
Phylogeographical Analysis of the Freshwater Gudgeon Huigobio chenhsienensis (Cypriniformes: Gobionidae) in Southern China



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Sequence Obtaining and Processing
2.3. Phylogeographic Analysis and Divergence Time Estimation
2.4. Genetic Structure and Population History
2.5. Ancestral Area Reconstruction
2.6. Inferring Palaeodrainage along the Coast
3. Results
3.1. Phylogenetic Relationships and Divergence Time
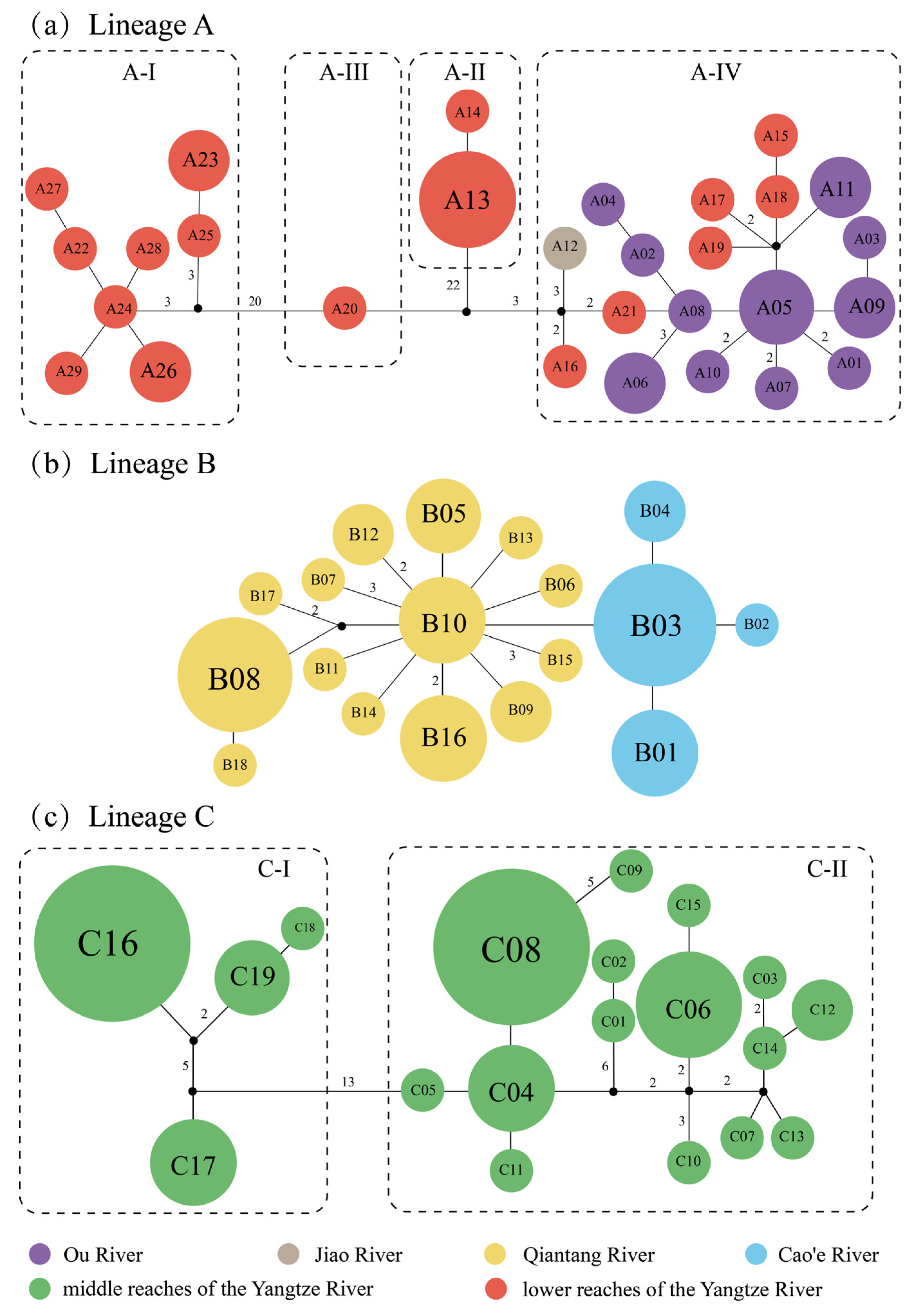
3.2. Genetic Structure
3.3. Demographic History
3.4. Ancestral Area Estimation
3.5. Palaeodrainage Reconstructions
4. Discussion
4.1. Phylogeographic Pattern and Processes
4.2. Genetic Structure and Historical Demography
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Emerson, B.C.; Hewitt, G.M. Phylogeography. Curr. Biol. 2005, 15, R367–R371. [Google Scholar] [CrossRef] [PubMed]
- Avise, J.C. Phylogeography: Retrospect and prospect. J. Biogeogr. 2009, 36, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.M.; Schmidt, D.J.; Finn, D.S. Genes in streams: Using DNA to understand the movement of freshwater fauna and their riverine habitat. Bioscience 2009, 59, 573–583. [Google Scholar] [CrossRef]
- Lyons, N.J.; Val, P.; Albert, J.S.; Willenbring, J.; Gasparini, N.M. Topographic controls on divide migration, stream capture, and diversification in riverine life. Earth Surf. Dyn. 2020, 8, 893–912. [Google Scholar] [CrossRef]
- Lima, S.M.Q.; Berbel, W.; Vilasboa, A.; Lazoski, C.; Volpi, T.; Lazzarotto, H.; Russo, C.A.M.; Tatarenkov, A.; Avise, J.C.; Sole-Cava, A.N.M. Rio de Janeiro and other palaeodrainages evidenced by the genetic structure of an Atlantic Forest catfish. J. Biogeogr. 2021, 48, 1475–1488. [Google Scholar] [CrossRef]
- Barreto, S.B.; Knowles, L.L.; Mascarenhas, R.; Affonso, P.R.A.D.M.; Batalha-Filho, H. Drainage rearrangements and in situ diversification of an endemic freshwater fish genus from north-eastern Brazilian rivers. Freshw. Biol. 2022, 67, 759–773. [Google Scholar] [CrossRef]
- Leigh, D.M.; van Rees, C.B.; Millette, K.L.; Breed, M.F.; Schmidt, C.; Bertola, L.D.; Hand, B.K.; Hunter, M.E.; Jensen, E.L.; Kershaw, F.; et al. Opportunities and challenges of macrogenetic studies. Nat. Rev. Genet. 2021, 22, 791–807. [Google Scholar] [CrossRef]
- Bridle, J.; Hoffmann, A. Understanding the biology of species ranges: When and how does evolution change the rules of ecological engagement? Philos. Trans. R. Soc. B-Biol. Sci. 2022, 337, 20210027. [Google Scholar] [CrossRef]
- Grant, W.S.; Bowen, B.W. Shallow population histories in deep evolutionary lineages of marine fishes: Insights from sardines and anchovies and lessons for conservation. J. Hered. 1998, 89, 415–426. [Google Scholar] [CrossRef]
- Chai, J. Studies on Phylogenetic Relationships, Historical Biogeography and Phylogeography of Squalidus Fishes. Ph.D. Thesis, Fudan University, Shanghai, China, 2020. (In Chinese). [Google Scholar]
- Wu, X.Y.; Luo, J.; Huang, S.; Chen, Z.M.; Xiao, H.; Zhang, Y.P. Molecular Phylogeography and evolutionary history of Poropuntius huangchuchieni (Cyprinidae) in Southwest China. PLoS ONE 2013, 8, e79975. [Google Scholar] [CrossRef] [Green Version]
- Avise, J.C. Phylogeography: The History and Formation of Species; Harvard University Press: Cambridge, MA, USA, 2000; pp. 135–147. ISBN 0-674-66638-0. [Google Scholar]
