. Sci. 2019, 27, 827–837. [Google Scholar] [CrossRef]
- Tao, Y.-W.; Gu, Y.-L.; Mao, X.-Q.; Zhang, L.; Pei, Y.-F. Effects of probiotics on type II diabetes mellitus: A meta-analysis. J. Transl. Med. 2020, 18, 30. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Si, Q.; Yang, S.; Jiao, T.; Zhu, H.; Tian, P.; Wang, L.; Li, X.; Gong, L.; Zhao, J.; et al. Lactic acid bacteria reduce diabetes symptoms in mice by alleviating gut microbiota dysbiosis and inflammation in different manners. Food Funct. 2020, 11, 5898–5914. [Google Scholar] [CrossRef]
- Li, X.; Wang, N.; Yin, B.; Fang, D.; Jiang, T.; Fang, S.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Effects of Lactobacillus plantarum CCFM0236 on hyperglycaemia and insulin resistance in high-fat and streptozotocin-induced type 2 diabetic mice. J. Appl. Microbiol. 2016, 121, 1727–1736. [Google Scholar] [CrossRef]
- Sharma, P.; Bhardwaj, P.; Singh, R. Administration of Lactobacillus casei and Bifidobacterium bifidum Ameliorated Hyperglycemia, Dyslipidemia, and Oxidative Stress in Diabetic Rats. Int. J. Prev. Med. 2016, 7, 102. [Google Scholar] [CrossRef]
- Bejar, W.; Hamden, K.; Ben Salah, R.; Chouayekh, H. Lactobacillus plantarum TN627 significantly reduces complications of alloxan-induced diabetes in rats. Anaerobe 2013, 24, 4–11. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, S.; Zeng, Z.; Qin, Y.; Shen, Q.; Li, P. Anti-diabetic effects of Bifidobacterium animalis 01 through improving hepatic insulin sensitivity in type 2 diabetic rat model. J. Funct. Foods 2020, 67, 103843. [Google Scholar] [CrossRef]
- Toejing, P.; Khat-Udomkiri, N.; Intakhad, J.; Sirilun, S.; Chaiyasut, C.; Lailerd, N. Putative Mechanisms Responsible for the Antihyperglycemic Action of Lactobacillus paracasei HII01 in Experimental Type 2 Diabetic Rats. Nutrients 2020, 12, 3015. [Google Scholar] [CrossRef]
- Farida, E.; Nuraida, L.; Giriwono, P.E.; Jenie, B.S.L. Lactobacillus rhamnosus Reduces Blood Glucose Level through Downregulation of Gluconeogenesis Gene Expression in Streptozotocin-Induced Diabetic Rats. Int. J. Food Sci. 2020, 2020, 6108575. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Wang, F.; Zhao, P.; Zhang, R.; Zeng, Q. Effect of heat-killed Streptococcus thermophilus on type 2 diabetes rats. PeerJ 2019, 7, e7117. [Google Scholar] [CrossRef] [Green Version]
- Micioni Di Bonaventura, M.V.; Coman, M.M.; Tomassoni, D.; Di Bonaventura, E.M.; Botticelli, L.; Gabrielli, M.G.; Rossolini, G.M.; Di Pilato, V.; Cecchini, C.; Amedei, A.; et al. Supplementation with Lactiplantibacillus plantarum IMC 510 Modifies Microbiota Composition and Prevents Body Weight Gain Induced by Cafeteria Diet in Rats. Int. J. Mol. Sci. 2021, 22, 11171. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Tomar, S.K.; Thakur, K.; Singh, A.K. The ameliorative effects of probiotic Lactobacillus fermentum strain RS-2 on alloxan induced diabetic rats. J. Funct. Foods 2017, 28, 275–284. [Google Scholar] [CrossRef]
- Xiaojie, L.; Yue, J.; Xiaofei, Z.; Naisheng, Z.; Jiandong, T.; Yongguo, C. Bacillus licheniformis Zhengchangsheng® Inhibits Obesity by Regulating the AMP-Activated Protein Kinase Signaling Pathway. Probiotics Antimicrob. Proteins 2021, 13, 1658–1667. [Google Scholar] [CrossRef]
- Choi, S.-I.; You, S.; Kim, S.; Won, G.; Kang, C.-H.; Kim, G.-H. Weissella cibaria MG5285 and Lactobacillus reuteri MG5149 attenuated fat accumulation in adipose and hepatic steatosis in high-fat diet-induced C57BL/6J obese mice. Food Nutr. Res. 2021, 65, 8087. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, P.S.; Ho, H.-H.; Tsao, S.P.; Hsieh, S.-H.; Lin, W.-Y.; Chen, J.-F.; Kuo, Y.-W.; Tsai, S.-Y.; Huang, H.-Y. Multi-strain probiotic supplement attenuates streptozotocin-induced type-2 diabetes by reducing inflammation and β-cell death in rats. PLoS ONE 2021, 16, e0251646. [Google Scholar] [CrossRef] [PubMed]
- Okyere, S.K.; Xie, L.; Wen, J.; Ran, Y.; Ren, Z.; Deng, J.; Hu, Y. Bacillus toyonensis SAU-19 Ameliorates Hepatic Insulin Resistance in High-Fat Diet/Streptozocin-Induced Diabetic Mice. Nutritions 2021, 13, 4512. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, T.; Li, W.; Zhao, Y.; Long, H.; Liu, R.; Sui, W.; Zhang, M. Lactobacillus casei LC89 exerts antidiabetic effects through regulating hepatic glucagon response and gut microbiota in type 2 diabetic mice. Food Funct. 2021, 12, 8288–8299. [Google Scholar] [CrossRef]
- Holowacz, S.; Guigné, C.; Chêne, G.; Mouysset, S.; Guilbot, A.; Seyrig, C.; Dubourdeau, M. A multispecies Lactobacillus- and Bifidobacterium-containing probiotic mixture attenuates body weight gain and insulin resistance after a short-term challenge with a high-fat diet in C57/BL6J mice. PharmaNutrition 2015, 3, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, P.-S.; Ho, H.-H.; Hsieh, S.-H.; Kuo, Y.-W.; Tseng, H.-Y.; Kao, H.-F.; Wang, J.-Y. Lactobacillus salivarius AP-32 and Lactobacillus reuteri GL-104 decrease glycemic levels and attenuate diabetes-mediated liver and kidney injury in db/db mice. BMJ Open Diabetes Res. Care 2020, 8, e001028. [Google Scholar] [CrossRef] [Green Version]
- Zhong, H.; Zhang, Y.; Zhao, M.; Zhang, J.; Zhang, H.; Xi, Y.; Cai, H.; Feng, F. Screening of novel potential antidiabetic Lactobacillus plantarum strains based on in vitro and in vivo investigations. LWT 2021, 139, 110526. [Google Scholar] [CrossRef]
- Ben Othman, M.; Sakamoto, K. Effect of inactivated Bifidobacterium longum intake on obese diabetes model mice (TSOD). Food Res. Int. 2020, 129, 108792. [Google Scholar] [CrossRef]
- Wang, L.; Shang, Q.; Guo, W.; Wu, X.; Wu, L.; Wu, L.; Chen, T. Evaluation of the hypoglycemic effect of probiotics via directly consuming glucose in intestines of STZ-induced diabetic mice and glucose water-induced diabetic mice. J. Funct. Foods 2020, 64, 103614. [Google Scholar] [CrossRef]
- Andersson, U.; Bränning, C.; Ahrné, S.; Molin, G.; Alenfall, J.; Önning, G.; Nyman, M.; Holm, C. Probiotics lower plasma glucose in the high-fat fed C57BL/6J mouse. Benef. Microbes 2010, 1, 189–196. [Google Scholar] [CrossRef]
- Sakai, T.; Taki, T.; Nakamoto, A.; Shuto, E.; Tsutsumi, R.; Toshimitsu, T.; Makino, S.; Ikegami, S. Lactobacillus plantarum OLL2712 regulates glucose metabolism in C57BL/6 mice fed a high-fat diet. J. Nutr. Sci. Vitaminol. 2013, 59, 144–147. [Google Scholar] [CrossRef] [Green Version]
- Naito, E.; Yoshida, Y.; Makino, K.; Kounoshi, Y.; Kunihiro, S.; Takahashi, R.; Matsuzaki, T.; Miyazaki, K.; Ishikawa, F. Beneficial effect of oral administration of Lactobacillus casei strain Shirota on insulin resistance in diet-induced obesity mice. J. Appl. Microbiol. 2011, 110, 650–657. [Google Scholar] [CrossRef]
- Wang, Y.; Dilidaxi, D.; Wu, Y.; Sailike, J.; Sun, X.; Nabi, X. Composite probiotics alleviate type 2 diabetes by regulating intestinal microbiota and inducing GLP-1 secretion in db/db mice. Biomed. Pharmacother. 2020, 125, 109914. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Y.; Sailike, J.; Sun, X.; Abuduwaili, N.; Tuoliuhan, H.; Yusufu, M.; Nabi, X. Fourteen composite probiotics alleviate type 2 diabetes through modulating gut microbiota and modifying M1/M2 phenotype macrophage in db/db mice. Pharmacol. Res. 2020, 161, 105150. [Google Scholar] [CrossRef]
