
Caffeine Decreases Hepcidin Expression to Alleviate Aberrant Iron Metabolism under Inflammation by Regulating the IL-6/STAT3 Pathway


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Quantitative Real-Time PCR (qPCR)
2.3. Western Blot Analysis
2.4. Inductively Coupled Plasma Mass Spectrometry (ICP–MS)
2.5. Measurement of Serum Iron
2.6. Enzyme Linked Immunosorbent Assay (ELISA)
2.7. Statistical Analysis
3. Results
3.1. LPS-Induced Elevated Hamp Expression Was Alleviated by Caffeine
3.2. Caffeine Suppressed LPS-Induced Upregulated Il6 Expression and STAT3 Phosphorylation
3.3. Caffeine Decreased Iron Levels and Attenuated LPS-Mediated Iron Accumulation in the Liver
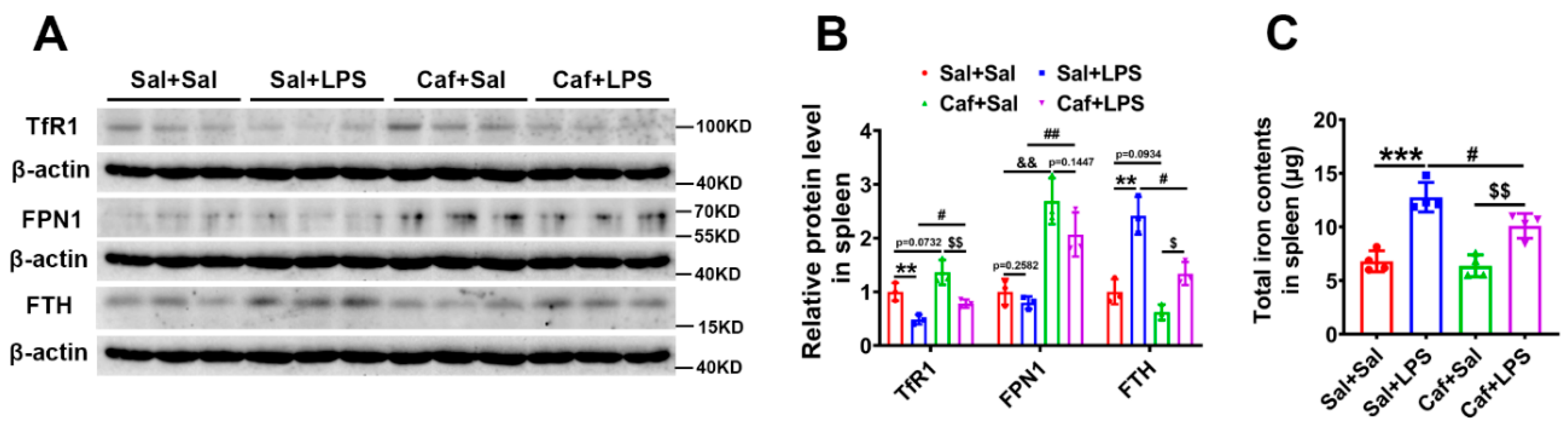
3.4. Caffeine Reduced Iron Content in the Spleen after LPS Treatment
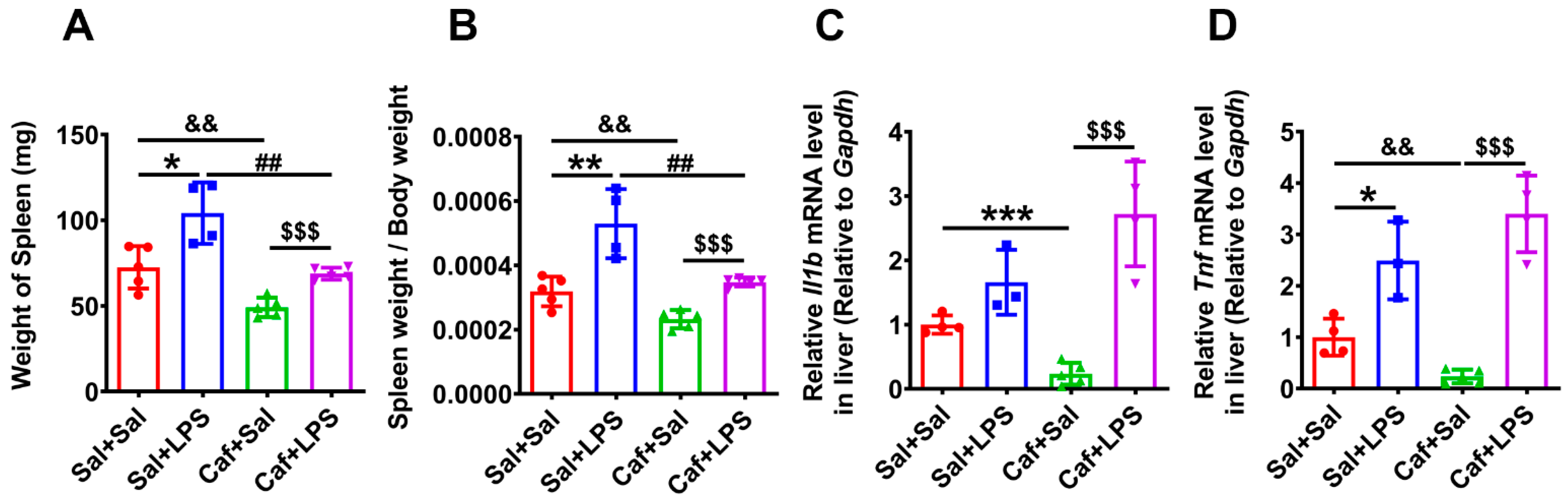
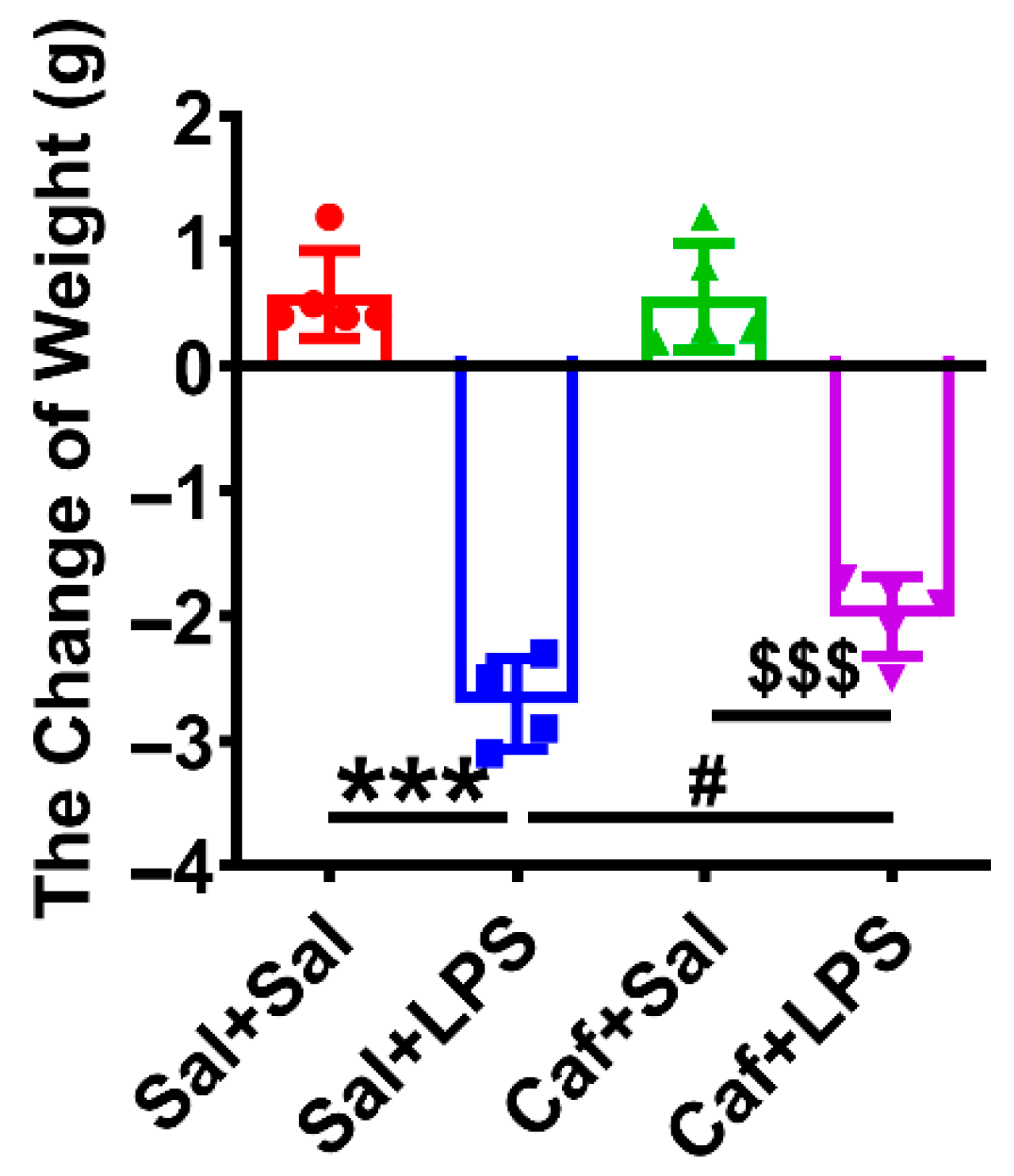
3.5. Caffeine Alleviated LPS-Induced Splenomegaly and Weight Loss
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Poole, R.; Kennedy, O.J.; Roderick, P.; Fallowfield, J.A.; Hayes, P.C.; Parkes, J. Coffee Consumption and Health: Umbrella Review of Meta-Analyses of Multiple Health Outcomes. BMJ 2017, 359, j5024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebeau, P.F.; Byun, J.H.; Platko, K.; Saliba, P.; Sguazzin, M.; MacDonald, M.E.; Paré, G.; Steinberg, G.R.; Janssen, L.J.; Igdoura, S.A.; et al. Caffeine Blocks SREBP2-Induced Hepatic PCSK9 Expression to Enhance LDLR-Mediated Cholesterol Clearance. Nat. Commun. 2022, 13, 770. [Google Scholar] [CrossRef]
- O’Keefe, J.H.; Bhatti, S.K.; Patil, H.R.; Dinicolantonio, J.J.; Lucan, S.C.; Lavie, C.J. Effects of Habitual Coffee Consumption on Cardiometabolic Disease, Cardiovascular Health, and All-Cause Mortality. J. Am. Coll. Cardiol. 2013, 62, 1043–1051. [Google Scholar] [CrossRef] [Green Version]
