
cAMP Is a Promising Regulatory Molecule for Plant Adaptation to Heat Stress

Abstract
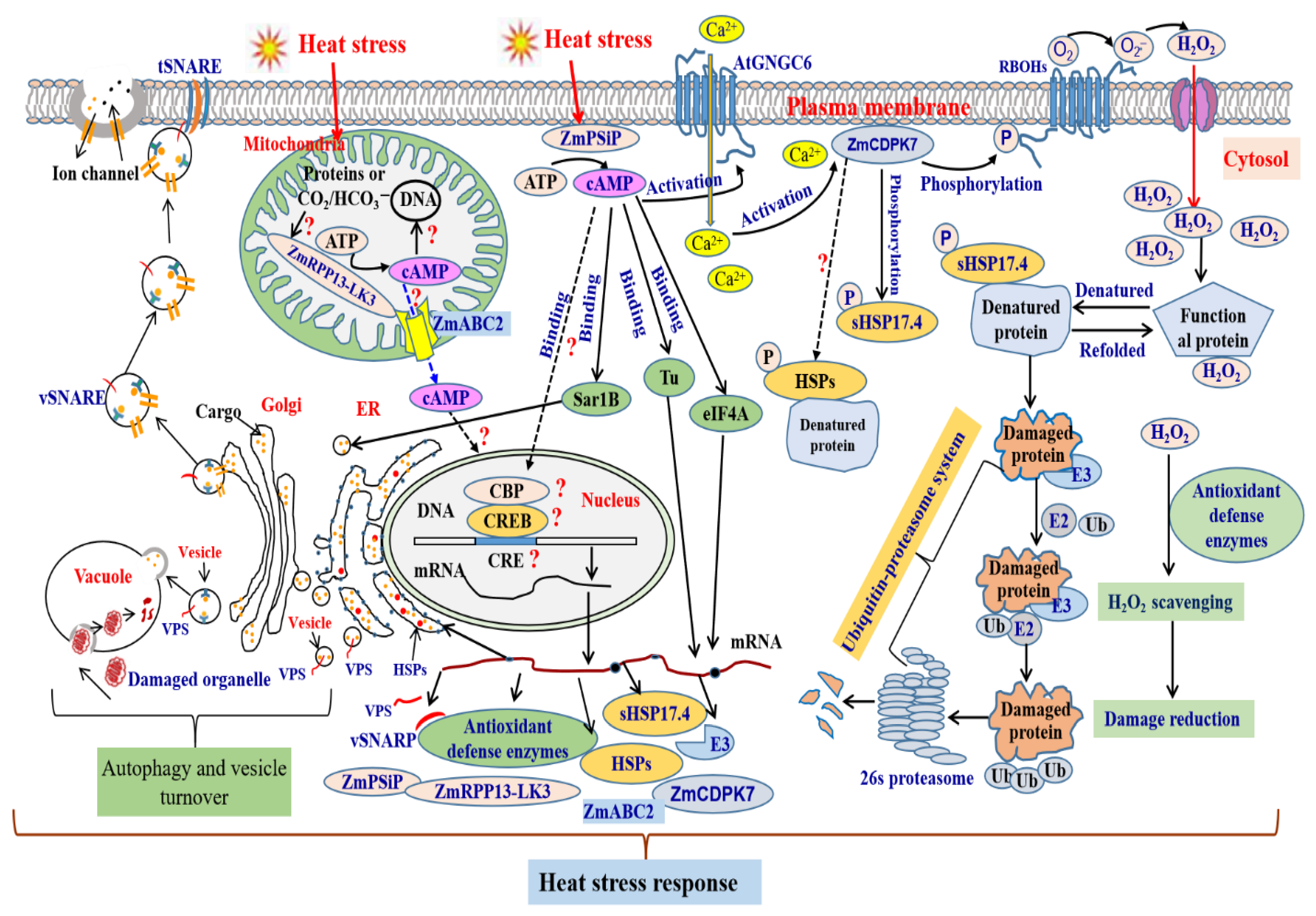
:1. Introduction
2. Diversity Features of the Catalytic Center Motifs in the Identified Plant ACs
3. cAMP Mediates Heat Stress Response in Plants
3.1. Crosstalk between cAMP and Ca2+ Signal under Heat Stress Conditions
3.2. cAMP Mediates H2O2 as Heat Stress Signals
3.3. Involvement of cAMP in ABA Signaling Pathway under Heat Stress
3.4. cAMP and HSP under Heat Stress
3.5. cAMP Mediates the Trafficking of Vesicle and Biomacromolecules under Heat Stress
3.6. cAMP Participating in Ubiquitin-Proteasome System (UPS) under Heat Stress
4. Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robichaux, W.G., 3rd; Cheng, X.D. Intracellular cAMP Sensor EPAC: Physiology, Pathophysiology, and Therapeutics Development. Physiol. Rev. 2018, 98, 919–1053. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Y.; Kong, Q.B.; Wang, J.; Jiang, Y.F.; Hua, H. Complex roles of cAMP-PKA-CREB signaling in cancer. Exp. Hematol. Oncol. 2020, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Lee, K. Epac: New emerging cAMP-binding protein. BMB Rep. 2021, 54, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Steegborn, C. Structure, mechanism, and regulation of soluble adenylyl cyclases—Similarities and differences to transmembrane adenylyl cyclases. BBA Mol. Basis Dis. 2014, 1842, 2535–2547. [Google Scholar] [CrossRef] [Green Version]
- Wiggins, S.V.; Steegborn, C.; Levin, L.R.; Buck, J. Pharmacological modulation of the CO2/HCO3−/pH-, calcium-, and ATP-sensing soluble adenylyl cyclase. Pharmacol. Ther. 2018, 190, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Blanco, E.; Fortunato, S.; Viggiano, L.; de Pinto, M.C. Cyclic AMP: A polyhedral signalling molecule in Plants. Int. J. Mol. Sci. 2020, 21, 4862. [Google Scholar] [CrossRef]
- Paradiso, A.; Domingo, G.; Blanco, E.; Buscaglia, A.; Fortunato, S.; Marsoni, M.; Scarcia, P.; Caretto, S.; Vannini, C.; de Pinto, M.C. Cyclic AMP mediates heat stress response by the control of redox homeostasis and ubiquitin-proteasome system. Plant Cell Environ. 2020, 43, 2727–2742. [Google Scholar] [CrossRef]
- Sabetta, W.; Vandelle, E.; Locato, V.; Costa, A.; Cimini, S.; Bittencourt Moura, A.; Luoni, L.; Graf, A.; Viggiano, L.; De Gara, L.; et al. Genetic buffering of cyclic AMP in Arabidopsis thaliana compromises the plant immune response triggered by an avirulent strain of Pseudomonas syringae pv. tomato. Plant J. 2019, 98, 590–606. [Google Scholar] [CrossRef]