- Machado, A.S.; Oliveira, J.R.; Lelis, D.d.F.; de Paula, A.M.B.; Guimarães, A.L.S.; Andrade, J.M.O.; Brandi, I.V.; Santos, S.H.S. Oral Probiotic Bifidobacterium Longum Supplementation Improves Metabolic Parameters and Alters the Expression of the Renin-Angiotensin System in Obese Mice Liver. Biol. Res. Nurs. 2020, 23, 100–108. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Lee, D.; Park, G.-S.; Ko, S.-H.; Park, J.; Lee, Y.-K.; Kang, J. Lactobacillus plantarum HAC01 ameliorates type 2 diabetes in high-fat diet and streptozotocin-induced diabetic mice in association with modulating the gut microbiota. Food Funct. 2021, 12, 6363–6373. [Google Scholar] [CrossRef]
- Zeng, Z.; Yuan, Q.; Yu, R.; Zhang, J.; Ma, H.; Chen, S. Ameliorative Effects of Probiotic Lactobacillus paracasei NL41 on Insulin Sensitivity, Oxidative Stress, and Beta-Cell Function in a Type 2 Diabetes Mellitus Rat Model. Mol. Nutr. Food Res. 2019, 63, 1900457. [Google Scholar] [CrossRef]
- Ashrafian, F.; Shahriary, A.; Behrouzi, A.; Moradi, H.R.; Keshavarz Azizi Raftar, S.; Lari, A.; Hadifar, S.; Yaghoubfar, R.; Ahmadi Badi, S.; Khatami, S.; et al. Akkermansia muciniphila-Derived Extracellular Vesicles as a Mucosal Delivery Vector for Amelioration of Obesity in Mice. Front. Microbiol. 2019, 10, 2155. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ba, T.; Cheng, Y.; Zhang, P.; Chang, X. Probiotics alleviate adipose inflammation in high-fat diet-induced obesity by restoring adipose invariant natural killer T cells. Nutrition 2021, 89, 111285. [Google Scholar] [CrossRef] [PubMed]
- Tarrah, A.; dos Santos Cruz, B.C.; Sousa Dias, R.; da Silva Duarte, V.; Pakroo, S.; Licursi de Oliveira, L.; Gouveia Peluzio, M.C.; Corich, V.; Giacomini, A.; Oliveira de Paula, S. Lactobacillus paracasei DTA81, a cholesterol-lowering strain having immunomodulatory activity, reveals gut microbiota regulation capability in BALB/c mice receiving high-fat diet. J. Appl. Microbiol. 2021, 131, 1942–1957. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Zhang, Y.; Li, Z.; Yan, H.; Li, J.; Wan, X. Mechanism analysis of improved glucose homeostasis and cholesterol metabolism in high-fat-induced obese mice treated with La-SJLH001 via transcriptomics and culturomics. Food Funct. 2019, 10, 3556–3566. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, L.; Zhang, J.; Li, Y.; He, Q.; Li, H.; Guo, X.; Guo, J.; Zhang, H. Probiotic Lactobacillus casei Zhang ameliorates high-fructose-induced impaired glucose tolerance in hyperinsulinemia rats. Eur. J. Nutr. 2014, 53, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Li, N.; Shi, J.; Li, H.; Yue, Y.; Jiao, W.; Wang, N.; Song, Y.; Huo, G.; Li, B. Lactobacillus acidophilus alleviates type 2 diabetes by regulating hepatic glucose, lipid metabolism and gut microbiota in mice. Food Funct. 2019, 10, 5804–5815. [Google Scholar] [CrossRef]
- Bonfili, L.; Cecarini, V.; Gogoi, O.; Berardi, S.; Scarpona, S.; Angeletti, M.; Rossi, G.; Eleuteri, A.M. Gut microbiota manipulation through probiotics oral administration restores glucose homeostasis in a mouse model of Alzheimer’s disease. Neurobiol. Aging 2020, 87, 35–43. [Google Scholar] [CrossRef]
- Rodrigues, R.R.; Gurung, M.; Li, Z.; García-Jaramillo, M.; Greer, R.; Gaulke, C.; Bauchinger, F.; You, H.; Pederson, J.W.; Vasquez-Perez, S.; et al. Transkingdom interactions between Lactobacilli and hepatic mitochondria attenuate western diet-induced diabetes. Nat. Commun. 2021, 12, 101. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Y.; Wang, B.; Xu, H.; Mei, X.; Xu, X.; Zhang, X.; Ni, J.; Li, W. Bacillus amyloliquefaciens SC06 Protects Mice Against High-Fat Diet-Induced Obesity and Liver Injury via Regulating Host Metabolism and Gut Microbiota. Front. Microbiol. 2019, 10, 1161. [Google Scholar] [CrossRef]
- Park, K.-Y.; Kim, B.; Hyun, C.-K. Lactobacillus rhamnosus GG improves glucose tolerance through alleviating ER stress and suppressing macrophage activation in db/db mice. J. Clin. Biochem. Nutr. 2015, 56, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.-J.; Chiu, C.-C.; Lee, M.-C.; Huang, W.-C. Combination of Treadmill Aerobic Exercise with Bifidobacterium longum OLP-01 Supplementation for Treatment of High-Fat Diet-Induced Obese Murine Model. Obes. Facts 2021, 14, 306–319. [Google Scholar] [CrossRef]
- Kim, S.J.; Choi, S.I.; Jang, M.; Jeong, Y.A.; Kang, C.H.; Kim, G.H. Combination of Limosilactobacillus fermentum MG4231 and MG4244 attenuates lipid accumulation in high-fat diet-fed obese mice. Benef. Microbes 2021, 12, 479–491. [Google Scholar] [CrossRef]
- Bonfili, L.; Cecarini, V.; Berardi, S.; Scarpona, S.; Suchodolski, J.S.; Nasuti, C.; Fiorini, D.; Boarelli, M.C.; Rossi, G.; Eleuteri, A.M. Microbiota modulation counteracts Alzheimer’s disease progression influencing neuronal proteolysis and gut hormones plasma levels. Sci. Rep. 2017, 7, 2426. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, Q.; Dang, H.; Liu, X.; Tian, F.; Zhao, J.; Chen, Y.; Zhang, H.; Chen, W. Oral administration of Lactobacillus rhamnosus CCFM0528 improves glucose tolerance and cytokine secretion in high-fat-fed, streptozotocin-induced type 2 diabetic mice. J. Funct. Foods 2014, 10, 318–326. [Google Scholar] [CrossRef]
- Komori, T. Functions of Osteocalcin in Bone, Pancreas, Testis, and Muscle. Int. J. Mol. Sci. 2020, 21, 7513. [Google Scholar] [CrossRef]
- Zoch, M.L.; Clemens, T.L.; Riddle, R.C. New insights into the biology of osteocalcin. Bone 2016, 82, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Zanatta, L.C.B.; Boguszewski, C.L.; Borba, V.Z.C.; Kulak, C.A.M. Osteocalcin, energy and glucose metabolism. Arq. Bras. Endocrinol. Metabol. 2014, 58, 444–451. [Google Scholar] [CrossRef] [Green Version]
- Dang, F.; Jiang, Y.; Pan, R.; Zhou, Y.; Wu, S.; Wang, R.; Zhuang, K.; Zhang, W.; Li, T.; Man, C. Administration of Lactobacillus paracasei ameliorates type 2 diabetes in mice. Food Funct. 2018, 9, 3630–3639. [Google Scholar] [CrossRef]
- Stavropoulou, E.; Bezirtzoglou, E. Probiotics in Medicine: A Long Debate. Front. Immunol. 2020, 11, 2192. [Google Scholar] [CrossRef]
- Hylemon, P.B.; Takabe, K.; Dozmorov, M.; Nagahashi, M.; Zhou, H. Bile acids as global regulators of hepatic nutrient metabolism. Liver Res. 2017, 1, 10–16. [Google Scholar] [CrossRef]
- Martoni, C.J.; Labbé, A.; Ganopolsky, J.G.; Prakash, S.; Jones, M.L. Changes in bile acids, FGF-19 and sterol absorption in response to bile salt hydrolase active L. reuteri NCIMB 30242. Gut Microbes 2015, 6, 57–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, J.A.; Luo, G.; Billin, A.N.; Bisi, J.; McNeill, Y.Y.; Kozarsky, K.F.; Donahee, M.; Wang, D.Y.; Mansfield, T.A.; Kliewer, S.A.; et al. Definition of a novel growth factor-dependent signal cascade for the suppression of bile acid biosynthesis. Genes Dev. 2003, 17, 1581–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Z.; Zhang, W.; Du, R.; Meng, H.; Zhang, H. Lactobacillus casei Zhang stimulates lipid metabolism in hypercholesterolemic rats by affecting gene expression in the liver. Eur. J. Lipid Sci. Technol. 2012, 114, 244–252. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, X.; Guo, J.; He, Q.; Li, H.; Song, Y.; Zhang, H. Lactobacillus casei reduces susceptibility to type 2 diabetes via microbiota-mediated body chloride ion influx. Sci. Rep. 2014, 4, 5654. [Google Scholar] [CrossRef] [Green Version]