- Crielaard, B.J.; Lammers, T.; Rivella, S. Targeting Iron Metabolism in Drug Discovery and Delivery. Nat. Rev. Drug Discov. 2017, 16, 400–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zecca, L.; Youdim, M.B.H.; Riederer, P.; Connor, J.R.; Crichton, R.R. Iron, Brain Ageing and Neurodegenerative Disorders. Nat. Rev. Neurosci. 2004, 5, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Camaschella, C. Iron-Deficiency Anemia. N. Engl. J. Med. 2015, 373, 485–486. [Google Scholar] [CrossRef] [Green Version]
- Pietrangelo, A. Iron and the Liver. Liver Int. 2016, 36, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Stockwell, B.R.; Friedmann Angeli, J.P.; Bayir, H.; Bush, A.I.; Conrad, M.; Dixon, S.J.; Fulda, S.; Gascón, S.; Hatzios, S.K.; Kagan, V.E.; et al. Ferroptosis: A Regulated Cell Death Nexus Linking Metabolism, Redox Biology, and Disease. Cell 2017, 171, 273–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muckenthaler, M.U.; Rivella, S.; Hentze, M.W.; Galy, B. A Red Carpet for Iron Metabolism. Cell 2017, 168, 344–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraenkel, P.G. Anemia of Inflammation: A Review. Med. Clin. N. Am. 2017, 101, 285–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardenghi, S.; Renaud, T.M.; Meloni, A.; Casu, C.; Crielaard, B.J.; Bystrom, L.M.; Greenberg-Kushnir, N.; Sasu, B.J.; Cooke, K.S.; Rivella, S. Distinct Roles for Hepcidin and Interleukin-6 in the Recovery from Anemia in Mice Injected with Heat-Killed Brucella Abortus. Blood 2014, 123, 1137–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, G.; Ganz, T.; Goodnough, L.T. Anemia of Inflammation. Blood 2014, 28, 671–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; He, S.; Li, R.; Zhou, X.; Zhang, S.; Yu, M.; Ye, Y.; Wang, Y.; Huang, C.; Wu, M. Pseudomonas Aeruginosa Infection Augments Inflammation through MIR-301b Repression of c-Myb-Mediated Immune Activation and Infiltration. Nat. Microbiol. 2016, 1, 16132. [Google Scholar] [CrossRef] [Green Version]
