
Modularity of the Human Musculoskeletal System: The Correlation between Functional Structures by Computer Tools Analysis


 , ,
, ,  and
and 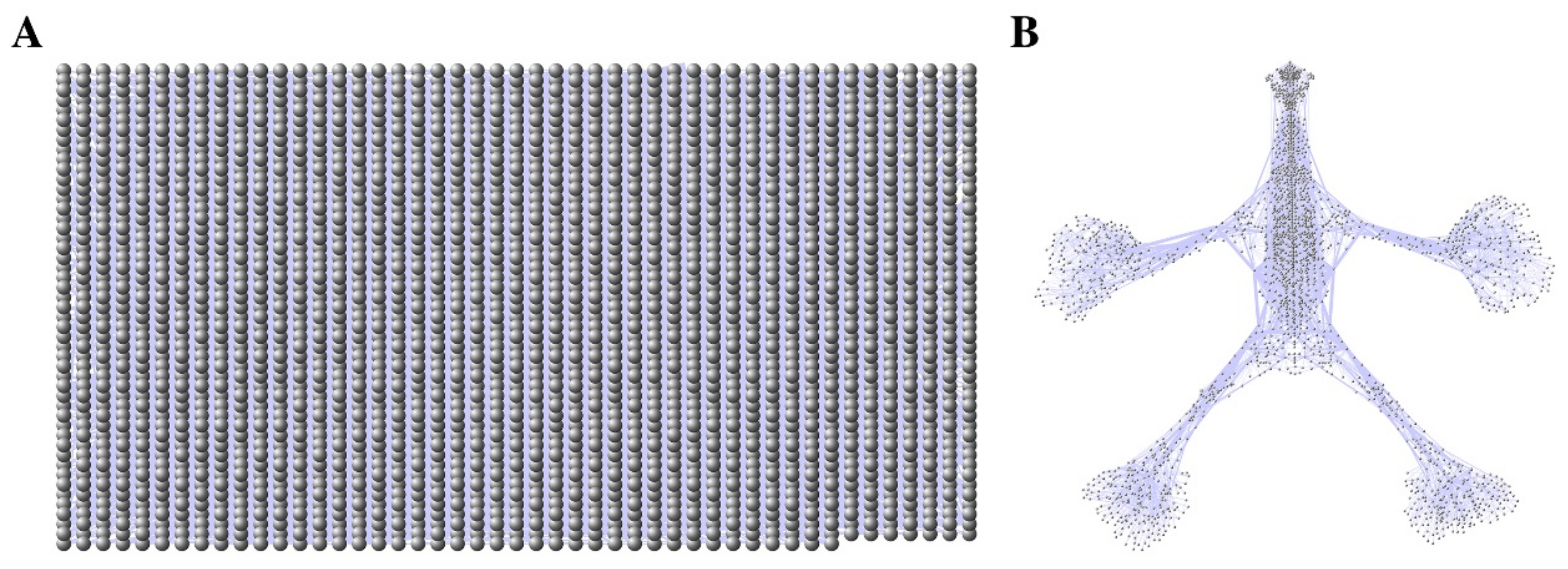{kind=link}
{kind=link}
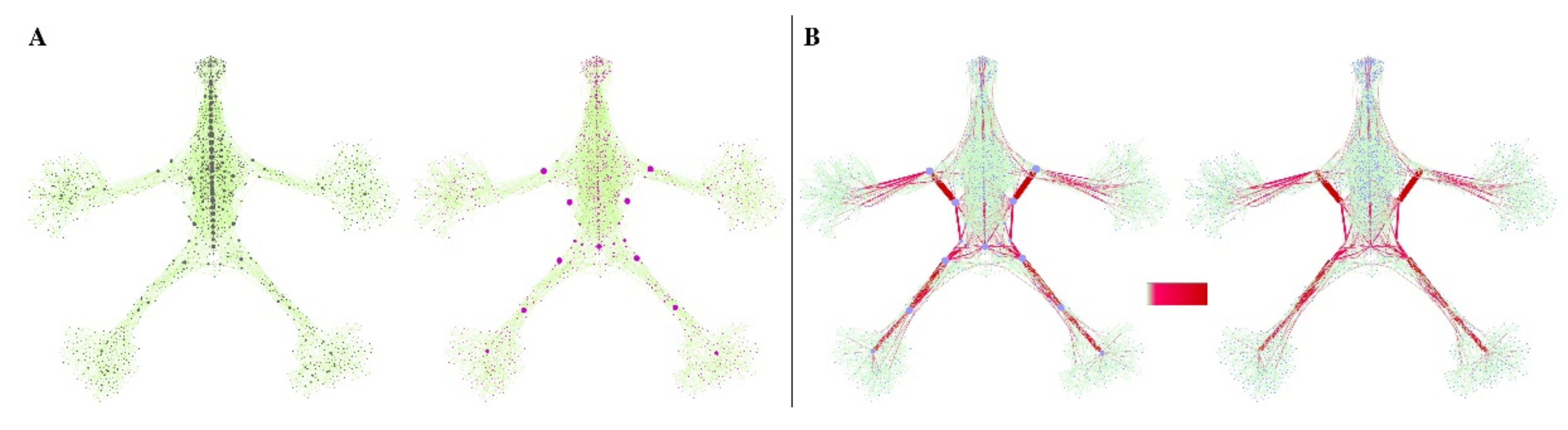
{kind=link}
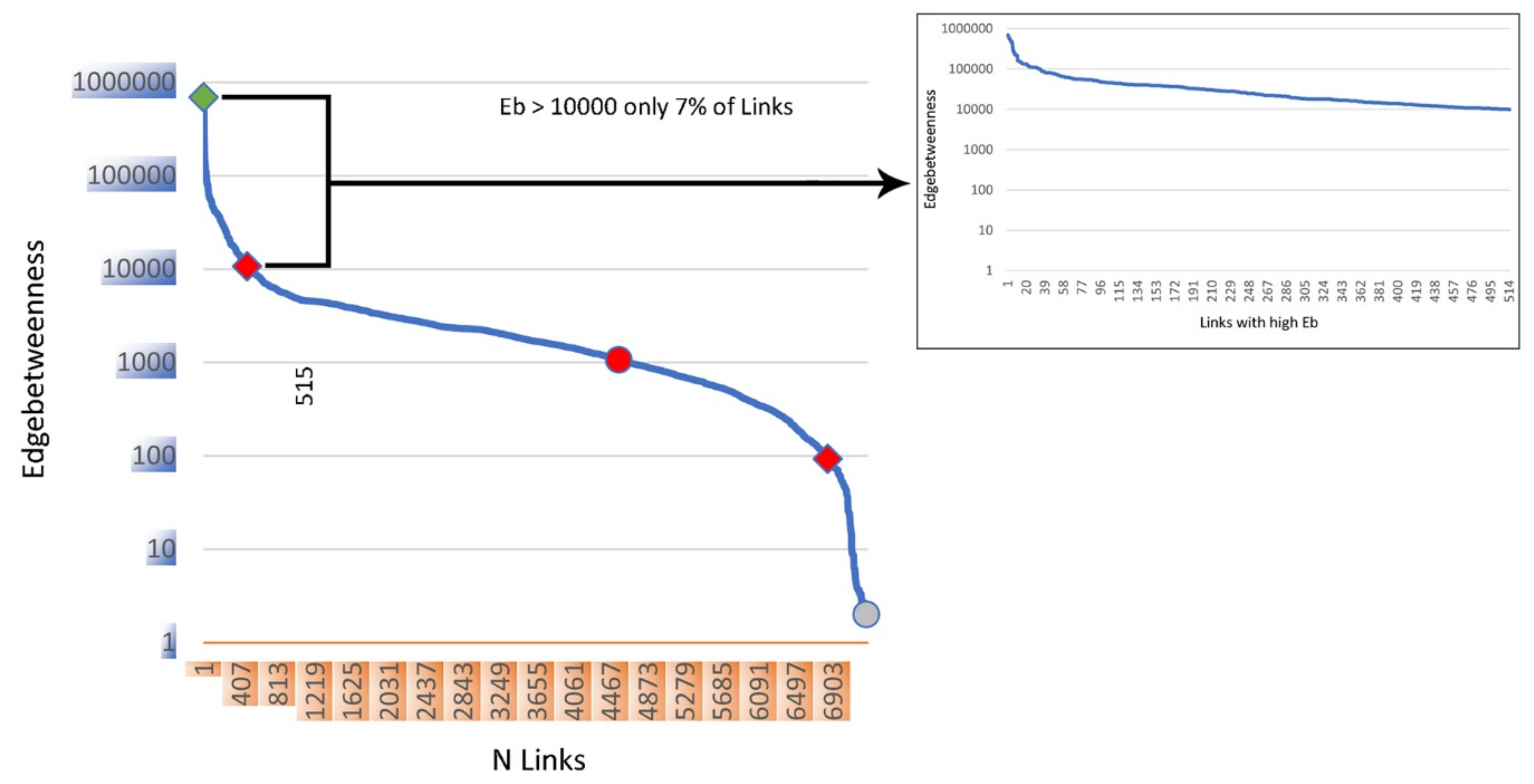
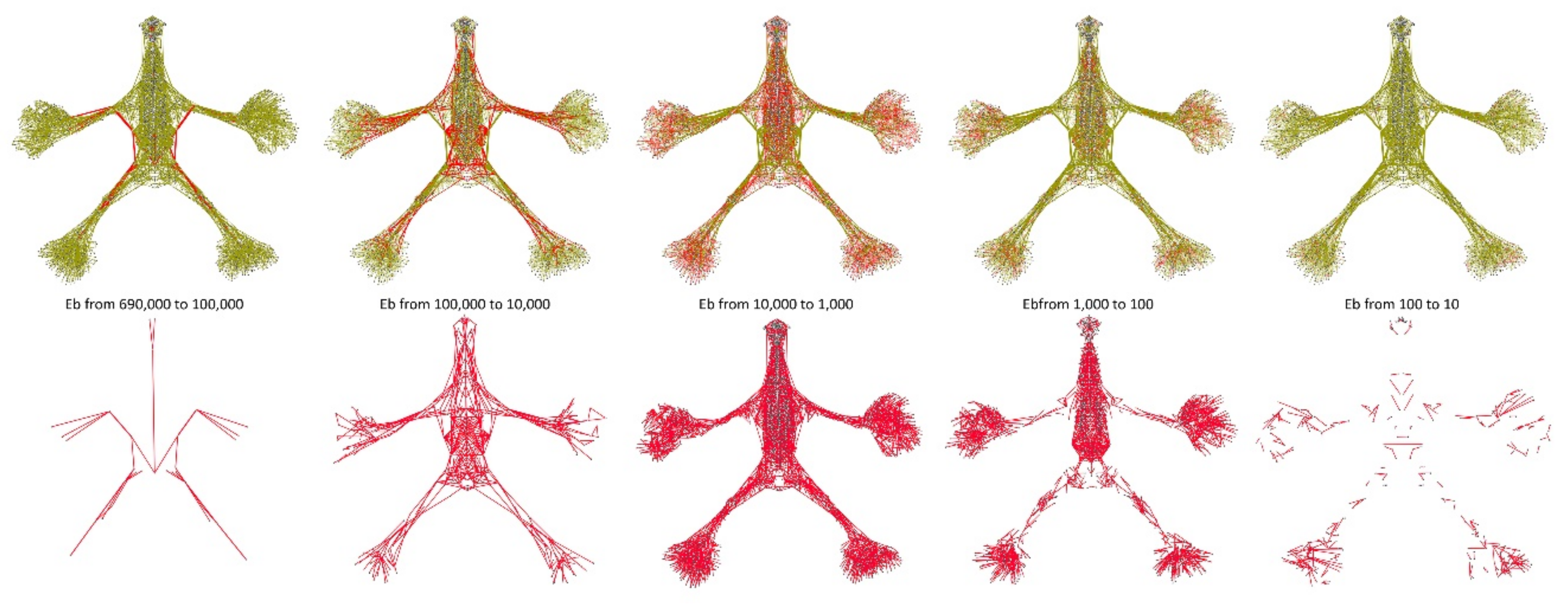{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Biological Modularity
1.2. The Modular Anatomical Organization of the Musculoskeletal System
1.3. Study on the Modularity of the Human Anatomical Biomechanical Network
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lipson, H.; Pollack, J.B.; Suh, N.P. On the Origin of Modular Variation. Evolution 2002, 56, 1549–1556. [Google Scholar] [CrossRef] [PubMed]
- Kashtan, N.; Mayo, A.E.; Kalisky, T.; Alon, U. An Analytically Solvable Model for Rapid Evolution of Modular Structure. PLoS Comput. Biol. 2009, 5, e1000355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogarad, L.D.; Deem, M.W. A Hierarchical Approach to Protein Molecular Evolution. Proc. Natl. Acad. Sci. USA 1999, 96, 2591–2595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rainey, P.B.; Cooper, T.F. Evolution of Bacterial Diversity and the Origins of Modularity. Res. Microbiol. 2004, 155, 370–375. [Google Scholar] [CrossRef]
- McAdams, H.H.; Srinivasan, B.; Arkin, A.P. The Evolution of Genetic Regulatory Systems in Bacteria. Nat. Rev. Genet. 2004, 5, 169–178. [Google Scholar] [CrossRef]
- Kitano, H. Biological Robustness. Nat. Rev. Genet. 2004, 5, 826–837. [Google Scholar] [CrossRef]
- Solé, R.V.; Valverde, S. Spontaneous Emergence of Modularity in Cellular Networks. J. R. Soc. Interface 2008, 5, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Callebaut, W.; Rasskin-Gutman, D. Modularity: Understanding the Development and Evolution of Natural Complex Systems; The MIT Press: Cambridge, MA, USA, 2005; ISBN 978-0-262-26969-8. [Google Scholar]