- Yang, H.; Zhao, Y.; Chen, N.; Liu, Y.; Yang, S.; Du, H.; Wang, W.; Wu, J.; Tai, F.; Chen, F.; et al. A new adenylyl cyclase, putative disease resistance RPP13-like protein 3, participates in abscisic acid-mediated heat stress resistance in maize. J. Exp. Bot. 2021, 72, 283–301. [Google Scholar] [CrossRef]
- Zhao, Y.L.; Du, H.W.; Wang, Y.K.; Wang, H.L.; Yang, S.Y.; Li, C.H.; Chen, N.; Yang, H.; Zhang, Y.H.; Zhu, Y.L.; et al. The calcium-dependent protein kinase ZmCDPK7 functions in heat-stress tolerance in maize. J. Integr. Plant Biol. 2021, 63, 510–527. [Google Scholar] [CrossRef]
- Moutinho, A.; Hussey, P.J.; Trewavas, A.J.; Malhó, R. cAMP acts as a second messenger in pollen tube growth and reorientation. Proc. Natl. Acad. Sci. USA 2001, 98, 10481–10486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruzvidzo, O.; Dikobe, B.T.; Kawadza, D.T.; Mabadahanye, G.H.; Chatukuta, P.; Kwezi, L. Recombinant expression and functional testing of candidate adenylate cyclase domains. Methods Mol. Biol. 2013, 1016, 13–25. [Google Scholar] [PubMed]
- Bianche, C.; Wong, A.; Quaglia, M.; Alqurashi, M.; Gehring Chris Ntoukakis, V.; Pasqualini, S. An Arabidopsis thaliana leucine-rich repeat protein harbors an adenylyl cyclase catalytic center and affects responses to pathogens. J. Plant Physiol. 2019, 232, 12–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Younis, I.; Wong, A.; Lemtiri-Chlieh, F.; Schmöckel, S.M.; Tester, M.; Gehring, C.; Donaldson, L. The Arabidopsis thaliana K+-uptake permease 5 (AtKUP5) contains a functional cytosolic adenylate cyclase essential for K+ transport. Front. Plant Sci. 2018, 9, 1645. [Google Scholar] [CrossRef] [Green Version]
- Al-Younis, I.; Wong, A.; Gehring, C. The Arabidopsis thaliana K+-uptake permease 7 (AtKUP7) contains a functional cytosolic adenylate cyclase catalytic centre. FEBS Lett. 2015, 589, 3848–3852. [Google Scholar] [CrossRef] [Green Version]
- Chatukuta, P.; Dikobe, T.B.; Kawadza, D.T.; Sehlabane, K.S.; Takundwa, M.M.; Wong, A.; Gehring, C.; Ruzvidzo, O. An Arabidopsis clathrin assembly protein with a predicted role in plant defense can function as an adenylate cyclase. Biomolecules 2018, 8, 15. [Google Scholar] [CrossRef] [Green Version]
- Ito, M.; Takahashi, H.; Sawasaki, T.; Ohnishi, K.; Hikichi, Y.; Kiba, A. Novel type of adenylyl cyclase participates in tabtoxinine-β-lactam-induced cell death and occurrence of wildfire disease in Nicotiana benthamiana. Plant Signal. Behav. 2014, 9, e27420. [Google Scholar] [CrossRef] [Green Version]
- Al-Younis, I.; Moosa, B.; Kwiatkowski, M.; Jaworski, K.; Wong, A.; Gehring, C. Functional Crypto-Adenylate Cyclases Operate in Complex Plant Proteins. Front. Plant Sci. 2021, 12, 711749. [Google Scholar] [CrossRef]