- Bleau, C.; Karelis, A.D.; St-Pierre, D.H.; Lamontagne, L. Crosstalk between intestinal microbiota, adipose tissue and skeletal muscle as an early event in systemic low-grade inflammation and the development of obesity and diabetes. Diabetes. Metab. Res. Rev. 2015, 31, 545–561. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Balakumar, M.; Prabhu, D.; Sathishkumar, C.; Prabu, P.; Rokana, N.; Kumar, R.; Raghavan, S.; Soundarajan, A.; Grover, S.; Batish, V.K.; et al. Improvement in glucose tolerance and insulin sensitivity by probiotic strains of Indian gut origin in high-fat diet-fed C57BL/6J mice. Eur. J. Nutr. 2018, 57, 279–295. [Google Scholar] [CrossRef]
- Lim, S.-M.; Jeong, J.-J.; Woo, K.H.; Han, M.J.; Kim, D.-H. Lactobacillus sakei OK67 ameliorates high-fat diet–induced blood glucose intolerance and obesity in mice by inhibiting gut microbiota lipopolysaccharide production and inducing colon tight junction protein expression. Nutr. Res. 2016, 36, 337–348. [Google Scholar] [CrossRef]
- Yoon, Y.; Kim, G.; Noh, M.; Park, J.; Jang, M.; Fang, S.; Park, H. Lactobacillus fermentum promotes adipose tissue oxidative phosphorylation to protect against diet-induced obesity. Exp. Mol. Med. 2020, 52, 1574–1586. [Google Scholar] [CrossRef]
- Wang, T.; Yan, H.; Lu, Y.; Li, X.; Wang, X.; Shan, Y.; Yi, Y.; Liu, B.; Zhou, Y.; Lü, X. Anti-obesity effect of Lactobacillus rhamnosus LS-8 and Lactobacillus crustorum MN047 on high-fat and high-fructose diet mice base on inflammatory response alleviation and gut microbiota regulation. Eur. J. Nutr. 2019, 59, 2709–2728. [Google Scholar] [CrossRef]
- Deng, L.; Ou, Z.; Huang, D.; Li, C.; Lu, Z.; Liu, W.; Wu, F.; Nong, C.; Gao, J.; Peng, Y. Diverse effects of different Akkermansia muciniphila genotypes on Brown adipose tissue inflammation and whitening in a high-fat-diet murine model. Microb. Pathog. 2020, 147, 104353. [Google Scholar] [CrossRef]
- Carvalho-Filho, M.A.; Ueno, M.; Hirabara, S.M.; Seabra, A.B.; Carvalheira, J.B.C.; de Oliveira, M.G.; Velloso, L.A.; Curi, R.; Saad, M.J.A. S-Nitrosation of the Insulin Receptor, Insulin Receptor Substrate 1, and Protein Kinase B/Akt: A Novel Mechanism of Insulin Resistance. Diabetes 2005, 54, 959–967. [Google Scholar] [CrossRef] [Green Version]
- Fang, H.; Judd, R.L. Adiponectin Regulation and Function. Compr. Physiol. 2018, 8, 1031–1063. [Google Scholar] [CrossRef]
- Le, T.K.C.; Hosaka, T.; Nguyen, T.T.; Kassu, A.; Dang, T.O.; Tran, H.B.; Pham, T.P.; Tran, Q.B.; Le, T.H.H.; Pham, X. Da Bifidobacterium species lower serum glucose, increase expressions of insulin signaling proteins, and improve adipokine profile in diabetic mice. Biomed. Res. 2015, 36, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Liu, Y.; Li, F.; Gu, Z.; Liu, M.; Shao, T.; Zhang, L.; Zhou, G.; Pan, C.; He, L.; et al. Probiotic culture supernatant improves metabolic function through FGF21-adiponectin pathway in mice. J. Nutr. Biochem. 2020, 75, 108256. [Google Scholar] [CrossRef]
- Tyagi, S.; Gupta, P.; Saini, A.S.; Kaushal, C.; Sharma, S. The peroxisome proliferator-activated receptor: A family of nuclear receptors role in various diseases. J. Adv. Pharm. Technol. Res. 2011, 2, 236–240. [Google Scholar] [CrossRef]
- Molina-Tijeras, J.A.; Diez-Echave, P.; Vezza, T.; Hidalgo-García, L.; Ruiz-Malagón, A.J.; Rodríguez-Sojo, M.J.; Romero, M.; Robles-Vera, I.; García, F.; Plaza-Diaz, J.; et al. Lactobacillus fermentum CECT5716 ameliorates high fat diet-induced obesity in mice through modulation of gut microbiota dysbiosis. Pharmacol. Res. 2021, 167, 105471. [Google Scholar] [CrossRef]
- Kwon, J.; Kim, B.; Lee, C.; Joung, H.; Kim, B.K.; Choi, I.S.; Hyun, C.K. Comprehensive amelioration of high-fat diet-induced metabolic dysfunctions through activation of the PGC-1α pathway by probiotics treatment in mice. PLoS ONE 2020, 15, e0228932. [Google Scholar] [CrossRef] [Green Version]
- López-Bermejo, A.; Botas-Cervero, P.; Ortega-Delgado, F.; Delgado, E.; García-Gil, M.M.; Funahashi, T.; Ricart, W.; Fernández-Real, J.M. Association of ADIPOR2 With Liver Function Tests in Type 2 Diabetic Subjects. Obesity 2008, 16, 2308–2313. [Google Scholar] [CrossRef]
- Olefsky, J.M.; Saltiel, A.R. PPAR gamma and the treatment of insulin resistance. Trends Endocrinol. Metab. 2000, 11, 362–368. [Google Scholar] [CrossRef]
- Mueckler, M.; Thorens, B. The SLC2 (GLUT) family of membrane transporters. Mol. Aspects Med. 2013, 34, 121–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Ding, Q.; Nie, S.-P.; Zhang, Y.-S.; Xiong, T.; Xie, M.-Y. Carrot Juice Fermented with Lactobacillus plantarum NCU116 Ameliorates Type 2 Diabetes in Rats. J. Agric. Food Chem. 2014, 62, 11884–11891. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liang, X.; Lv, Y.; Yi, H.; Chen, Y.; Bai, L.; Zhou, H.; Liu, T.; Li, R.; Zhang, L. Evaluation of probiotics for improving and regulation metabolism relevant to type 2 diabetes in vitro. J. Funct. Foods 2020, 64, 103664. [Google Scholar] [CrossRef]
- Yadav, H.; Quijano, C.; Kamaraju, A.K.; Gavrilova, O.; Malek, R.; Chen, W.; Zerfas, P.; Zhigang, D.; Wright, E.C.; Stuelten, C.; et al. Protection from obesity and diabetes by blockade of TGF-β/Smad3 signaling. Cell Metab. 2011, 14, 67–79. [Google Scholar] [CrossRef] [Green Version]
- Sousa-Pinto, B.; Gonçalves, L.; Rodrigues, A.R.; Tomada, I.; Almeida, H.; Neves, D.; Gouveia, A.M. Characterization of TGF-β expression and signaling profile in the adipose tissue of rats fed with high-fat and energy-restricted diets. J. Nutr. Biochem. 2016, 38, 107–115. [Google Scholar] [CrossRef]
- Lee, E.; Jung, S.-R.; Lee, S.-Y.; Lee, N.-K.; Paik, H.-D.; Lim, S.-I. Lactobacillus plantarum Strain Ln4 Attenuates Diet-Induced Obesity, Insulin Resistance, and Changes in Hepatic mRNA Levels Associated with Glucose and Lipid Metabolism. Nutrients 2018, 10, 643. [Google Scholar] [CrossRef] [Green Version]
- Flamment, M.; Hajduch, E.; Ferré, P.; Foufelle, F. New insights into ER stress-induced insulin resistance. Trends Endocrinol. Metab. 2012, 23, 381–390. [Google Scholar] [CrossRef]
- Kim, S.-H.; Huh, C.-S.; Choi, I.-D.; Jeong, J.-W.; Ku, H.-K.; Ra, J.-H.; Kim, T.-Y.; Kim, G.-B.; Sim, J.-H.; Ahn, Y.-T. The anti-diabetic activity of Bifidobacterium lactis HY8101 in vitro and in vivo. J. Appl. Microbiol. 2014, 117, 834–845. [Google Scholar] [CrossRef]
- Kim, S.-W.; Park, K.-Y.; Kim, B.; Kim, E.; Hyun, C.-K. Lactobacillus rhamnosus GG improves insulin sensitivity and reduces adiposity in high-fat diet-fed mice through enhancement of adiponectin production. Biochem. Biophys. Res. Commun. 2013, 431, 258–263. [Google Scholar] [CrossRef]
- Oberley, L.W. Free radicals and diabetes. Free Radic. Biol. Med. 1988, 5, 113–124. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Zhao, J.; Zhang, H.; Chen, W. Lactobacillus casei CCFM419 attenuates type 2 diabetes via a gut microbiota dependent mechanism. Food Funct. 2017, 8, 3155–3164. [Google Scholar] [CrossRef]
- Calcinaro, F.; Dionisi, S.; Marinaro, M.; Candeloro, P.; Bonato, V.; Marzotti, S.; Corneli, R.B.; Ferretti, E.; Gulino, A.; Grasso, F.; et al. Oral probiotic administration induces interleukin-10 production and prevents spontaneous autoimmune diabetes in the non-obese diabetic mouse. Diabetologia 2005, 48, 1565–1575. [Google Scholar] [CrossRef] [Green Version]