- de Alcântara Almeida, I.; Mancebo Dorvigny, B.; Souza Tavares, L.; Nunes Santana, L.; Vitor Lima-Filho, J. Anti-Inflammatory Activity of Caffeine (1,3,7-Trimethylxanthine) after Experimental Challenge with Virulent Listeria Monocytogenes in Swiss Mice. Int. Immunopharmacol. 2021, 100, 108090. [Google Scholar] [CrossRef] [PubMed]
- Weichelt, U.; Cay, R.; Schmitz, T.; Strauss, E.; Sifringer, M.; Bührer, C.; Endesfelder, S. Prevention of Hyperoxia-Mediated Pulmonary Inflammation in Neonatal Rats by Caffeine. Eur. Respir. J. 2013, 41, 966–973. [Google Scholar] [CrossRef] [Green Version]
- Kupfer, G.M.; D’Andrea, A.D. The Effect of the Fanconi Anemia Polypeptide, FAC, upon P53 Induction and G2 Checkpoint Regulation. Blood 1996, 88, 1019–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Floride, E.; Föller, M.; Ritter, M.; Lang, F. Caffeine Inhibits Suicidal Erythrocyte Death. Cell. Physiol. Biochem. 2008, 22, 253–260. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, Y.; Zhang, J.H.; Han, K.; Zhang, X.; Bai, X.; You, L.H.; Yu, P.; Shi, Z.; Chang, Y.Z.; et al. Astrocyte Hepcidin Ameliorates Neuronal Loss through Attenuating Brain Iron Deposition and Oxidative Stress in APP/PS1 Mice. Free Radic. Biol. Med. 2020, 158, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wu, Q.; Wu, W.; Li, H.; Guo, Y.; Yu, P.; Gao, G.; Shi, Z.; Zhao, B.; Chang, Y.Z. Mitochondrial Ferritin Deletion Exacerbates β -Amyloid-Induced Neurotoxicity in Mice. Oxid. Med. Cell. Longev. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.-H.; Nie, G.; Duan, X.-L.; Rouault, T.; Wu, W.-S.; Ning, B.; Zhang, N.; Chang, Y.-Z.; Zhao, B.-L. Neuroprotective Mechanism of Mitochondrial Ferritin on 6-Hydroxydopamine–Induced Dopaminergic Cell Damage: Implication for Neuroprotection in Parkinson’s Disease. Antioxid. Redox Signal. 2010, 13, 783–796. [Google Scholar] [CrossRef]