- Swieżawska, B.; Jaworski, K.; Pawełek, A.; Grzegorzewska, W.; Szewczuk, P.; Szmidt-Jaworska, A. Molecular cloning and characterization of a novel adenylyl cyclase gene, HpAC1, involved in stress signaling in Hippeastrum × hybridum. Plant Physiol. Biochem. 2014, 80, 41–52. [Google Scholar] [CrossRef]
- Kasahara, M.; Suetsugu, N.; Urano, Y.; Yamamoto, C.; Ohmori, M.; Takada, Y.; Okuda, S.; Nishiyama, T.; Sakayama, H.; Kohchi, T.; et al. An adenylyl cyclase with a phosphodiesterase domain in basal plants with a motile sperm system. Sci. Rep. 2016, 6, 39232. [Google Scholar] [CrossRef] [Green Version]
- Wong, A.; Tian, X.C.; Gehring, C.; Marondedze, C. Discovery of novel functional centers with rationally designed amino acid motifs. Comput. Struct. Biotechnol. J. 2018, 16, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Ruzvidzo, O.; Gehring, C.; Wong, A. New Perspectives on Plant Adenylyl Cyclases. Front. Mol. Biosci. 2019, 6, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, S.; Schachtschabel, J.; Mishkind, M.; Munnik, T.; Arisz, S.A. Hot topic: Thermosensing in plants. Plant Cell Environ. 2021, 44, 2018–2033. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.L.; Liu, Y.P.; Sun, J.F.; Lv, S.S.; Luo, Y.M.; Yang, H.; Zhang, Y.P.; Zhao, X.; Hu, X.L. Physiological and proteomic analyses reveal cAMP-regulated key factors in maize root tolerance to heat stress. Food Energy Secur. 2021, 10, e309. [Google Scholar] [CrossRef]
- Liao, C.C.; Zheng, Y.; Guo, Y. MYB30 transcription factor regulates oxidative and heat stress responses through ANNEXIN-mediated cytosolic calcium signaling in Arabidopsis. New Phytol. 2017, 16, 163–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenzoni, G.; Knight, M.R. Increases in absolute temperature stimulate free calcium concentration elevations in the chloroplast. Plant Cell Physiol. 2019, 60, 538–548. [Google Scholar] [CrossRef]
- Pollastri, S.; Sukiran, N.A.; Jacobs, B.C.I.C.; Knight, M.R. Chloroplast calcium signalling regulates thermomemory. J. Plant Physiol. 2021, 264, 153470. [Google Scholar] [CrossRef]
- Niu, W.T.; Han, X.W.; Wei, S.S.; Shang, Z.L.; Wang, J.; Yang, D.W.; Fan, X.; Gao, F.; Zheng, S.Z.; Bai, J.T.; et al. Arabidopsis cyclic nucleotide-gated channel 6 is negatively modulated by multiple calmodulin isoforms during heat shock. J. Exp. Bot. 2020, 71, 90–104. [Google Scholar] [CrossRef]
- Thomas, L.; Marondedze, C.; Ederli, L.; Pasqualini, S.; Gehring, C. Proteomic signatures implicate cAMP in light and temperature responses in Arabidopsis thaliana. J. Proteom. 2013, 83, 47–59. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, L.P.; Qi, Y.; Xu, H. Mitochondrial cAMP signaling. Cell Mol. Life Sci. 2016, 73, 4577–4590. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Zaid, A.; Wani, S.H. Reactive Oxygen Species Generation, Scavenging and Signaling in Plant Defense Responses. In Bioactive Molecules in Plant Defense; Jogaiah, S., Abdelrahman, M., Eds.; Springer: Cham, Switzerland, 2019; pp. 111–132. [Google Scholar]