- Chen, X.L.; Chiang, T.Y.; Lin, H.D.; Zheng, H.; Shao, K.; Zhang, H.S.; Shao, K.T.; Zhang, Q.; Hsu, K.C. Mitochondrial DNA phylogeography of Glyptothorax fokiensis and Glyptothorax hainanensis in Asia. J. Fish Biol. 2007, 70, 75–93. [Google Scholar] [CrossRef]
- Huang, X.X.; Hsu, K.C.; Kang, B.; Kuo, P.H.; Tsai, W.H.; Liang, C.; Lin, H.; Wang, W.K. Population structure of Aphyocypris normalis: Phylogeography and systematics. ZooKeys 2019, 872, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Liang, X.F.; Tang, W.Q.; Zhao, J. Phylogeography of Coreoperca whiteheadi (Perciformes: Coreoperca) in China based on mitochondrial and nuclear gene sequences. Biochem. Syst. Ecol. 2013, 50, 223–231. [Google Scholar] [CrossRef]
- Yang, L.; Mayden, R.L.; He, S.P. Population genetic structure and geographical differentiation of the Chinese catfish Hemibagrus macropterus (Siluriformes, Bagridae): Evidence for altered drainage patterns. Mol. Phylogenet. Evol. 2009, 51, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.Q.; Lin, H.D.; Hsu, K.C.; Kuo, P.H.; Wang, W.K.; Tang, W.Q.; Liu, D.; Yang, J.Q. Spatial genetic structure of the cyprinid fish Onychostoma lepturum on Hainan Island. Mitochondrial DNA A 2017, 28, 901–908. [Google Scholar] [CrossRef]
- Yang, J.Q.; Hsu, K.C.; Liu, Z.Z.; Su, L.W.; Kuo, P.H.; Tang, W.Q.; Zhou, Z.C.; Liu, D.; Bao, B.L.; Lin, H.D. The population history of Garra orientalis (Teleostei: Cyprinidae) using mitochondrial DNA and microsatellite data with approximate Bayesian computation. BMC Evol. Biol. 2016, 16, 73. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.; Armbruster, J.W. Phylogenetic classification of extant genera of fishes of the order Cypriniformes (Teleostei: Ostariophysi). Zootaxa 2018, 4476, 6–39. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.S. Systematics and Biogeography of Microphysogobio Sensu Lato in the Family Gobionidae. Ph.D. Thesis, Fudan University, Shanghai, China, 2021. (In Chinese). [Google Scholar]
- Yue, P.Q. Gobioninae. In Fauna Sinica, Osteichthyes: Cypriniformes (II); Chen, Y.Y., Ed.; Science Press: Beijing, China, 1998; pp. 262–389. ISBN 7-03-005824-0. (In Chinese) [Google Scholar]
- Jiang, Z.G.; Zhang, E. Molecular evidence for taxonomic status of the gudgeon genus Huigobio Fang, 1938 (Teleostei: Cypriniformes), with a description of a new species from Guangdong Province, South China. Zootaxa 2013, 3731, 171–182. [Google Scholar] [CrossRef]
- Sun, Z.X.; Li, X.J.; Tang, W.Q.; Zhao, Y.H. A new species of the gudgeon genus Huigobio Fang, 1938 (Cypriniformes: Cyprinidae) from the Yangtze River Basin, southern China. Zool. Res. 2022, 43, 33–39. [Google Scholar] [CrossRef]
- Mao, J.R. Cypriniformes. In Fauna of Zhejiang-Fresh Water Fishes; Mao, J.R., Xu, S.S., Eds.; Zhejiang Scientific and Technical Press: Hangzhou, China, 1991; pp. 136–137. ISBN 7-5341-0306-1. (In Chinese) [Google Scholar]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef] [Green Version]
- Yin, W. Studies on Phylogeny and Biogeography of the Opsariichthine Fishes. Ph.D. Thesis, Fudan University, Shanghai, China, 2015. [Google Scholar]
- Chai, J.; Fu, C.Z. Three mitochondrial genomes of freshwater fishes in the genus Squalidus (Cypriniformes: Gobionidae). Mitochondrial DNA Part B Resour. 2020, 5, 3761–3763. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, X.H. DAMBE6: New tools for microbial genomics, phylogenetics, and molecular evolution. J. Hered. 2017, 108, 431–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Khnert, D.; Maio, N.D.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.R.; Drummond, A.J. bModelTest: Bayesian phylogenetic site model averaging and model comparison. BMC Evol. Biol. 2017, 17, 42. [Google Scholar] [CrossRef] [Green Version]