- Heeney, D.D.; Zhai, Z.; Bendiks, Z.; Barouei, J.; Martinic, A.; Slupsky, C.; Marco, M.L. Lactobacillus plantarum bacteriocin is associated with intestinal and systemic improvements in diet-induced obese mice and maintains epithelial barrier integrity in vitro. Gut Microbes 2018, 10, 382–397. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Gao, J.; Li, C.; Nong, C.; Huang, W.; Zheng, X.; Li, S.; Peng, Y. A potential probiotic bacterium for antipsychotic-induced metabolic syndrome: Mechanisms underpinning how Akkermansia muciniphila subtype improves olanzapine-induced glucose homeostasis in mice. Psychopharmacology 2021, 238, 2543–2553. [Google Scholar] [CrossRef]
- Park, S.-S.; Yang, G.; Kim, E. Lactobacillus acidophilus NS1 Reduces Phosphoenolpyruvate Carboxylase Expression by Regulating HNF4α Transcriptional Activity. Korean J. Food Sci. Anim. Resour. 2017, 37, 529–534. [Google Scholar] [CrossRef] [Green Version]
- Walesky, C.; Apte, U. Role of hepatocyte nuclear factor 4α (HNF4α) in cell proliferation and cancer. Gene Expr. 2015, 16, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, L.; Zhou, L.; Song, Y.; Ma, S.; Yu, C.; Zhao, J.; Xu, C.; Gao, L. Thyroid stimulating hormone increases hepatic gluconeogenesis via CRTC2. Mol. Cell. Endocrinol. 2017, 446, 70–80. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, L.; Zhang, Y.; Liu, X.; Wu, Z.; Gilbert, R.G.; Deng, B.; Wang, K. Dendrobium officinale polysaccharide ameliorates diabetic hepatic glucose metabolism via glucagon-mediated signaling pathways and modifying liver-glycogen structure. J. Ethnopharmacol. 2020, 248, 112308. [Google Scholar] [CrossRef]
- Fountain, J.H.; Lappin, S.L. Physiology, Renin Angiotensin System; StatPearls: Treasure Island, FL, USA, 2021. [Google Scholar]
- Xu, J.; Fan, J.; Wu, F.; Huang, Q.; Guo, M.; Lv, Z.; Han, J.; Duan, L.; Hu, G.; Chen, L.; et al. The ACE2/angiotensin-(1-7)/Mas receptor axis: Pleiotropic roles in cancer. Front. Physiol. 2017, 8, 276. [Google Scholar] [CrossRef]
- Santos, S.H.S.; Fernandes, L.R.; Mario, É.G.; Ferreira, A.V.M.; Pôrto, L.C.J.; Alvarez-Leite, J.I.; Botion, L.M.; Bader, M.; Alenina, N.; Santos, R.A.S. Mas Deficiency in FVB/N Mice Produces Marked Changes in Lipid and Glycemic Metabolism. Diabetes 2008, 57, 340–347. [Google Scholar] [CrossRef] [Green Version]
- Bilman, V.; Mares-Guia, L.; Nadu, A.P.; Bader, M.; Campagnole-Santos, M.J.; Santos, R.A.S.; Santos, S.H.S. Decreased hepatic gluconeogenesis in transgenic rats with increased circulating angiotensin-(1-7). Peptides 2012, 37, 247–251. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xu, Q.; Jiang, T.; Fang, S.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. A comparative study of the antidiabetic effects exerted by live and dead multi-strain probiotics in the type 2 diabetes model of mice. Food Funct. 2016, 7, 4851–4860. [Google Scholar] [CrossRef]
- Niibo, M.; Shirouchi, B.; Umegatani, M.; Morita, Y.; Ogawa, A.; Sakai, F.; Kadooka, Y.; Sato, M. Probiotic Lactobacillus gasseri SBT2055 improves insulin secretion in a diabetic rat model. J. Dairy Sci. 2019, 102, 997–1006. [Google Scholar] [CrossRef] [Green Version]
- Wright, E.M.; Loo, D.D.F.; Hirayama, B.A. Biology of Human Sodium Glucose Transporters. Physiol. Rev. 2011, 91, 733–794. [Google Scholar] [CrossRef] [Green Version]
- Korkmaz, O.A.; Sumlu, E.; Koca, H.B.; Pektas, M.B.; Kocabas, A.; Sadi, G.; Akar, F.; Korkmaz, O.A.; Sumlu, E.; Koca, H.B.; et al. Effects of Lactobacillus Plantarum and Lactobacillus Helveticus on Renal Insulin Signaling, Inflammatory Markers, and Glucose Transporters in High-Fructose-Fed Rats. Medicina 2019, 55, 207. [Google Scholar] [CrossRef] [Green Version]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Voreades, N.; Kozil, A.; Weir, T.L. Diet and the development of the human intestinal microbiome. Front. Microbiol. 2014, 5, 494. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [Green Version]
- Stenvinkel, P. Obesity-a disease with many aetiologies disguised in the same oversized phenotype: Has the overeating theory failed? Nephrol. Dial. Transplant 2014, 30, 1656–1664. [Google Scholar] [CrossRef] [Green Version]
- Kallus, S.J.; Brandt, L.J. The intestinal microbiota and obesity. J. Clin. Gastroenterol. 2012, 46, 16–24. [Google Scholar] [CrossRef]
- Santacruz, A.; Collado, M.C.; García-Valdés, L.; Segura, M.T.; Martín-Lagos, J.A.; Anjos, T.; Martí-Romero, M.; Lopez, R.M.; Florido, J.; Campoy, C.; et al. Gut microbiota composition is associated with body weight, weight gain and biochemical parameters in pregnant women. Br. J. Nutr. 2010, 104, 83–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Armougom, F.; Henry, M.; Vialettes, B.; Raccah, D.; Raoult, D. Monitoring bacterial community of human gut microbiota reveals an increase in Lactobacillus in obese patients and Methanogens in anorexic patients. PLoS ONE 2009, 4, e7125. [Google Scholar] [CrossRef] [PubMed]
- Million, M.; Maraninchi, M.; Henry, M.; Armougom, F.; Richet, H.; Carrieri, P.; Valero, R.; Raccah, D.; Vialettes, B.; Raoult, D. Obesity-associated gut microbiota is enriched in Lactobacillus reuteri and depleted in Bifidobacterium animalis and Methanobrevibacter smithii. Int. J. Obes. 2012, 36, 817–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collado, M.C.; Isolauri, E.; Laitinen, K.; Salminen, S. Distinct composition of gut microbiota during pregnancy in overweight and normal-weight women. Am. J. Clin. Nutr. 2008, 88, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Schwiertz, A.; Taras, D.; Schäfer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in lean and overweight healthy subjects. Obesity 2010, 18, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Mai, V.; McCrary, Q.M.; Sinha, R.; Glei, M. Associations between dietary habits and body mass index with gut microbiota composition and fecal water genotoxicity: An observational study in African American and Caucasian American volunteers. Nutr. J. 2009, 8, 49. [Google Scholar] [CrossRef] [Green Version]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.-M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- Million, M.; Lagier, J.-C.; Yahav, D.; Paul, M. Gut bacterial microbiota and obesity. Clin. Microbiol. Infect. 2013, 19, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Larsen, N.; Vogensen, F.K.; van den Berg, F.W.J.; Nielsen, D.S.; Andreasen, A.S.; Pedersen, B.K.; Al-Soud, W.A.; Sørensen, S.J.; Hansen, L.H.; Jakobsen, M. Gut Microbiota in Human Adults with Type 2 Diabetes Differs from Non-Diabetic Adults. PLoS ONE 2010, 5, e9085. [Google Scholar] [CrossRef]
- Ivey, K.L.; Hodgson, J.M.; Kerr, D.A.; Lewis, J.R.; Thompson, P.L.; Prince, R.L. The effects of probiotic bacteria on glycaemic control in overweight men and women: A randomised controlled trial. Eur. J. Clin. Nutr. 2014, 68, 447–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solito, A.; Bozzi Cionci, N.; Calgaro, M.; Caputo, M.; Vannini, L.; Hasballa, I.; Archero, F.; Giglione, E.; Ricotti, R.; Walker, G.E.; et al. Supplementation with Bifidobacterium breve BR03 and B632 strains improved insulin sensitivity in children and adolescents with obesity in a cross-over, randomized double-blind placebo-controlled trial. Clin. Nutr. 2021, 40, 4585–4594. [Google Scholar] [CrossRef] [PubMed]