- Espinosa-Soto, C.; Wagner, A. Specialization Can Drive the Evolution of Modularity. PLoS Comput. Biol. 2010, 6, e1000719. [Google Scholar] [CrossRef]
- Litvin, O.; Causton, H.C.; Chen, B.-J.; Pe’er, D. Modularity and Interactions in the Genetics of Gene Expression. Proc. Natl. Acad. Sci. USA 2009, 106, 6441–6446. [Google Scholar] [CrossRef] [Green Version]
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Cluster Analysis and Display of Genome-Wide Expression Patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868. [Google Scholar] [CrossRef] [Green Version]
- Karlin, D.; Ferron, F.; Canard, B.; Longhi, S. Structural Disorder and Modular Organization in Paramyxovirinae N and P. J. Gen. Virol. 2003, 84, 3239–3252. [Google Scholar] [CrossRef]
- Serban, M. Exploring Modularity in Biological Networks. Phil. Trans. R. Soc. B 2020, 375, 20190316. [Google Scholar] [CrossRef] [Green Version]
- Scarr, G. Biotensegrity: What Is the Big Deal? J. Bodyw. Mov. Ther. 2020, 24, 134–137. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Wu, J.; Paulino, G.H.; Qi, H.J. Programmable Deployment of Tensegrity Structures by Stimulus-Responsive Polymers. Sci. Rep. 2017, 7, 3511. [Google Scholar] [CrossRef] [Green Version]
- Valero-Cuevas, F.J.; Yi, J.W.; Brown, D.; McNamara, R.V.; Paul, C.; Lipson, H. The Tendon Network of the Fingers Performs Anatomical Computation at a Macroscopic Scale. IEEE Trans. Biomed. Eng. 2007, 54, 1161–1166. [Google Scholar] [CrossRef]
- Csermely, P. Creative Elements: Network-Based Predictions of Active Centres in Proteins and Cellular and Social Networks. Trends Biochem. Sci. 2008, 33, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Gustafsson, M.; Nestor, C.E.; Zhang, H.; Barabási, A.-L.; Baranzini, S.; Brunak, S.; Chung, K.F.; Federoff, H.J.; Gavin, A.-C.; Meehan, R.R.; et al. Modules, Networks and Systems Medicine for Understanding Disease and Aiding Diagnosis. Genome Med. 2014, 6, 82. [Google Scholar] [CrossRef] [Green Version]