- Stout, C.C.; Tan, M.; Lemmon, A.R.; Lemmon, E.M.; Armbruster, J.W. Resolving Cypriniformes relationships using an anchored enrichment approach. BMC Evol. Biol. 2016, 16, 244–257. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhang, X.H.; Mei, X.; Zhao, Q.H.; Guo, X.W.; Zhao, W.N.; Liu, J.X.; Saito, Y.; Wu, Z.Q.; Li, J.; et al. The sedimentary succession of the last ~ 3.50 Myr in the western south Yellow Sea: Paleoenvironmental and tectonic implications. Mar. Geol. 2018, 399, 47–65. [Google Scholar] [CrossRef]
- Zhou, J.J. The Cyprinidae fossils from middle Miocene of Shanwang basin. Vert. PalAsiat. 1990, 28, 95–127. (In Chinese) [Google Scholar]
- Tang, K.L.; Agnew, M.K.; Chen, W.J.; Hirt, M.V.; Raley, M.E.; Sado, T.; Schneider, L.M.; Yang, L.; Bart, H.L.; He, S.P.; et al. Phylogeny of the gudgeons (Teleostei: Cyprinidae: Gobioninae). Mol. Phylogenet. Evol. 2011, 61, 103–124. [Google Scholar] [CrossRef]
- Deng, T.; Hou, S.K.; Wang, S.Q. Neogene integrative stratigraphy and timescale of China. Sci. Chin. Earth Sci. 2019, 62, 310–323. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pons, O.; Petit, R.J. Measuring and testing genetic differentiation with ordered versus unordered alleles. Genetics 1996, 144, 1237–1245. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Dupanloup, I.; Schneider, S.; Excoffier, L. A simulated annealing approach to define the genetic structure of populations. Mol. Ecol. 2002, 11, 2571–2581. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Fu, Y.X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef]
- Ray, N.; Currat, M.; Excoffier, L. Intra-Deme Molecular Diversity in Spatially Expanding Populations. Mol. Biol. Evol. 2003, 20, 76–86. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Matzke, N.J. Probabilistic historical biogeography: New models for founder-event speciation, imperfect detection, and fossils allow improved accuracy and modeltesting. Front. Biogeogr. 2013, 5, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Matzke, N.J. Model Selection in historical biogeography reveals that founder-event speciation is a crucial process in island clades. Syst. Biol. 2014, 63, 951–970. [Google Scholar] [CrossRef] [PubMed]
- Lambeck, K.; Rouby, H.; Purcell, A.; Sun, Y.Y.; Sambridge, M. Sea level and global ice volumes from the Last Glacial Maximum to the Holocene. Proc. Natl. Acad. Sci. USA 2014, 111, 15296–15303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, Y.; Esat, T.M.; Thompson, W.G.; Thomas, A.L.; Webster, J.M.; Miyairi, Y.; Sawada, C.; Aze, T.; Matsuzaki, H.; Okuno, J.; et al. Rapid glaciation and a two-step sea level plunge into the Last Glacial Maximum. Nature 2018, 559, 603–607. [Google Scholar] [CrossRef]
- Thomaz, A.T.; Malabarba, L.R.; Bonatto, S.L.; Knowles, L.L. Testing the effect of palaeodrainages versus habitat stability on genetic divergence in riverine systems: Study of a Neotropical fish of the Brazilian coastal Atlantic Forest. J. Biogeogr. 2015, 42, 2389–2401. [Google Scholar] [CrossRef]