- Zavratnik, A.; Krajnc, M. Analysis of pregnant women with the diagnosis of gestational diabetes mellitus without oral glucose tolerance test confirmation in early pregnancy. Acta Med. -Biotech. 2018, 11, 27–36. [Google Scholar]
- Laitinen, K.; Poussa, T.; Isolauri, E.; Nutrition, Allergy, Mucosal Immunology and Intestinal Microbiota Group. Probiotics and dietary counselling contribute to glucose regulation during and after pregnancy: A randomised controlled trial. Br. J. Nutr. 2008, 101, 1679–1687. [Google Scholar] [CrossRef] [Green Version]
- Dallanora, S.; Medeiros de Souza, Y.; Deon, R.G.; Tracey, C.A.; Freitas-Vilela, A.A.; Wurdig Roesch, L.F.; Hack Mendes, R. Do probiotics effectively ameliorate glycemic control during gestational diabetes? A systematic review. Arch. Gynecol. Obstet. 2018, 298, 477–485. [Google Scholar] [CrossRef]
- Karamali, M.; Dadkhah, F.; Sadrkhanlou, M.; Jamilian, M.; Ahmadi, S.; Tajabadi-Ebrahimi, M.; Jafari, P.; Asemi, Z. Effects of probiotic supplementation on glycaemic control and lipid profiles in gestational diabetes: A randomized, double-blind, placebo-controlled trial. Diabetes Metab. 2016, 42, 234–241. [Google Scholar] [CrossRef]
- Wickens, K.L.; Barthow, C.A.; Murphy, R.; Abels, P.R.; Maude, R.M.; Stone, P.R.; Mitchell, E.A.; Stanley, T.V.; Purdie, G.L.; Kang, J.M.; et al. Early pregnancy probiotic supplementation with Lactobacillus rhamnosus HN001 may reduce the prevalence of gestational diabetes mellitus: A randomised controlled trial. Br. J. Nutr. 2017, 117, 804–813. [Google Scholar] [CrossRef] [Green Version]
- Dolatkhah, N.; Hajifaraji, M.; Abbasalizadeh, F.; Aghamohammadzadeh, N.; Mehrabi, Y.; Mesgari Abbasi, M. Is there a value for probiotic supplements in gestational diabetes mellitus? A randomized clinical trial. J. Health Popul. Nutr. 2015, 33, 25. [Google Scholar] [CrossRef] [Green Version]
- Sahhaf Ebrahimi, F.; Homayouni Rad, A.; Mosen, M.; Abbasalizadeh, F.; Tabrizi, A.; Khalili, L. Effect of L. acidophilus and B. lactis on blood glucose in women with gestational diabetes mellitus: A randomized placebo-controlled trial. Diabetol. Metab. Syndr. 2019, 11, 75. [Google Scholar] [CrossRef]
- Shahriari, A.; Karimi, E.; Shahriari, M.; Aslani, N.; Khooshideh, M.; Arab, A. The effect of probiotic supplementation on the risk of gestational diabetes mellitus among high-risk pregnant women: A parallel double-blind, randomized, placebo-controlled clinical trial. Biomed. Pharmacother. 2021, 141, 111915. [Google Scholar] [CrossRef]
- Tsigalou, C.; Konstantinidis, T.; Stavropoulou, E.; Bezirtzoglou, E.E.; Tsakris, A. Potential Elimination of Human Gut Resistome by Exploiting the Benefits of Functional Foods. Front. Microbiol. 2020, 11, 50. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Huang, Y.; Song, L.; Xiao, Y.; Lu, S.; Xu, J.; Li, J.; Ren, Z. Lactobacillus plantarum prevents obesity via modulation of gut microbiota and metabolites in high-fat feeding mice. J. Funct. Foods 2020, 73, 104103. [Google Scholar] [CrossRef]
- Li, X.; Wang, E.; Yin, B.; Fang, D.; Chen, P.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Effects of Lactobacillus casei CCFM419 on insulin resistance and gut microbiota in type 2 diabetic mice. Benef. Microbes 2017, 8, 421–432. [Google Scholar] [CrossRef]
- Li, K.K.; Tian, P.J.; Wang, S.D.; Lei, P.; Qu, L.; Huang, J.P.; Shan, Y.J.; Li, B. long Targeting gut microbiota: Lactobacillus alleviated type 2 diabetes via inhibiting LPS secretion and activating GPR43 pathway. J. Funct. Foods 2017, 38, 561–570. [Google Scholar] [CrossRef]
- Luo, J.; Zhang, H.; Lu, J.; Ma, C.L.; Chen, T. Antidiabetic effect of an engineered bacterium Lactobacillus plantarum-pMG36e -GLP-1 in monkey model. Synth. Syst. Biotechnol. 2021, 6, 272–282. [Google Scholar] [CrossRef]
- Zheng, Q.-X.; Jiang, X.-M.; Wang, H.-W.; Ge, L.; Lai, Y.-T.; Jiang, X.-Y.; Chen, F.; Huang, P.-P. Probiotic supplements alleviate gestational diabetes mellitus by restoring the diversity of gut microbiota: A study based on 16S rRNA sequencing. J. Microbiol. 2021, 59, 827–839. [Google Scholar] [CrossRef]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. SCFA: Mechanisms and functional importance in the gut. Proc. Nutr. Soc. 2021, 80, 37–49. [Google Scholar] [CrossRef]
- Primec, M.; Mičetić-Turk, D.; Langerholc, T. Analysis of short-chain fatty acids in human feces: A scoping review. Anal. Biochem. 2017, 526, 9–21. [Google Scholar] [CrossRef]
- Yadav, H.; Lee, J.-H.; Lloyd, J.; Walter, P.; Rane, S.G. Beneficial Metabolic Effects of a Probiotic via Butyrate-induced GLP-1 Hormone Secretion. J. Biol. Chem. 2013, 288, 25088–25097. [Google Scholar] [CrossRef] [Green Version]
- Troy, S.; Soty, M.; Ribeiro, L.; Laval, L.; Migrenne, S.; Fioramonti, X.; Pillot, B.; Fauveau, V.; Aubert, R.; Viollet, B.; et al. Intestinal Gluconeogenesis Is a Key Factor for Early Metabolic Changes after Gastric Bypass but Not after Gastric Lap-Band in Mice. Cell Metab. 2008, 8, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Le Poul, E.; Loison, C.; Struyf, S.; Springael, J.Y.; Lannoy, V.; Decobecq, M.E.; Brezillon, S.; Dupriez, V.; Vassart, G.; Van Damme, J.; et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J. Biol. Chem. 2003, 278, 25481–25489. [Google Scholar] [CrossRef] [Green Version]
- Horiuchi, H.; Kamikado, K.; Aoki, R.; Suganuma, N.; Nishijima, T.; Nakatani, A.; Kimura, I. Bifidobacterium animalis subsp. lactis GCL2505 modulates host energy metabolism via the short-chain fatty acid receptor GPR43. Sci. Rep. 2020, 10, 4158. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Qiao, S.; Peng, C.; Yan, W.; Xu, Z.; Qu, J.; Hou, Y.; Zhao, S.; Chen, P.; Wang, T. Bornlisy Attenuates Colitis-Associated Colorectal Cancer via Inhibiting GPR43-Mediated Glycolysis. Front. Nutr. 2021, 8, 835. [Google Scholar] [CrossRef]
- Drucker, D.J. Mechanisms of Action and Therapeutic Application of Glucagon-like Peptide-1. Cell Metab. 2018, 27, 740–756. [Google Scholar] [CrossRef] [Green Version]
- Nauck, M.A.; Meier, J.J. Incretin hormones: Their role in health and disease. Diabetes Obes. Metab. 2018, 20, 5–21. [Google Scholar] [CrossRef]
- Simon, M.-C.; Strassburger, K.; Nowotny, B.; Kolb, H.; Nowotny, P.; Burkart, V.; Zivehe, F.; Hwang, J.-H.; Stehle, P.; Pacini, G.; et al. Intake of Lactobacillus reuteri Improves Incretin and Insulin Secretion in Glucose-Tolerant Humans: A Proof of Concept. Diabetes Care 2015, 38, 1827–1834. [Google Scholar] [CrossRef] [Green Version]
- Bjerg, A.T.; Kristensen, M.; Ritz, C.; Holst, J.J.; Rasmussen, C.; Leser, T.D.; Wellejus, A.; Astrup, A. Lactobacillus paracasei subsp paracasei L. casei W8 suppresses energy intake acutely. Appetite 2014, 82, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Yadav, H.; Jain, S.; Sinha, P.R. Oral administration of dahi containing probiotic Lactobacillus acidophilus and Lactobacillus casei delayed the progression of streptozotocin-induced diabetes in rats. J. Dairy Res. 2008, 75, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Yamano, T.; Tanida, M.; Niijima, A.; Maeda, K.; Okumura, N.; Fukushima, Y.; Nagai, K. Effects of the probiotic strain Lactobacillus johnsonii strain La1 on autonomic nerves and blood glucose in rats. Life Sci. 2006, 79, 1963–1967. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Liu, Q.; Zhou, Z.; Ikeda, Y. PDX1, Neurogenin-3, and MAFA: Critical transcription regulators for beta cell development and regeneration. Stem Cell Res. Ther. 2017, 8, 240. [Google Scholar] [CrossRef] [Green Version]