- Yao, Y.; He, R.J.; Xie, Q.L.; Zhao, X.H.; Deng, X.M.; He, J.B.; Song, L.; He, J.; Marchant, A.; Chen, X.Y.; et al. ETHYLENE RESPONSE FACTOR 74 (ERF74) plays an essential role in controlling a respiratory burst oxidase homolog D (RbohD) dependent mechanism in response to different stresses in Arabidopsis. New Phytol. 2017, 213, 1667–1681. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Miller, G.; Morales, J.; Shulaev, V.; Torres, M.A.; Mittler, R. Respiratory burst oxidases: The engines of ROS signaling. Curr. Opin. Plant Biol. 2011, 14, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Konigshofer, H.; Tromballa, H.W.; Loppert, H.G. Early events in signalling high-temperature stress in tobacco BY2 cells involve alterations in membrane fluidity and enhanced hydrogen peroxide production. Plant Cell Environ. 2008, 31, 1771–1780. [Google Scholar] [CrossRef]
- Sgobba, A.; Paradiso, A.; Dipierro, S.; De Gara, L.; de Pinto, M.C. Changes in antioxidants are critical in determining cell responses to short- and long-term heat stress. Physiol. Plant. 2015, 153, 68–78. [Google Scholar] [CrossRef]
- Gusan, S.; Anand-Srivastava, M.B. cAMP attenuates the enhanced expression of Gi proteins and hyperproliferation of vascular smooth muscle cells from SHR: Role of ROS and ROS-mediated signaling. Am. J. Physiol. 2013, 304, 1198–1209. [Google Scholar] [CrossRef] [Green Version]
- Palorini, R.; De Rasmo, D.; Gaviraghi, M.; Sala Danna, L.; Signorile, A.; Cirulli, C.; Chiaradonna, F.; Alberghina, L.; Papa, S. Oncogenic K-ras expression is associated with derangement of the cAMP/PKA pathway and forskolin-reversible alterations of mitochondrial dynamics and respiration. Oncogene 2013, 32, 352–362. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Li, Y.; Anand-Srivastava, M.B. Reduced levels of cyclic AMP contribute to the enhanced oxidative stress in vascular smooth muscle cells from spontaneously hypertensive rats. Can. J. Physiol. Pharm. 2008, 86, 190–198. [Google Scholar] [CrossRef]
- Peshavariya, H.M.; Liu, G.S.; Chang, C.W.; Jiang, F.; Chan, E.C.; Dusting, G.J. Prostacyclin signaling boosts NADPH oxidase 4 in the endothelium promoting cytoprotection and angiogenesis. Antioxid. Redox Signal. 2014, 20, 2710–2725. [Google Scholar] [CrossRef]
- Iqbal, N.; Umar, S.; Khan, N.A.; Corpas, F.J. Crosstalk between abscisic acid and nitric oxide under heat stress: Exploring new vantage points. Plant Cell Rep. 2021, 40, 1429–1450. [Google Scholar] [CrossRef]