- Gai, C.C.; Liu, Q.S.; Roberts, A.P.; Chou, Y.M.; Zhao, X.X.; Jiang, Z.X.; Liu, J.X. East Asian monsoon evolution since the late Miocene from the South China Sea. Earth Planet. Sci. Lett. 2020, 530, 115960. [Google Scholar] [CrossRef]
- Wang, Y.C.; Lu, H.Y.; Wang, K.X.; Wang, Y.; Li, Y.X.; Clemens, S.; Lv, H.Z.; Huang, Z.H.; Wang, H.L.; Hu, X.Z.; et al. Combined high- and low-latitude forcing of East Asian monsoon precipitation variability in the Pliocene warm period. Sci. Adv. 2020, 6, eaba2414. [Google Scholar] [CrossRef]
- Lu, J.Y.; Yang, H.; Griffiths, M.L.; Burls, N.; Xiao, G.Q.; Yang, J.L.; Wang, J.K.; Johnson, K.R.; Xie, S.C. Asian monsoon evolution linked to Pacific temperature gradients since the Late Miocene. Earth Planet. Sci. Lett. 2021, 563, 116882. [Google Scholar] [CrossRef]
- Yang, S.L.; Ding, Z.L.; Feng, S.H.; Jiang, W.Y.; Huang, X.F.; Guo, L.C. A strengthened East Asian summer monsoon during Pliocene warmth: Evidence from ‘red clay’ sediments at Pianguan, northern China. J. Asian Earth Sci. 2018, 155, 124–133. [Google Scholar] [CrossRef]
- Huang, X.F.; Jiang, D.B.; Dong, X.X.; Yang, S.L.; Su, B.H.; Li, X.Y.; Tang, Z.H.; Wang, Y.D. Northwestward migration of the northern edge of the East Asian Summer Monsoon during the Mid-Pliocene warm period: Simulations and reconstructions. J. Geophys. Res. Atmos. 2019, 124, 1392–1404. [Google Scholar] [CrossRef]
- Abell, J.T.; Winckler, G.; Anderson, R.F.; Herbert, T.D. Poleward and weakened westerlies during Pliocene warmth. Nature 2021, 589, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.G.; Browning, J.V.; Schmelz, W.J.; Kopp, R.E.; Mountain, G.S.; Wright, J.D. Cenozoic sea-level and cryospheric evolution from deep-sea geochemical and continental margin records. Sci. Adv. 2020, 6, eaaz1346. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.B.; Pan, B.T.; Ma, J.F.; Hu, X.F.; Geng, H.P.; Pang, H.L. Paleoclimatic record from the clay mineralogy of Quaternary sediments of drill core DWJ from Jiudong subbasin (western Hexi Corridor), NW China. Quat. Res. 2020, 93, 124–138. [Google Scholar] [CrossRef]
- Cheng, H.; Edwards, R.L.; Sinha, A.; Spötl, C.; Li, Y.; Chen, S.T.; Kelly, M.; Kathayat, G.; Wang, X.F.; Li, X.L.; et al. The Asian monsoon over the past 640,000 years and ice age terminations. Nature 2016, 534, 640–646. [Google Scholar] [CrossRef]
- Wang, L.Y.; Li, G.X.; Liu, J.; Mei, X.; Zhang, Y. Astronomical dating of Quaternary strata in the South Yellow Sea and its indication for paleoclimatic evolution. Mar. Geol. 2021, 439, 106557. [Google Scholar] [CrossRef]
- Araripe, J.; do Rêgo, P.S.; Queiroz, H.; Sampaio, I.; Schneider, H. Dispersal capacity and genetic structure of Arapaima gigas on different geographic scales using microsatellite markers. PLoS ONE 2013, 8, e54470. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.T.; Zhong, Z.X.; Dai, W.; Fan, Q.; He, S.P. Phylogeographic structure, cryptic speciation and demographic history of the sharpbelly (Hemiculter leucisculus), a freshwater habitat generalist from Southern China. BMC Evol. Biol. 2017, 17, 216. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.Y.; Nie, J.S.; Li, Z.J.; Qiao, Q.Q.; Abell, J.T.; Wang, F.; Xiao, W.J. Eccentricity forcing of East Asian monsoonal systems over the past 3 million years. Proc. Natl. Acad. Sci. USA 2021, 118, e2107055118. [Google Scholar] [CrossRef]
- Yang, L.; He, S.P. Phylogeography of the freshwater catfish Hemibagrus guttatus (Siluriformes, Bagridae): Implications for South China biogeography and influence of sea level changes. Mol. Phylogenet. Evol. 2008, 49, 393–398. [Google Scholar] [CrossRef]