- Lei, P.; Ayton, S.; Finkelstein, D.I.; Spoerri, L.; Ciccotosto, G.D.; Wright, D.K.; Wong, B.X.W.; Adlard, P.A.; Cherny, R.A.; Lam, L.Q.; et al. Tau Deficiency Induces Parkinsonism with Dementia by Impairing APP-Mediated Iron Export. Nat. Med. 2012, 18, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Liu, S.Y.; Wang, H.J.; Zhang, T.W.; Yu, P.; Duan, X.L.; Zhao, S.E.; Chang, Y.Z. Effects of Pregnancy and Lactation on Iron Metabolism in Rats. Biomed Res. Int. 2015, 2015, 105325. [Google Scholar] [CrossRef] [PubMed]
- Verga Falzacappa, M.V.; Spasic, M.V.; Kessler, R.; Stolte, J.; Hentze, M.W.; Muckenthaler, M.U. STAT3 Mediates Hepatic Hepcidin Expression and Its Inflammatory Stimulation. Blood 2007, 109, 353–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knutson, M.D.; Oukka, M.; Koss, L.M.; Aydemir, F.; Wessling-Resnick, M. Iron Release from Macrophages after Erythrophagocytosis Is Up-Regulated by Ferroportin 1 Overexpression and down-Regulated by Hepcidin. Proc. Natl. Acad. Sci. USA 2005, 102, 1324–1328. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.L.; Senecal, T.; Ghosh, M.C.; Ollivierre-Wilson, H.; Tu, T.; Rouault, T.A. Hepcidin Regulates Ferroportin Expression and Intracellular Iron Homeostasis of Erythroblasts. Blood 2011, 118, 2868–2877. [Google Scholar] [CrossRef] [Green Version]
- Rouault, T.A. Iron Metabolism in the CNS: Implications for Neurodegenerative Diseases. Nat. Rev. Neurosci. 2013, 14, 551–564. [Google Scholar] [CrossRef]
- Hentze, M.W.; Muckenthaler, M.U.; Galy, B.; Camaschella, C. Two to Tango: Regulation of Mammalian Iron Metabolism. Cell 2010, 142, 24–38. [Google Scholar] [CrossRef] [Green Version]
- You, L.H.; Yan, C.Z.; Zheng, B.J.; Ci, Y.Z.; Chang, S.Y.; Yu, P.; Gao, G.F.; Li, H.Y.; Dong, T.Y.; Chang, Y.Z. Astrocyte Hepcidin Is a Key Factor in LPS-Induced Neuronal Apoptosis. Cell Death Dis. 2017, 8, e2676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaacs, J.D.; Harari, O.; Kobold, U.; Lee, J.S.; Bernasconi, C. Effect of Tocilizumab on Haematological Markers Implicates Interleukin-6 Signalling in the Anaemia of Rheumatoid Arthritis. Arthritis Res. Ther. 2013, 15, R204. [Google Scholar] [CrossRef] [Green Version]