- Huang, Y.C.; Niu, C.Y.; Yang, C.R.; Jinn, T.L. The Heat Stress Factor HSFA6b Connects ABA Signaling and ABA-Mediated Heat Responses. Plant Physiol. 2016, 172, 1182–1199. [Google Scholar] [CrossRef] [PubMed]
- Bellotti, M.; Salis, A.; Grozio, A.; Damonte, G.; Vigliarolo, T.; Galatini, A.; Zocchi, E.; Benatti, U.; Millo, E. Synthesis, structural characterization and effect on human granulocyte intracellular cAMP levels of abscisic acid analogs. Bioorg. Med. Chem. 2015, 23, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Sturla, L.; Fresia, C.; Guida, L.; Bruzzone, S.; Scarfì, S.; Usai, C.; Fruscione, F.; Magnone, M.; Millo, E.; Basile, G.; et al. LANCL2 is necessary for abscisic acid binding and signaling in human granulocytes and in rat insulinoma cells. J. Biol. Chem. 2009, 284, 28045–28057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigliarolo, T.; Guida, L.; Millo, E.; Fresia, C.; Turco, E.; De Flora, A.; Zocchi, E. Abscisic acid transport in human erythrocytes. J. Biol. Chem. 2015, 290, 13042–13052. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Wang, X.F.; Xu, M.; Gao, C.J.; Zeng, Y.L.; Cui, Y.; Shen, W.J.; Jiang, L.W. The roles of endomembrane trafficking in plant abiotic stress responses. J. Integr. Plant Biol. 2020, 62, 55–69. [Google Scholar] [CrossRef]
- Garcia, V.J.; Xu, S.L.; Ravikumar, R.; Wang, W.; Elliott, L.; Gonzalez, E.; Fesenko, M.; Altmann, M.; Brunschweiger, B.; Falter-Braun, P.; et al. TRIPP Is a Plant-Specific Component of the Arabidopsis TRAPPII Membrane Trafficking Complex with Important Roles in Plant Development. Plant Cell 2020, 32, 2424–2443. [Google Scholar] [CrossRef]
- Bian, C.; Ruan, Q.; Peng, Z.; Ji, H.; Jiang, L.; Li, J.; Yuan, L. Cloning, expression and characterization of the putative nuclear transport factor 2 (NTF2) gene from moss Conocephalum conicum (L.) Dum. Mol. Biol. Rep. 2011, 38, 2023–2032. [Google Scholar] [CrossRef]
- Xu, F.Q.; Xue, H.W. The ubiquitin-proteasome system in plant responses to environments. Plant Cell Environ. 2019, 42, 2931–2944. [Google Scholar] [CrossRef] [Green Version]
- Vierstra, R.D. The ubiquitin-26S proteasome system at the nexus of plant biology. Nat. Rev. Mol. Cell Biol. 2009, 10, 385–397. [Google Scholar] [CrossRef]
- McLoughlin, F.; Kim, M.; Marshall, R.S.; Vierstra, R.D.; Vierling, E. HSP101 interacts with the proteasome and promotes the clearance of ubiquitylated protein aggregates. Plant Physiol. 