- Šlechtová, V.; Bohlen, J.; Freyhof, J.; Persat, H.; Delmastro, G.B. The Alps as barrier to dispersal in cold-adapted freshwater fishes? Phylogeographic history and taxonomic status of the bullhead in the Adriatic freshwater drainage. Mol. Phylogenet. Evol. 2004, 33, 225–239. [Google Scholar] [CrossRef]
- Barluenga, M.; Meyer, A. Old fish in a young lake: Stone loach (Pisces: Barbatula barbatula) populations in Lake Constance are genetically isolated by distance. Mol. Ecol. 2005, 14, 1229–1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, T.Y.; Lin, H.D.; Zhao, J.; Kuo, P.H.; Lee, T.W.; Hsu, K.C. Diverse processes shape deep phylogeographical divergence in Cobitis sinensis (Teleostei: Cobitidae) in East Asia. J. Zool. Syst. Evol. Res. 2013, 51, 316–326. [Google Scholar] [CrossRef]
- Li, G.G.; Peng, Z.G.; Zhang, R.Y.; Tang, Y.T.; Tong, C.; Feng, C.G.; Zhang, C.F.; Zhao, K. Mito-nuclear phylogeography of the cyprinid fish Gymnodiptychus dybowskii in the arid Tien Shan region of Central Asia. Biol. J. Linnean Soc. 2016, 118, 304–314. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.T.; Li, C.; Chen, F.C.; Li, Y.F.; Yang, J.P.; Li, J.; Li, X.H. Phylogeographic analyses of a migratory freshwater fish (Megalobrama terminalis) reveal a shallow genetic structure and pronounced effects of sea-level changes. Gene 2020, 737, 144478. [Google Scholar] [CrossRef] [PubMed]
- Grant, K.M.; Rohling, E.J.; Ramsey, C.B.; Cheng, H.; Edwards, R.L.; Florindo, F.; Heslop, D.; Marra, F.; Roberts, A.P.; Tamisiea, M.E.; et al. Sea-level variability over five glacial cycles. Nat. Commun. 2014, 5, 5076. [Google Scholar] [CrossRef]
- Zhao, L.J.; Zhou, X.Y.; Liu, Q.G.; Zhang, H. Genetic variation and phylogeography of Sinibrama macrops (Teleostei: Cyprinidae) in Qiantang River basin, China. Biochem. Syst. Ecol. 2013, 49, 10–20. [Google Scholar] [CrossRef]
- Chen, W.T.; Du, K.; He, S.P. Genetic structure and historical demography of Schizothorax nukiangensis (Cyprinidae) in continuous habitat. Ecol. Evol. 2015, 5, 984–995. [Google Scholar] [CrossRef]
- Zheng, L.P.; Yang, J.X. Genetic diversity and population demography of the endemic species Acrossocheilus longipinnis (Teleostei, Cyprinidae) based on mtDNA COI and Cytb gene sequences. Mitochondrial DNA Part A 2018, 29, 403–408. [Google Scholar] [CrossRef]
- Zhao, J.; Hsu, K.C.; Luo, J.Z.; Wang, C.H.; Chan, B.P.; Li, J.; Kuo, P.H.; Lin, H.D. Genetic diversity and population history of Tanichthys albonubes (Teleostei: Cyprinidae): Implications for conservation. Aquat. Conserv.-Mar. Freshw. Ecosyst. 2018, 28, 422–434. [Google Scholar] [CrossRef]
- Zhang, W.J.; Wang, J.J.; Li, C.; Chen, J.Q.; Li, W.; Jiang, S.Y.; Hsu, K.C.; Zhao, M.T.; Lin, H.D.; Zhao, J. Spatial genetic structure of Opsariichthys hainanensis in South China. Mitochondrial DNA Part A 2020, 31, 98–107. [Google Scholar] [CrossRef]
- Chen, A.H. Studies on Molecular Phylogeny of the Gobioninae (Teleostei: Cyprinidae). Master Dissertation, Fudan University, Shanghai, China, 2014. [Google Scholar]