- Booij, G.; Lehmann, C.; Mugdan, J.; Kesselheim, W.; Skopeteas, S. Morphologie; De Gruyter Mouton: Berlin, Germany, 2004; ISBN 978-3-11-017278-2. [Google Scholar]
- Kapandji, I. Anatomia Funzionale; Maloine-Monduzzi: Paris, France, 2011; ISBN 88-6521-038-9. [Google Scholar]
- Diogo, R.; Ziermann, J.M.; Smith, C.; Alghamdi, M.; Fuentes, J.S.M.; Duerinckx, A. First Use of Anatomical Networks to Study Modularity and Integration of Heads, Forelimbs and Hindlimbs in Abnormal Anencephalic and Cyclopic vs. Normal Human Development. Sci. Rep. 2019, 9, 7821. [Google Scholar] [CrossRef]
- Diogo, R.; Esteve-Altava, B.; Smith, C.; Boughner, J.C.; Rasskin-Gutman, D. Anatomical Network Comparison of Human Upper and Lower, Newborn and Adult, and Normal and Abnormal Limbs, with Notes on Development, Pathology and Limb Serial Homology vs. Homoplasy. PLoS ONE 2015, 10, e0140030. [Google Scholar] [CrossRef]
- Powell, V.; Esteve-Altava, B.; Molnar, J.; Villmoare, B.; Pettit, A.; Diogo, R. Primate Modularity and Evolution: First Anatomical Network Analysis of Primate Head and Neck Musculoskeletal System. Sci. Rep. 2018, 8, 2341. [Google Scholar] [CrossRef] [Green Version]
- Paternostro, F.; Santosuosso, U.; Della Posta, D.; Francia, P. An Application of the Graph Theory to the Study of the Human Locomotor System. Italy J. Anat. Embryol. 2019, 124, 353–363. [Google Scholar] [CrossRef]
- Della Posta, D.; Gulisano, M.; Paternostro, F.; Santosuosso, U.; Veltro, C.; Francia, P.G. The Graph Theory Applied to the Study of the Human Locomotor System: A Simulated Amputation Changes the Characteristics of the System. In Proceedings of the 72nd National Meeting SIAI, Parma, Italy, 20–22 September 2018; FUPRESS: Florence, Italy, 2018; Volume 123, p. 76. [Google Scholar]