- Duan, F.; Curtis, K.L.; March, J.C. Secretion of Insulinotropic Proteins by Commensal Bacteria: Rewiring the Gut To Treat Diabetes. Appl. Environ. Microbiol. 2008, 74, 7437–7438. [Google Scholar] [CrossRef] [Green Version]
- Belguesmia, Y.; Alard, J.; Mendil, R.; Ravallec, R.; Grangette, C.; Drider, D.; Cudennec, B. In vitro probiotic properties of selected lactobacilli and multi-strain consortium on immune function, gut barrier strengthening and gut hormone secretion. J. Funct. Foods 2019, 57, 382–391. [Google Scholar] [CrossRef]
- Hung, S.-C.; Tseng, W.-T.; Pan, T.-M. Lactobacillus paracasei subsp. paracasei NTU 101 ameliorates impaired glucose tolerance induced by a high-fat, high-fructose diet in Sprague-Dawley rats. J. Funct. Foods 2016, 24, 472–481. [Google Scholar] [CrossRef]
- Tian, P.; Li, B.; He, C.; Song, W.; Hou, A.; Tian, S.; Meng, X.; Li, K.; Shan, Y. Antidiabetic (type 2) effects of Lactobacillus G15 and Q14 in rats through regulation of intestinal permeability and microbiota. Food Funct. 2016, 7, 3789–3797. [Google Scholar] [CrossRef]
- Rooj, A.K.; Kimura, Y.; Buddington, R.K. Metabolites produced by probiotic Lactobacilli rapidly increase glucose uptake by Caco-2 cells. BMC Microbiol. 2010, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Primec, M.; Škorjanc, D.; Langerholc, T.; Mičetić-Turk, D.; Gorenjak, M. Specific Lactobacillus probiotic strains decrease transepithelial glucose transport through GLUT2 downregulation in intestinal epithelial cell models. Nutr. Res. 2021, 86, 10–22. [Google Scholar] [CrossRef]
- Kolb, H.; Kempf, K.; Röhling, M.; Lenzen-Schulte, M.; Schloot, N.C.; Martin, S. Ketone bodies: From enemy to friend and guardian angel. BMC Med. 2021, 19, 313. [Google Scholar] [CrossRef]
- Liao, C.A.; Huang, C.H.; Ho, H.H.; Chen, J.F.; Kuo, Y.W.; Lin, J.H.; Tsai, S.Y.; Tsai, H.Y.; Yeh, Y.T. A Combined Supplement of Probiotic Strains AP-32, bv-77, and CP-9 Increased Akkermansia mucinphila and Reduced Non-Esterified Fatty Acids and Energy Metabolism in HFD-Induced Obese Rats. Nutrients 2022, 14, 527. [Google Scholar] [CrossRef]
- Zhang, Z.; Mocanu, V.; Cai, C.; Dang, J.; Slater, L.; Deehan, E.C.; Walter, J.; Madsen, K.L. Impact of Fecal Microbiota Transplantation on Obesity and Metabolic Syndrome-A Systematic Review. Nutrients 2019, 11, 2291. [Google Scholar] [CrossRef] [Green Version]
- Aron-Wisnewsky, J.; Clément, K.; Nieuwdorp, M. Fecal Microbiota Transplantation: A Future Therapeutic Option for Obesity/Diabetes? Curr. Diab. Rep. 2019, 19, 51. [Google Scholar] [CrossRef] [PubMed]
- Huda, M.N.; Kim, M.; Bennett, B.J. Modulating the Microbiota as a Therapeutic Intervention for Type 2 Diabetes. Front. Endocrinol. 2021, 12, 632335. [Google Scholar] [CrossRef] [PubMed]
- Allegretti, J.R.; Kassam, Z.; Hurtado, J.; Marchesi, J.R.; Mullish, B.H.; Chiang, A.; Thompson, C.C.; Cummings, B.P. Impact of fecal microbiota transplantation with capsules on the prevention of metabolic syndrome among patients with obesity. Hormones 2021, 20, 209–211. [Google Scholar] [CrossRef] [PubMed]
- Proença, I.M.; Allegretti, J.R.; Bernardo, W.M.; de Moura, D.T.H.; Ponte Neto, A.M.; Matsubayashi, C.O.; Flor, M.M.; Kotinda, A.P.S.T.; de Moura, E.G.H. Fecal microbiota transplantation improves metabolic syndrome parameters: Systematic review with meta-analysis based on randomized clinical trials. Nutr. Res. 2020, 83, 1–14. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Altered Blood Parameters | Type of Probiotic | Physiological/ Pathological Condition | Reference |
|---|---|---|---|
| ↓ Postprandial blood glucose | Lacticaseibacillus casei 01 (paraprobiotic) | None | [42] |
| ↓ FPG | Lactobacillus acidophilus La-5 and Bifidobacterium animalis subsp lactis BB-12 | T2D | [43] |
| Lactobacillus acidophilus La-5 and Bifidobacterium animalis subsp lactis BB-12 | Metabolic syndrome | [44] | |
| Bifidobacterium longum APC1472 | Healthy overweight/obese | [45] | |
| Lactobacillus plantarum OLL2712 | Pre-diabetic | [46] | |
| Ecologic® Barrier (multi-strain probiotics) | T2D | [47] | |
| Lactobacillus casei 01 | T2D | [48] | |
| Lactobacillus casei | T2D | [49] | |
| ↓ HbA1c | Lactobacillus acidophilus La-5 and Bifidobacterium animalis subsp lactis BB-12 | T2D | [50] |
| Lactobacillus reuteri ADR-1 | T2D | [51] | |
| “Symbiter” (multi-strain) | T2D | [52] | |
| Lactobacillus acidophilus La-5 and Bifidobacterium animalis subsp lactis BB-12 | T2D | [43] | |
| Lactobacillus acidophilus La-5 and Bifidobacterium animalis subsp lactis BB-12 | T2D | [53] | |
| HOMA-IR improvement (reduction) | Lactobacillus plantarum OLL2712 | Pre-diabetic | [46] |
| “Symbiter” (multi-strain) | T2D | [52] | |
| Ecologic® Barrier (multi-strain probiotics) | T2D | [47] | |
| Lactobacillus casei 01 | T2D | [48] | |
| Lactobacillus casei | T2D | [49] | |
| Lactobacillus acidophilus La-5 and Bifidobacterium animalis subsp lactis BB-12 | Metabolic syndrome | [44] | |
| ↓ Fructosamine | Lactobacillus acidophilus La-5 and Bifidobacterium animalis subsp lactis BB-12 | T2D | [50] |
| ↓ Fasting insulin levels | Ecologic® Barrier (multi-strain probiotics) | T2D | [47] |
| Lactobacillus casei | T2D | [49] | |
| Lactobacillus casei 01 | T2D | [48] | |
| Lactobacillus acidophilus La-5 and Bifidobacterium animalis subsp lactis BB-12 | Metabolic syndrome | [44] | |
| Insulin sensitivity improvement (↑ ISI, QUICKI test or others) | Lactobacillus reuteri DSM 17938 | T2D | [54] |
| Ecologic® Barrier (multi-strain probiotics) | T2D | [47] | |
| Lactobacillus acidophilus NCFM | T2D | [55] | |
| Lactobacillus plantarum OLL2712 | Pre-diabetic | [46] | |
| Lactobacillus acidophilus La-5 and Bifidobacterium animalis subsp lactis BB-12 | Metabolic syndrome | [44] | |
| ↓ Ghrelin | Bifidobacterium longum APC1472 | Healthy overweight/obese | [45] |
| ↓ Cortisol | Bifidobacterium longum APC1472 | Healthy overweight/obese | [45] |
| ↓ Glycoalbumin | Lactobacillus plantarum OLL2712 | Pre-diabetic | [46] |
| ↓ Fetuin-A | Lactobacillus casei 01 | T2D | [48] |
| Lactobacillus casei | T2D | [49] | |
| ↑ SIRTs | Lactobacillus casei 01 | T2D | [48] |
| Lactobacillus casei | T2D | [49] | |
| ↓ IL-6 | Lactobacillus plantarum OLL2712 | Pre-diabetic | [46] |
| Ecologic® Barrier (multi-strain probiotics) | T2D | [47] | |
| ↓ TNF-α | “Symbiter” (multi-strain) | T2D | [52] |
| Lactobacillus plantarum OLL2712 | Pre-diabetic | [46] | |
| Lactobacillus acidophilus La-5 and Bifidobacterium animalis subsp lactis BB-12 | T2D | [50] | |
| Ecologic® Barrier (multi-strain probiotics) | T2D | [47] | |
| ↓ IL-1β | “Symbiter” (multi-strain) | T2D | [52] |
| Lactobacillus plantarum OLL2712 | Pre-diabetic | [46] | |
| Lactobacillus reuteri ADR-3 | T2D | [51] | |
| ↓ MCP-1 | Lactobacillus plantarum OLL2712 | Pre-diabetic | [46] |
| Altered Blood Parameters | Type of Probiotic | Species | Physiological/Pathological Condition and/or Diet | Reference |
|---|---|---|---|---|
| ↓ Postprandial blood glucose | Lactobacillus rhamnosus (YC, 7-1), Bifidobacterium adolescentis (N3, 7-2) and Bifidobacterium bifidum (M2) | Mouse | HFD STZ-induced diabetes | [77] |
| Lactobacillus plantarum CCFM0236 | Mouse | STZ-induced diabetes | [78] | |