- Kim, P.; Han, J.H.; An, S.L. Complete mitochondrial genome of freshwater fish, Microphysogobio jeoni (Actinopterygii, Cypriniformes, Cyprinidae), from South Korea. Mitochondrial DNA Part B-Resour. 2020, 5, 856–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.Y.; Wang, J.P.; Ho, C.W.; Cheng, J.W.; Ueng, Y.T. Complete mitochondrial DNA genome of Microphysogobio brevirostris (Cypriniformes: Cyprinidae). Mitochondrial DNA 2015, 26, 293–294. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.S.; Song, H.B.; Lee, J.S. Complete mitochondrial genome of the freshwater gudgeon, Microphysogobio koreensis (Cypriniformes, Cyprinidae). Mitochondrial DNA 2014, 25, 15–16. [Google Scholar] [CrossRef]
- Kim, K.R.; Bang, I.C. Complete mitochondrial genome of the endangered freshwater fish Microphysogobio rapidus (Cypriniformes, Cyprinidae) from Korea. Mitochondrial DNA Part B-Resour. 2020, 5, 447–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.Y.; Lin, H.D.; Tzeng, S.J.; Chiang, T.Y. Complete mitochondrial genome of Microphysogobio alticorpus (Cypriniformes, Cyprinidae). Mitochondrial DNA 2014, 25, 173–174. [Google Scholar] [CrossRef]
- Horikawa, M.; Nakajima, J.; Mukai, T. Distribution of indigenous and non-indigenous mtDNA haplotypes of Biwia zezera (Cyprinidae) in northern Kyushu, Japan. Jpn. J. Ichthyol. 2007, 54, 149–159. (In Japanese) [Google Scholar]
- Hwang, D.S.; Byeon, H.K.; Lee, J.S. Complete mitochondrial genome of the freshwater gudgeon, Gobiobotia macrocephala (Cypriniformes; Cyprinidae). Mitochondrial DNA 2014, 25, 31–32. [Google Scholar] [CrossRef]
- Li, Y.H.; Cao, K.; Fu, C.Z. Ten fish mitogenomes of the tribe Gobionini (Cypriniformes: Cyprinidae: Gobioninae). Mitochondrial DNA Part B-Resour. 2018, 3, 802–803. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.Y.; Jean, C.T.; Chen, Y.J.; Lin, K.Y.; Liu, M.Y. The complete mitochondrial genome sequence of Belligobio nummifer (Cypriniformes, Cyprinidae). Mitochondrial DNA 2016, 27, 435–436. [Google Scholar] [CrossRef]
- Kim, K.Y.; Lim, Y.H.; Bang, I.C.; Nam, Y.K. Phylogenetic relationships among three new Hemibarbus mitogenome sequences belonging to the subfamily Gobioninae (Teleostei, Cypriniformes, and Cyprinidae). Mitochondrial DNA 2009, 20, 119–125. [Google Scholar] [CrossRef]
- Chen, Y.; Fu, C.Z. Three complete mitochondrial genomes of freshwater fishes in the genus Abbottina (Cypriniformes: Gobionidae). Mitochondrial DNA Part B-Resour. 2019, 4, 2179–2180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.W.; Fu, C.Z. Three mitochondrial genomes of Pseudogobio fishes (Cypriniformes: Gobionidae). Mitochondrial DNA Part B-Resour. 2020, 5, 3046–3047. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Fu, C.Z. Four complete mitochondrial genomes of Saurogobio fishes (Cypriniformes: Gobionidae). Mitochondrial DNA Part B-Resour. 2019, 4, 2175–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, T.Y.; Fu, C.Z. Two mitochondrial genomes of freshwater gudgeons in the genus Gobio (Cypriniformes: Gobionidae). Mitochondrial DNA Part B-Resour. 2020, 5, 3054–3055. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, K.; Sado, T.; Mayden, R.L.; Hanzawa, N.; Nakamura, K.; Nishida, M.; Miya, M. Mitogenomic evolution and interrelationships of the Cypriniformes (Actinopterygii: Ostariophysi): The first evidence toward resolution of higher-level relationships of the world’s largest freshwater fish clade based on 59 whole mitogenome sequences. J. Mol. Evol. 2006, 63, 826–841. [Google Scholar] [CrossRef]
- Tao, W.J.; Zhao, H.P. The complete mitogenome of Gnathopogon polytaenia (Cypriniformes; Cyprinidae). Mitochondrial DNA 2016, 27, 1307–1308. [Google Scholar] [CrossRef]