- Poli, M.; Girelli, D.; Campostrini, N.; Maccarinelli, F.; Finazzi, D.; Luscieti, S.; Nai, A.; Arosio, P. Heparin: A Potent Inhibitor of Hepcidin Expression in vitro and in vivo. Blood 2011, 117, 997–1004. [Google Scholar] [CrossRef]
- Song, S.N.J.; Tomosugi, N.; Kawabata, H.; Ishikawa, T.; Nishikawa, T.; Yoshizaki, K. Down-Regulation of Hepcidin Resulting from Long-Term Treatment with an Anti-IL-6 Receptor Antibody (Tocilizumab) Improves Anemia of Inflammation in Multicentric Castleman Disease. Blood 2010, 116, 3627–3634. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| Gapdh | 5′-TGCACCACCAACTGCTTAGC-3′ | 5′-GGCATGGACTGTGGTCATGAG-3′ |
| Hamp | 5′-AGACATTGCGATACCAATGCA-3′ | 5′-GCAACAGATACCACACTGGGAA-3′ |
| Il6 | 5′-ACCGCTATGAAGTTCCTCTC-3′ | 5′-CTCTGTGAAGTCTCCTCTCC-3′ |
| Il1b | 5′-CCAGCAGGTTATCATCATCATCC-3′ | 5′-CTCGCAGCAGCACATCAAC-3′ |
| Tnf | 5′-TACTGAACTTCGGGGTGATTGGTCC-3′ | 5′-CAGCCTTGTCCCTTGAAGAGAACC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.-D.; Geng, M.-Y.; Dou, S.-R.; Wang, X.; Zhang, Z.-H.; Chang, Y.-Z. Caffeine Decreases Hepcidin Expression to Alleviate Aberrant Iron Metabolism under Inflammation by Regulating the IL-6/STAT3 Pathway. Life 2022, 12, 1025. https://doi.org/10.3390/life12071025
Li Z-D, Geng M-Y, Dou S-R, Wang X, Zhang Z-H, Chang Y-Z. Caffeine Decreases Hepcidin Expression to Alleviate Aberrant Iron Metabolism under Inflammation by Regulating the IL-6/STAT3 Pathway. Life. 2022; 12(7):1025. https://doi.org/10.3390/life12071025
Chicago/Turabian StyleLi, Zhong-Da, Meng-Yu Geng, Song-Rui Dou, Xuan Wang, Zi-Han Zhang, and Yan-Zhong Chang. 2022. "Caffeine Decreases Hepcidin Expression to Alleviate Aberrant Iron Metabolism under Inflammation by Regulating the IL-6/STAT3 Pathway" Life 12, no. 7: 1025. https://doi.org/10.3390/life12071025