2019, 180, 1829–1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, J.; Yashiroda, H.; Maruya, M.; Yahara, I.; Tanaka, K. Proteasomes and molecular chaperones: Cellular machinery responsible for folding and destruction of unfolded proteins. Cell Cycle 2003, 2, 585–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stangl, K.; Günther, C.; Frank, T.; Lorenz, M.; Meiners, S.; Röpke, T.; Stelter, L.; Moobed, M.; Baumann, G.; Kloetzel, P.M.; et al. Inhibition of the ubiquitin-proteasome pathway induces differential heat-shock protein response in cardiomyocytes and renders early cardiac protection. Biochem. Biophys. Res. Commun. 2002, 291, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, L.; Meier, S.; Gehring, C. The arabidopsis cyclic nucleotide interactome. Cell Commun. Signal. 2016, 14, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shalaby, T.A.; Abd-Alkarim, E.; El-Aidy, F.; Hamed, E.S.; Sharaf-Eldin, M.; Taha, N.; El-Ramady, H.; Bayoumi, Y.; Dos Reis, A.R. Nano-selenium, silicon and H2O2 boost growth and productivity of cucumber under combined salinity and heat stress. Ecotoxicol. Environ. Saf. 2021, 212, 111962. [Google Scholar] [CrossRef] [PubMed]
- Seth, R.; Maritim, T.K.; Parmar, R.; Sharma, R.K. Underpinning the molecular programming attributing heat stress associated thermotolerance in tea (Camellia sinensis (L.) O. Kuntze). Hortic. Res. 2021, 8, 99. [Google Scholar] [CrossRef]
- Jin, X.C.; Wu, W.H. Involvement of cyclic AMP in ABA- and Ca2+-mediated signal transduction of stomatal regulation in Vicia faba. Plant Cell Physiol. 1999, 40, 1127–1133. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Han, X.; Wu, J.; Zheng, S.; Shang, Z.; Sun, D.; Zhou, R.; Li, B. A heat-activated calcium-permeable channel—Arabidopsis cyclic nucleotide-gated ion channel 6—Is involved in heat shock responses. Plant J. 2012, 70, 1056–1069. [Google Scholar] [CrossRef]


{kind=link}
| Organism | Protein Name | Accession Numbers | Catalytic Center Motif by [RKS]X[DE]X(9,11)[KR]X(1,3)[DE] | Extra Catalytic Center Motif by [RKS]X[DE]X(9,11)[KR]X(1,3) |
|---|---|---|---|---|
| Zea mays | Putative disease resistance RPP13-like protein 3 (ZmRPP13-LK3) | A0A1D6NWF3 | K174VDNARKMMTEKEEKKIWE192 | S23TEERMKKLFGDFEK37 |
| K185EEKKIWEDQKAKELEE201 | S539GEVSLPKDIHQMR552 | |||
| S498LERGVGTTTRKLRTLLD515 | S585IDHIPASLWRNR597 | |||
| S69GDQVGVIGFDEQIKQIE86 | S707NDHKILELGRIK719 | |||
| S820DDEKVFQHLPIWR833 | ||||
| Zea mays | Pollen-Signaling Protein (ZmPSiP) | AJ307886 | S146DDAKRDWFSRDVCKNCSD164 | R43IDAIISHEERRR55 |
| S346EELATLKDVGIKIAE361 | S155RDVCKNCSDAMK167 | |||
| S698LDRATSGASALANKPFLE716 | K193VDVFAIVGAVGIGK207 | |||
| R115HEIGFTIRDIDLRLRE131 | K259EELLILLASALSKR273 | |||
| K741EEKEGQERSNGQCRGDE758 | S482VERCWITHHLLR494 | |||
| S1049CDGKRYFRYNKSRRIYE1066 | S528MENSLDGPISLK540 | |||
| S532LDGPISLKQQMGLR546 | ||||
| K736DETEKEEKEGQER749 | ||||
| S933DELHLKDNKVLQR946 | ||||