- Kim, K.Y.; Nam, Y.K.; Bang, I. Phylogeny and divergence time estimation of Coreoleuciscus splendidus populations (Teleostei: Cypriniformes) endemic to Korea based on complete mitogenome sequences. Genes Genom. 2012, 34, 149–156. [Google Scholar] [CrossRef]
- Chen, A.H.; Xia, R.; Lei, G.C.; Fu, C.Z. Complete mitochondrial genome of Pseudorasbora elongata (Cypriniformes: Cyprinidae). Mitochondrial DNA 2015, 26, 250–251. [Google Scholar] [CrossRef]
- Yu, J.N.; Kim, S.; Kwak, M. Complete mitochondrial genome sequence of a Korean Pungtungia herzi (Cypriniformes, Gobioninae). Mitochondrial DNA 2014, 25, 414–415. [Google Scholar] [CrossRef]
- Hwang, D.S.; Lee, W.K.; Lee, J.S. Complete mitochondrial genome of the freshwater gudgeon, Pseudopungtungia nigra (Cypriniformes, Gobioninae). Mitochondrial DNA 2014, 25, 1–2. [Google Scholar] [CrossRef]
- Chen, X.; Zhou, Z.M.; Chen, Z.J.; Ai, W.M. Complete mitochondrial genome of Sarcocheilichthys parvus (Cypriniformes, Cyprinidae). Mitochondrial DNA 2013, 24, 97–98. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.R.; Lin, H.D.; Su, L.W.; Tang, W.Q.; Wu, C.Y.; Yang, J.Q. The complete mitochondrial genome sequence of Sarcocheilichthys nigripinnis (Cypriniformes, Cyprinidae). Mitochondrial DNA 2013, 24, 478–480. [Google Scholar] [CrossRef] [PubMed]
- Du, F.K.; Xu, G.C.; Nie, Z.J.; Xu, P.; Gu, R.B. Complete mitochondrial genome of Paracanthobrama guichenoti. Mitochondrial DNA 2016, 27, 727–728. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.D.; Li, P.; Zhang, Y.G.; Peng, Z.G. Comparative study of the complete mitochondrial genomes of the bronze gudgeon (Coreius heterodon) and largemouth bronze gudgeon (Coreius guichenoti). Mitochondrial DNA 2013, 24, 189–190. [Google Scholar] [CrossRef] [PubMed]
- An, Q.; Fu, C.Z. Complete mitochondrial genome of Ladislavia taczanowskii (Cypriniformes: Gobionidae). Mitochondrial DNA Part B-Resour. 2021, 6, 2227–2228. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.P.; Ma, Z.H.; Yang, X.F.; Xu, H.X.; Yang, R.B. Complete mitochondrial genome of the Chinese bitterling Acheilognathus macropterus (Cypriniformes: Cyprinidae). Mitochondrial DNA Part A 2016, 27, 589–590. [Google Scholar] [CrossRef]
- Xu, C.Z.; Xie, F.; Zhang, X.; Zhao, S. The complete mitochondrial genome of Amur bitterling (Rhodeus sericeus) from China. Mitochondrial DNA Part A 2016, 27, 2377–2378. [Google Scholar] [CrossRef]
- Luo, Y.; Cao, X.J.; Zhu, Y.R. The complete mitochondrial genome of Tanakia limbata (Cypriniformes: Cyprinidae). Mitochondrial DNA 2016, 27, 1713–1714. [Google Scholar] [CrossRef]
- Song, H.Y.; Bang, I.C. Coreoleuciscus aeruginos (Teleostei: Cypriniformes: Cyprinidae), a new species from the Seomjin and Nakdong rivers, Korea. Zootaxa 2015, 3931, 140–150. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Basin and Locality | Lat. | Long. | N | Cytb Haplotype |
|---|---|---|---|---|
| Ou River | ||||
| 1. Yongjia County, Zhejiang Province | 28.35° N | 120.75° E | 14 | A01(1), A02(1), A03(1), A04(1), A05(3), A06(2), A07(1), A08(1), A09(2), A10(1) |
| 2. Jinyun County, Zhejiang Province | 28.66° N | 120.09° E | 2 | A11(2) |
| Jiao River | ||||
| 3. Tiantai County, Zhejiang Province | 29.14° N | 121.03° E | 1 | A12(1) |
| Cao’e River | ||||
| 4. Xinchang County, Zhejiang Province | 29.51° N | 120.91° E | 15 | B01(4), B02(1), B03(8), B04(2) |
| Qiantang River | ||||