- Della Posta, D.; Veltro, C.; Santosuosso, U.; Zecchi, S.; Paternostro, F. Forces Distribution during Plantar Stand among the Myo-Osteo-Joint Components of the Foot. Simulations and Analysis on a Human Anatomical Network Model. In Proceedings of the 71st National Meeting SIAI, Taormina, Italy, 20–22 September 2017; FUPRESS: Florence, Italy, 2017; Volume 119, p. 73. [Google Scholar]
- Della Posta, D.; Veltro, C.; Paternostro, F. Anatomical Network Analysis Reveals Centralities in Human Biomechanical Structure. In Proceedings of the 70th National Meeting SIAI, Rome, Italy, 15–17 September 2016; FUPRESS: Florence, Italy, 2016; Volume 121, p. 212. [Google Scholar]
- Barabási, A.L.; Pósfai, M. Network Science; Cambridge University Press: Cambridge, UK, 2016; ISBN 978-1-107-07626-6. [Google Scholar]
- Della Posta, D. Network e Regolazione in Terapia Manuale; A.I.R.O.P.: Rome, Italy, 2021; ISBN 978-88-949401-4-5. [Google Scholar]
- Federative Commitee on Anatomical Terminology (FCAT). Terminologia Anatomica: International Anatomical Terminology; Georg Thieme Verlag: Stuttgart, Germany, 1998; ISBN 3-13-115251-6. [Google Scholar]
- Kachlik, D.; Baca, V.; Bozdechova, I.; Cech, P.; Musil, V. Anatomical Terminology and Nomenclature: Past, Present and Highlights. Surg. Radiol. Anat. 2008, 30, 459–466. [Google Scholar] [CrossRef]
- Allen, W.E. Terminologia Anatomica: International Anatomical Terminology and Terminologia Histologica: International Terms for Human Cytology and Histology. J. Anat. 2009, 215, 221. [Google Scholar] [CrossRef]
- Ten Donkelaar, H.J.; Broman, J.; Neumann, P.E.; Puelles, L.; Riva, A.; Tubbs, R.S.; Kachlik, D. Towards a Terminologia Neuroanatomica. Clin. Anat. 2017, 30, 145–155. [Google Scholar] [CrossRef]
- Musil, V.; Blankova, A.; Baca, V. A Plea for an Extension of the Anatomical Nomenclature: The Locomotor System. Bosn. J. Basic Med. Sci. 2018, 18, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Onorati, M.; Castiglioni, V.; Biasci, D.; Cesana, E.; Menon, R.; Vuono, R.; Talpo, F.; Goya, R.L.; Lyons, P.A.; Bulfamante, G.P.; et al. Molecular and Functional Definition of the Developing Human Striatum. Nat. Neurosci. 2014, 17, 1804–1815. [Google Scholar] [CrossRef]