| Arabidopsis thaliana | LRR and NB-ARC domains-containing disease resistance protein (AtLRRAC1) | At3g14460 | K121MEKVVRLLEHHVKHIE137 | R310SEIVSTVAKAEK322 |
| R1302I EWGLRDLENLRNLE1317 | R505LEDDNIPEIPSTTR519 | |||
| R159PDDLPQGRLVGRVED174 | S1128LESFPGSHPPTTLK1142 | |||
| S1377IDEDLPPLSCLR1389 | ||||
| Arabidopsis thaliana | Pentatricopeptide (PPR) repeat-containing protein (AtPPR-AC1) | AT1G62590- | K100FDVVISLGEKMQRLE115 | K65LDDAIGLFGGMVK78 |
| R238GDTDLALNLLNKME252 | R380LDKAKQMFEFMVSK394 | |||
| K485LEKALEVFDYMQKSE500 | R415VEDGTELFREMSHR429 | |||
| K520VDDGWDLFCSLSLK534 | ||||
| Arabidopsis thaliana | K+ uptake permease 7 (AtKUP7) | AT5G09400 | S80FDVEALEVPGAPRNDYE97 | S61DEDEIPEHRLIR73 |
| S697LEKFIRREAQERSLE712 | S774SDSSVSEAEQSLER788 | |||
| S785LERELSFIHKAK797 | ||||
| Arabidopsis thaliana | K+ uptake permease 5 (AtKUP5) | AT4G33530 | S81FDVDALEIPGTQKNEIE98 | S61DEEDDNVEQRLIR74 |
| S212PELERSLIIKERLE226 | S782LEKELSFIHKAK794 | |||
| S698LEKFIRKEAQERALE713 | ||||
| Arabidopsis thaliana | Clathrin assembly protein (AtClAP) | AT1G68110 | K329WEIFEDDYRCFDRKDKWE347 | S43HDDSSVDYSNAHR56 |
| S48VDYSNAHRVYKWIR62 | ||||
| S165LEKTSDSVIQELER179 | ||||
| K236SEAATVLKIVNK248 | ||||
| K307EDEKAMVVLEQPKK321 | ||||
| Hippeastrum hybrid cultivar | Adenylate cyclase (HpAC1) | ADM83595 | S160YEIECETTEPERVKGLLE178 | |
| Marchantia polymorpha | Hypothetical protein MARPO_0068s0004 (MpCAPE) | PTQ35772 | K486GEAVQEGYQQDHSKLE502 | K7DDKGKDQEENDEAK21 |
| K788AEPETEDEYNQRYE802 | S326SDAPPSPKRLPDK339 | |||
| R800YEAPLSLLFGAPTREE816 | S597QEQRGSPSPGVQYR611 | |||
| R814EELMLPAKIGKDK827 | ||||
| S1053PDYEEEEKFLSK1065 | ||||
| S1242RDSNRLLIQPIER1255 | ||||
| S1419IETSQSLAEFAK1431 | ||||
| Nicotiana benthamiana | Adenylyl cyclase (NbAC) | ACR77530 | R354LEVIKRQKDEKRKE368 | R135EDVNFLHDILQRLR149 |
| Arabidopsis thaliana | Nine-cis-epoxycaroteoid dioygenase 3 (crypto-AC) | S311YDVVSKPYLKYFRFSPD328 | S29SDLSYCSSLPMASR43 | |
| S304GELFALSYDVVSK317 | ||||
| S441DENLKSVLSEIR453 | ||||
| S567LEVEATVKLPSR579 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, S.; Sun, J.; Luo, Y.; Lv, S.; Chen, J.; Liu, Y.; Hu, X. cAMP Is a Promising Regulatory Molecule for Plant Adaptation to Heat Stress. Life 2022, 12, 885. https://doi.org/10.3390/life12060885
Liang S, Sun J, Luo Y, Lv S, Chen J, Liu Y, Hu X. cAMP Is a Promising Regulatory Molecule for Plant Adaptation to Heat Stress. Life. 2022; 12(6):885. https://doi.org/10.3390/life12060885
Chicago/Turabian StyleLiang, Shuang, Jinfeng Sun, Yanmin Luo, Shanshan Lv, Jiajia Chen, Yanpei Liu, and Xiuli Hu. 2022. "cAMP Is a Promising Regulatory Molecule for Plant Adaptation to Heat Stress" Life 12, no. 6: 885. https://doi.org/10.3390/life12060885