| 5. Pan’an County, Zhejiang Province | 29.06° N | 120.45° E | 1 | B05(1) |
| 6. Yuqian Town, Lin’an City, Zhejiang Province | 30.19° N | 119.40° E | 6 | B06(2), B07(1), B08(2), B09(1) |
| 7. Changhua Town, Lin’an City, Zhejiang Province | 30.17° N | 119.22° E | 10 | B08(3), B10(4), B11(1), B12(1), B13(1) |
| 8. Yunti Village, Ningguo City, Anhui Province | 30.37° N | 119.21° E | 11 | B05(1), B08(2), B12(1), B14(1), B15(1), B16(4), B17(1) |
| 9. Huangshan City, Anhui Province | 29.72° N | 118.31° E | 1 | B05(1) |
| 10. Jiangshan City, Zhejiang Province | 28.76° N | 118.66° E | 1 | B18(1) |
| Yangtze River | ||||
| 11. Anji County, Zhejiang Province | 30.64° N | 119.67° E | 6 | A13(5), A14(1) |
| 12. Ningguo City, Anhui Province | 30.62° N | 118.95° E | 7 | A15(1), A16(1), A17(1), A18(1), A19(1), A20(1), A21(1) |
| 13. Shitai County, Anhui Province | 30.21° N | 117.50° E | 10 | A22(1), A23(2), A24(1), A25(1), A26(2), A27(1), A28(1), A29(1) |
| 14. Fuliang County, Jiangxi Province | 29.52° N | 117.28° E | 1 | C01(1) |
| 15. Yushan County, Jiangxi Province | 28.69° N | 118.28° E | 1 | C02(1) |
| 16. Fengxin County, Jiangxi Province | 28.71° N | 115.38° E | 6 | C03(1), C04(2), C05(1), C06(1), C07(1) |
| 17. Jing’an County, Jiangxi Province | 28.85° N | 115.39° E | 18 | C04(2), C08(13), C09(1), C10(1), C11(1) |
| 18. Tonggu County, Jiangxi Province | 28.53° N | 114.39° E | 9 | C06(5), C12(2), C13(1), C14(1) |
| 19. Xiushui County, Jiangxi Province | 29.02° N | 114.54° E | 1 | C15(1) |
| 20. Ningdu County, Jiangxi Province | 26.46° N | 116.00° E | 5 | C16(3), C17(2) |
| 21. Suichuan County, Jiangxi Province | 26.31° N | 114.52° E | 15 | C16(9), C17(2), C18(1), C19(3) |
| 22. Anfu County, Jiangxi Province | 27.39° N | 114.63° E | 1 | C16(1) |
| Basin | Number | Number of HaploTypes (Nh) | Number of Private Haplotypes (Nph) | Haplotype Diversity (h) | Nucleotide Diverstiy (π) |
|---|---|---|---|---|---|
| Ou River | 16 | 11 | 11 | 0.9500 ± 0.0364 | 0.0028 ± 0.0017 |
| Jiao River | 1 | 1 | 1 | - | - |
| Cao’e River | 15 | 4 | 4 | 0.6667 ± 0.0991 | 0.0007 ± 0.0006 |
| Qiantang River | 30 | 14 | 14 | 0.9126 ± 0.0307 | 0.0024 ± 0.0015 |
| Yangtze River | 80 | 36 | 36 | 0.9370 ± 0.0149 | 0.0371 ± 0.0180 |
| Total | 142 | 66 | 66 | 0.9722 ± 0.0054 | 0.0460 ± 0.0222 |
| Ou River | Cao’e River | Qiantang River | Yangtze River | |
|---|---|---|---|---|
| Ou River | 0.000 | 0.000 | 0.000 | |
| Cao’e River | 0.986 | 0.000 | 0.000 | |
| Qiantang River | 0.981 | 0.367 | 0.000 | |
| Yangtze River | 0.546 | 0.510 | 0.559 |
| Lineage | Tajima’s D | Fu’s Fs |
|---|---|---|
| A | 0.320 (p = 0.706) | −4.231 (p = 0.095) |
| B | −1.768 (p = 0.014) | −9.126 (p = 0.000) |
| C | 0.528 (p = 0.781) | 1.220 (p = 0.714) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Ni, X.; Fu, C. Phylogeographical Analysis of the Freshwater Gudgeon Huigobio chenhsienensis (Cypriniformes: Gobionidae) in Southern China. Life 2022, 12, 1024. https://doi.org/10.3390/life12071024
Yang X, Ni X, Fu C. Phylogeographical Analysis of the Freshwater Gudgeon Huigobio chenhsienensis (Cypriniformes: Gobionidae) in Southern China. Life. 2022; 12(7):1024. https://doi.org/10.3390/life12071024
Chicago/Turabian StyleYang, Xishu, Xiaomin Ni, and Cuizhang Fu. 2022. "Phylogeographical Analysis of the Freshwater Gudgeon Huigobio chenhsienensis (Cypriniformes: Gobionidae) in Southern China" Life 12, no. 7: 1024. https://doi.org/10.3390/life12071024