| ↓ FPG | Lactobacillus casei and Bifidobacterium bifidum (alone or combination of both) | rat | STZ-induced diabetes | [79] |
| Lactobacillus plantarum TN627 | Rat | Alloxan-induced diabetes | [80] | |
| Bifidobacterium animalis 01 | Rat | High-fat chow diet STZ-induced diabetes | [81] | |
| Lactobacillus paracasei HII01 | Rat | HFD STZ-induced diabetes | [82] | |
| Lactobacillus rhamnosus (YC, 7-1), Bifidobacterium adolescentis (N3) and Bifidobacterium bifidum (M2) | Mouse | HFD STZ-induced diabetes | [77] | |
| Lactobacillus rhamnosus (BSL and R23) | Rat | STZ-induced diabetes | [83] | |
| Heat-inactivated Streptococcus thermophilus | Rat | HFD ZDF diabetes | [84] | |
| Lactiplantibacillus plantarum IMC 510 | Rat | Diet-induced obesity | [85] | |
| Lactobacillus fermentum RS-2 | Rat | Alloxan-induced diabetes | [86] | |
| Bacillus licheniformis Zhengchangsheng® | Mouse | HFD-induced obesity | [87] | |
| Weissella cibaria MG5285, Limosilactobacillus reuteri MG5149, Lacticaseibacillus rhamnosus MG4502, Lactobacillus gasseri MG4524 | Mouse | HFD-induced obesity | [88] | |
| ProbiogluTM | Rat | STZ-induced diabetes | [89] | |
| Bacillus toyonensis SAU-19 | Mouse | HFD ZDF-induced diabetes | [90] | |
| Lactobacillus casei LC89 | Mouse | STZ-induced diabetes | [91] | |
| Lactibiane Tolérance® | Mouse | HFD | [92] | |
| Lactobacillus salivarus AP-32 Lactobacillus reuteri GL-104 | Mouse | Db/db obesity | [93] | |
| Lactobacillus plantarum CCFM0236 | Mouse | STZ-induced diabetes | [78] | |
| Lactobacillus plantarum YJ7 | Mouse | HFD | [94] | |
| Bifidobacterium longum BR-108 (inactivated) | Mouse | Obese diabetes | [95] | |
| L. rhamnosus L12, L. acidophilus, L. plantarum HM218749, B. animalis subsp. lactis LPL-RH, B. longum subsp. longum BAMA-B05/BAu-B1024) | Mouse | Glucose STZ-induced diabetes | [96] | |
| L. rhamnosus L12, L. acidophilus, L. plantarum HM218749, B. animalis subsp. lactis LPL-RH, B. longum subsp. longum BAMA-B05/BAu-B1024) | Mouse | Glucose water -induced diabetes | [96] | |
| Lactobacillus plantarum DSM 15313 | Mouse | HFD-induced obesity | [97] | |
| Lactobacillus plantarum OLL2712 | Mouse | HFD-induced obesity | [98] | |
| Lactobacillus casei Shirota YIT 9029 | Mouse | HFD-induced obesity | [99] | |
| 14 composite probiotics | Mouse | Db/db diabetes | [100] | |
| 14 composite probiotics | Mouse | Db/db diabetes | [101] | |
| Bifidobacterium longum | Mouse | HFD-induced obesity | [102] | |
| Lactobacillus plantarum HAC01 | Mouse | HFD STZ-induced diabetes | [103] | |
| Lactobacillus paracasei NL41 | Rat | HFD STZ-induced diabetes | [104] | |
| Akkermansia municiphila and its extracellular vesicles | Mouse | HFD | [105] | |
| VSL#3 composite probiotics | Mouse | HFD-induced obesity | [106] | |
| Lactobacillus paracasei DTA81 | Mouse | HFD | [107] | |
| Lactobacillus acidophilus SJLH001 | Mouse | HFD | [108] | |
| Lactobacillus casei Zhang | Rat | Hyperinsulinemia | [109] | |
| Lactobacillus acidophilus KLDS1.1003 and KLDS1.0901 | Mouse | HFD STZ-induced diabetes | [110] | |
| ↓ HbA1c | Lactobacillus casei and Bifidobacterium bifidum (alone or combination of both) | Rat | STZ-induced diabetes | [79] |
| Lactobacillus reuteri ADR-1 | Rat | High-fructose diet | [51] | |
| Bifidobacterium animalis 01 | Rat | High-fat chow diet STZ-induced diabetes | [81] | |
| Heat-inactivated Streptococcus thermophilus | Rat | HFD ZDF diabetes | [84] | |
| Lactobacillus plantarum CCFM0236 | Mouse | STZ-induced diabetes | [78] | |
| 14 composite probiotics | Mouse | Db/db diabetes | [100] | |
| 14 composite probiotics | Mouse | Db/db diabetes | [101] | |
| Lactobacillus plantarum HAC01 | Mouse | HFD STZ-induced diabetes | [103] | |
| Lactobacillus paracasei NL41 | Rat | HFD STZ-induced diabetes | [104] | |
| Lactobacillus acidophilus KLDS1.1003 and KLDS1.0901 | Mouse | HFD STZ-induced diabetes | [110] | |
| SLAB51 composite probiotics | Mouse | AD | [111] | |
| HOMA-IR improvement (reduction) | Bifidobacterium animalis 01 | Rat | High-fat chow diet STZ-induced diabetes | [81] |
| Lactobacillus paracasei HII01 | Rat | HFD STZ-induced diabetes | [82] | |
| Lactobacillus rhamnosus (YC), Bifidobacterium adolescentis (N3) and Bifidobacterium bifidum (M2) | Mouse | HFD STZ-induced diabetes | [77] | |
| Heat-inactivated Streptococcus thermophilus | Rat | HFD ZDF diabetes | [84] | |
| Bacillus licheniformis Zhengchangsheng® | Mouse | HFD-induced obesity | [87] | |
| ProbiogluTM | Rat | STZ-induced diabetes | [89] | |
| Bacillus toyonensis SAU-19 | Mouse | HFD ZDF-induced diabetes | [90] | |
| Lactobacillus casei LC89 | Mouse | STZ-induced diabetes | [91] | |
| Lactibiane Tolérance® | Mouse | HFD | [92] | |
| Lactobacillus plantarum CCFM0236 | Mouse | STZ-induced diabetes | [78] | |
| Lactobacillus plantarum YJ7 | Mouse | HFD | [94] | |
| Lactobacillus plantarum DSM 15313 | Mouse | HFD-induced obesity | [97] | |
| 14 composite probiotics | Mouse | Db/db diabetes | [100] | |
| 14 composite probiotics | Mouse | Db/db diabetes | [101] | |
| Lactobacillus plantarum HAC01 | Mouse | HFD STZ-induced diabetes | [103] | |
| Lactobacillus paracasei NL41 | Rat | HFD STZ-induced diabetes | [104] | |
| VSL#3 composite probiotics | Mouse | HFD-induced obesity | [106] | |
| Lactobacillus acidophilus KLDS1.0901 | Mouse | HFD STZ-induced diabetes | [110] | |
| HOMA-B reduction | Romboutsia ilealis DSM 25109 | Mouse | Western diet-induced diabetes | [112] |
| ↓ fasting insulin levels | Lactobacillus paracasei HII01 | Rat | HFD STZ-induced diabetes | [82] |
| Heat-inactivated Streptococcus thermophilus | Rat | HFD ZDF diabetes | [84] | |
| Bacillus licheniformis Zhengchangsheng® | Mouse | HFD-induced obesity | [87] | |
| ProbiogluTM | Rat | STZ-induced diabetes | [89] | |
| Lactobacillus casei LC89 | Mouse | STZ-induced diabetes | [91] | |
| Romboutsia ilealis DSM 25109 | Mouse | Western diet-induced diabetes | [112] | |
| Lactibiane Tolérance® | Mouse | HFD | [92] | |
| Lactobacillus plantarum CCFM0236 | Mouse | STZ-induced diabetes | [78] | |
| Lactobacillus acidophilus KLDS1.0901 | Mouse | HFD STZ-induced diabetes | [110] | |
| ↑ insulin levels | Lactobacillus casei and Bifidobacterium bifidum (alone or combination of both) | Rat | STZ-induced diabetes | [79] |
| Lactobacillus rhamnosus (BSL and R23) | Rat | STZ-induced diabetes | [83] | |
| 14 composite probiotics | Mouse | Db/db diabetes | [100] | |
| Insulin sensitivity improvement (↑ ISI, QUICKI test or others) | Lactobacillus rhmanosus (GG, YC), Bifidobacterium adolescentis (7-2) and Bifidobacterium bifidum (35) | Mouse | HFD STZ-induced diabetes | [77] |
| Lactobacillus rhamnosus (BSL and R23) | Rat | STZ-induced diabetes | [83] | |
| Bacillus amyloliquefaciens SC06 | Mouse | HFD-induced obesity | [113] | |
| Lactobacillus plantarum CCFM0236 | Mouse | STZ-induced diabetes | [78] | |
| Lactobacillus plantarum DSM 15313 | Mouse | HFD-induced obesity | [97] | |
| Lactobacillus acidophilus KLDS1.0901 | Mouse | HFD STZ-induced diabetes | [110] | |
| Glucose tolerance improvement (OGTT) | Bifidobacterium longum APC1472 | Mouse | HFD | [45] |
| Lactobacillus casei and Bifidobacterium bifidum (alone or combination of both) | Rat | STZ-induced diabetes | [79] | |
| Lactobacillus rhmanosus, Bifidobacterium adolescentis and Bifidobacterium bifidum (all together 9 strains) | Mouse | HFD STZ-induced diabetes | [77] | |