- Assenov, Y.; Ramírez, F.; Schelhorn, S.-E.; Lengauer, T.; Albrecht, M. Computing Topological Parameters of Biological Networks. Bioinformatics 2008, 24, 282–284. [Google Scholar] [CrossRef] [Green Version]
- Szalay-Bekő, M.; Palotai, R.; Szappanos, B.; Kovács, I.A.; Papp, B.; Csermely, P. ModuLand Plug-in for Cytoscape: Determination of Hierarchical Layers of Overlapping Network Modules and Community Centrality. Bioinformatics 2012, 28, 2202–2204. [Google Scholar] [CrossRef] [Green Version]
- Kovács, I.A.; Palotai, R.; Szalay, M.S.; Csermely, P. Community Landscapes: An Integrative Approach to Determine Overlapping Network Module Hierarchy, Identify Key Nodes and Predict Network Dynamics. PLoS ONE 2010, 5, e12528. [Google Scholar] [CrossRef] [Green Version]
- Girvan, M.; Newman, M.E.J. Community Structure in Social and Biological Networks. Proc. Natl. Acad. Sci. USA 2002, 99, 7821–7826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, T.; Mezzarane, R.A.; Klarner, T.; Barss, T.S.; Hundza, S.R.; Komiyama, T.; Zehr, E.P. Neural Mechanisms Influencing Interlimb Coordination during Locomotion in Humans: Presynaptic Modulation of Forearm H-Reflexes during Leg Cycling. PLoS ONE 2013, 8, e76313. [Google Scholar] [CrossRef] [PubMed]
- Frigon, A. The Neural Control of Interlimb Coordination during Mammalian Locomotion. J. Neurophysiol. 2017, 117, 2224–2241. [Google Scholar] [CrossRef] [PubMed]
- Frigon, A.; Collins, D.F.; Zehr, E.P. Effect of Rhythmic Arm Movement on Reflexes in the Legs: Modulation of Soleus H-Reflexes and Somatosensory Conditioning. J. Neurophysiol. 2004, 91, 1516–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Standrig, S. Anatomia del Gray—Le basi Anatomiche per la Pratica Clinica; Edra: Milan, Italy, 2017; ISBN 8821441474. [Google Scholar]