| Lactobacillus paracasei HII01 | Rat | HFD STZ-induced diabetes | [82] | |
| Bifidobacterium animalis 01 | Rat | High-fat chow diet STZ-induced diabetes | [81] | |
| Lactobacillus rhamnosus (BSL and R23) | Rat | STZ-induced diabetes | [83] | |
| Heat-inactivated Streptococcus thermophilus | Rat | HFD ZDF diabetes | [84] | |
| Weissella cibaria MG5285, Limosilactobacillus reuteri MG5149, Lacticaseibacillus rhamnosus MG4502, Lactobacillus gasseri MG4524 | Mouse | HFD-induced obesity | [88] | |
| Romboutsia ilealis DSM 25109 | Mouse | Western diet-induced diabetes | [112] | |
| ProbiogluTM | Rat | STZ-induced diabetes | [89] | |
| Lactobacillus salivarus AP-32 Lactobacillus reuteri GL-104 | Mouse | Db/db obesity | [93] | |
| Bifidobacterium longum BR-108 (inactivated) | Mouse | Obese diabetes | [95] | |
| L. rhamnosus L12, L. acidophilus, L. plantarum HM218749, B. animalis subsp. lactis LPL-RH, B. longum subsp. longum BAMA-B05/BAu-B1024) | Mouse | Glucose STZ-induced diabetes | [96] | |
| L. rhamnosus L12, L. acidophilus, L. plantarum HM218749, B. animalis subsp. lactis LPL-RH, B. longum subsp. longum BAMA-B05/BAu-B1024) | Mouse | Glucose water- Induced diabetes | [96] | |
| Lactobacillus rhamnosus GG | Mouse | Db/db obesity | [114] | |
| Bifidobacterium longum | Mouse | HFD-induced obesity | [102] | |
| Lactobacillus plantarum HAC01 | Mouse | HFD STZ-induced diabetes | [103] | |
| Bifidobacterium longum OLP-01 | Mouse | HFD-induced obesity | [115] | |
| Limosilactobacillus fermentum MG4231 and MG4244 | Mouse | HFD-induced obesity | [116] | |
| Lactobacillus paracasei NL41 | Rat | HFD STZ-induced diabetes | [104] | |
| VSL#3 composite probiotics | Mouse | HFD-induced obesity | [106] | |
| Lactobacillus acidophilus SJLH001 | Mouse | HFD | [108] | |
| Lactobacillus casei Zhang | Rat | Hyperinsulinemia | [109] | |
| ↑ osteocalcin | Lactobacillus casei Zhang | Rat | Hyperinsulinemia | [109] |
| ↓ IL-6 | Heat-inactivated Streptococcus thermophilus | Rat | HFD ZDF-induced diabetes | [84] |
| ProbiogluTM | Rat | STZ-induced diabetes | [89] | |
| Lactobacillus plantarum YJ7 | Mouse | HFD-induced obesity | [94] | |
| Bacillus amyloliquefaciens SC06 | Mouse | HFD-induced obesity | [113] | |
| Lactobacillus casei LC89 | Mouse | STZ-induced diabetes | [91] | |
| ↓ TNF-α | Lactobacillus plantarum YJ7 | Mouse | HFD-induced obesity | [94] |
| Bacillus amyloliquefaciens SC06 | Mouse | HFD-induced obesity | [113] | |
| Heat-inactivated Streptococcus thermophilus | Rat | HFD ZDF-induced diabetes | [84] | |
| ProbiogluTM | Rat | STZ-induced diabetes | [89] | |
| Lactobacillus casei LC89 | Mouse | STZ-induced diabetes | [91] | |
| SLAB51 composite probiotics | Mouse | AD | [117] | |
| Bifidobacterium animalis 01 | Rat | High-fat chow diet STZ-induced diabetes | [81] | |
| Lactobacillus plantarum CCFM0236 | Mouse | HFD STZ-induced diabetes | [78] | |
| ↓ IL-1β | Lactobacillus casei LC89 | Mouse | STZ-induced diabetes | [91] |
| Lactobacillus plantarum YJ7 | Mouse | HFD-induced obesity | [94] | |
| Heat-inactivated Streptococcus thermophilus | Rat | HFD ZDF-induced diabetes | [84] | |
| ProbiogluTM | Rat | STZ-induced diabetes | [89] | |
| SLAB51 composite probiotics | Mouse | AD | [117] | |
| ↑ IL-10 | Heat-inactivated Streptococcus thermophilus | Rat | HFD ZDF-induced diabetes | [84] |
| Lactobacillus casei LC89 | Mouse | STZ-induced diabetes | [91] | |
| Bifidobacterium animalis 01 | Rat | High-fat chow diet STZ-induced diabetes | [81] | |
| Lactobacillus plantarum CCFM0236 | Mouse | HFD STZ-induced diabetes | [78] |
| Animal Model/ Pathology | Administered Probiotic | Altered Intestinal Microbiota | Effect on Glucose Metabolism | Reference | |
|---|---|---|---|---|---|
| Genera or Family | |||||
| Increase | Decrease | ||||
| T2D mouse | Lactobacillus casei LC89 | Alloprevotella (B), Bacteroides (B), Parabacteroides (B), Ruminococcus (F) | Lachnospiraceae _NK4A136_group (F), Odoribacter (B) Mucispirillum (D) | Glucose tolerance improvement | [91] |
| HFD- induced obese mouse | Lactobacillus plantarum HT121 | Akkermansia (V), Allobaculum (F), Lactobacillus (F), Prevotella (B), Sutterella (P) and S24-7 family (B) | Anaerostipes (F) Anaerotruncus (F) Bilophila (P) Candidatus_Arthromitus (F) Coprococcus (F), Dorea (F), Mucispirillum (D), Oscillospira (F), Ruminococcus (F), Streptococcus (F) and families of Peptococcaceae (F), Ruminococcaceae (F) | Glucose tolerance improvement; blood glucose decrease | [197] |
| HFD- induced obese mouse | Lactobacillus plantarum Q180 | Rikenellaceae (B), Ruminococcaceae (F), Lachnospiraceae (F) | glucose tolerance improvement | [142] | |
| T2D mouse | Lactobacillus casei CCFM419 | Allobaculum (F), Bacteroides (B) | Alleviation of type 2 diabetes symptoms (insulin resistance and hyperglycaemia amelioration) | [198] | |
| T2D mouse | Lactobacillus plantarum HAC01 | Akkermansiaceae (V) | Dusulfovibrionaceae (P) | Reduction in glucose-mediated insulin secretion (SCFA increase) | [103] |
| T2D mouse (db/db) | 14 composite probiotics | Bifidobacterium (A), Lactobacillus (F), Clostridium leptum (F), Roseburia (F), Prevotella (B), | Enterococcus faecium (F), Escherichia coli (P), Bacteroides thetaiotaomicron (B) | Improvement in glucose absorption | [101] |
| T2D rat | Lactobacillus G15 and Q14 | Clostridium leptum (F), Bacteroides (B), Prevotella (B) | Bifidobacterium (A), Lactobacillus (F) | Improvement in blood glucose and insulin disorders (glucose tolerance) | [199] |
| T2D mouse | *^Bifidobacterium adolescentis N3, *^Bifidobacterium bifidum M2, ^Bifidobacterium adolescentis 7-2, °Lactobacillus rhamnosus YC, ’Lactobacillus rhamnosus 7-1 | * Bacteroidales S24-7 (B), ^Parabacteroides (B), °Mucispirillum (D), ’Coprococcus (F), ’Streptococcus (F) | Glucose metabolism and insulin resistance improvement (reduction in blood glucose levels and insulin resistance, regulation of SCFAs levels) | [77] | |
| T2D mouse | Lactobacillus acidophilus KLDS1.003 and KLDS1.0901 | Blautia (F), Roseburia (F), Anaerotruncus (F) | Desulfovibrio (T), Alistipes (B) Bacteroides (B) | Glucose and lipid metabolism- related signalling route improvement of intestinal microbiota | [110] |
| T2D rat | Streptococcus thermophilus | Ruminococcaceae (F), Veillonella (F), Coprococcus (F), Barnesiella (B) | Moderation of insulin resistance (HOMA-IR, HbA1c improvement) | [84] | |
| T2D monkey | Lactobacillus plantarum - pMG36e-GLP-1 | Alistipes (B) | Prevotella (B) | Blood glucose reduction (increase in SCFAs) | [200] |
| GDM pregnant rat | Lactobacillus rhamnosus LGG and Bifidobacterium animalis subsp. lactis Bb12 | Lachnospiraceae (F), Dubosiella (F) | Muribaculaceae (F), Ruminococcacea_ UCG-005 (F) | Carbohydrate metabolism and membrane transport pathway inhibition | [201] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pintarič, M.; Langerholc, T. Probiotic Mechanisms Affecting Glucose Homeostasis: A Scoping Review. Life 2022, 12, 1187. https://doi.org/10.3390/life12081187
Pintarič M, Langerholc T. Probiotic Mechanisms Affecting Glucose Homeostasis: A Scoping Review. Life. 2022; 12(8):1187. https://doi.org/10.3390/life12081187
Chicago/Turabian StylePintarič, Maša, and Tomaž Langerholc. 2022. "Probiotic Mechanisms Affecting Glucose Homeostasis: A Scoping Review" Life 12, no. 8: 1187. https://doi.org/10.3390/life12081187