- Myers, T.W. Meridiani Miofasciali. Percorsi Anatomici Per i Terapisti Del Corpo e Del Movimento; Tecniche Nuove: Milan, Italy, 2016; ISBN 88-481-3097-6. [Google Scholar]
- Rieffel, J.A.; Valero-Cuevas, F.J.; Lipson, H. Morphological Communication: Exploiting Coupled Dynamics in a Complex Mechanical Structure to Achieve Locomotion. J. R. Soc. Interface 2010, 7, 613–621. [Google Scholar] [CrossRef] [Green Version]
- Hauser, H.; Corucci, F. Morphosis—Taking Morphological Computation to the Next Level. In Soft Robotics: Trends, Applications and Challenges; Laschi, C., Rossiter, J., Iida, F., Cianchetti, M., Margheri, L., Eds.; Biosystems & Biorobotics; Springer International Publishing: Cham, Switzerland, 2017; Volume 17, pp. 117–122. ISBN 978-3-319-46459-6. [Google Scholar]
- Collins, S. Efficient Bipedal Robots Based on Passive-Dynamic Walkers. Science 2005, 307, 1082–1085. [Google Scholar] [CrossRef] [Green Version]
- Odifreddi, P. La Legge Della Pigrizia Cosmica; GEDI Gruppo Editoriale: Rome, Italy, 2015; p. 566. [Google Scholar]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Della Posta, D.; Branca, J.J.V.; Guarnieri, G.; Veltro, C.; Pacini, A.; Paternostro, F. Modularity of the Human Musculoskeletal System: The Correlation between Functional Structures by Computer Tools Analysis. Life 2022, 12, 1186. https://doi.org/10.3390/life12081186
Della Posta D, Branca JJV, Guarnieri G, Veltro C, Pacini A, Paternostro F. Modularity of the Human Musculoskeletal System: The Correlation between Functional Structures by Computer Tools Analysis. Life. 2022; 12(8):1186. https://doi.org/10.3390/life12081186
Chicago/Turabian StyleDella Posta, Daniele, Jacopo Junio Valerio Branca, Giulia Guarnieri, Cristiana Veltro, Alessandra Pacini, and Ferdinando Paternostro. 2022. "Modularity of the Human Musculoskeletal System: The Correlation between Functional Structures by Computer Tools Analysis" Life 12, no. 8: 1186. https://doi.org/10.3390/life12081186