
Daeshiho-tang Attenuates Atherosclerosis by Regulating Cholesterol Metabolism and Inducing M2 Macrophage Polarization


Abstract
:1. Introduction
2. Materials and Methods
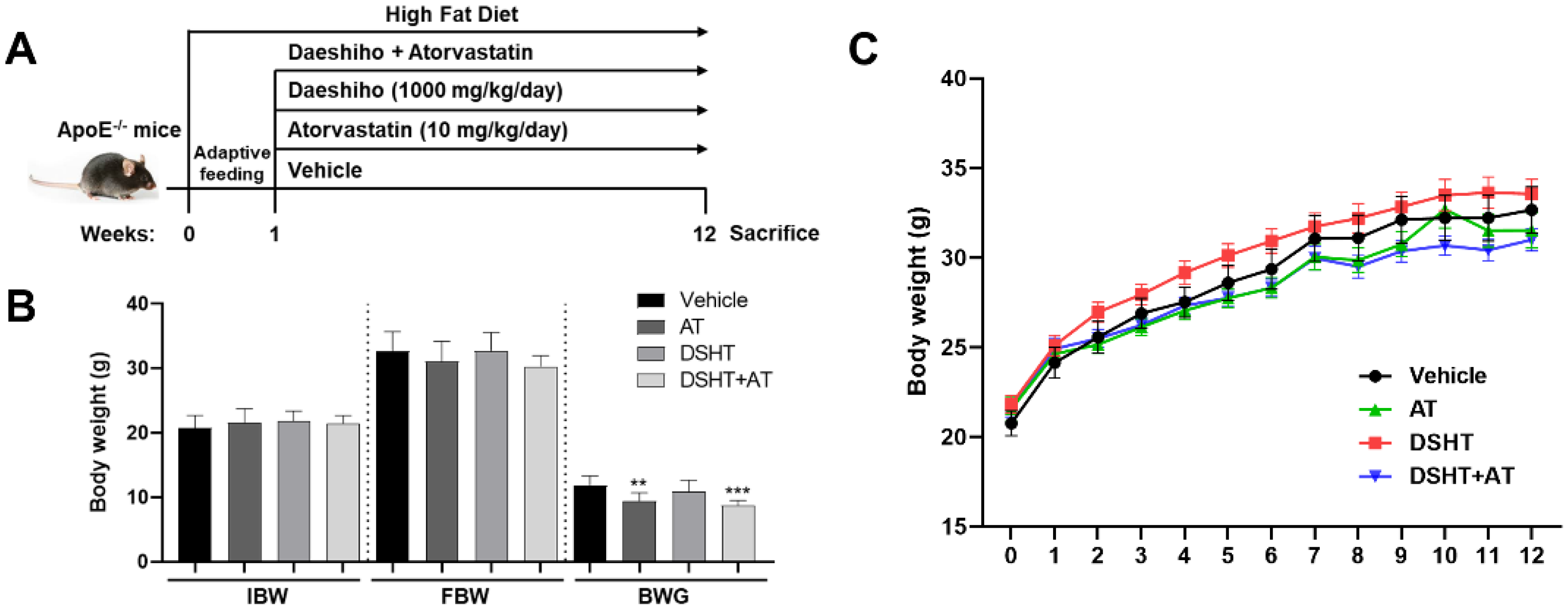
2.1. Animals
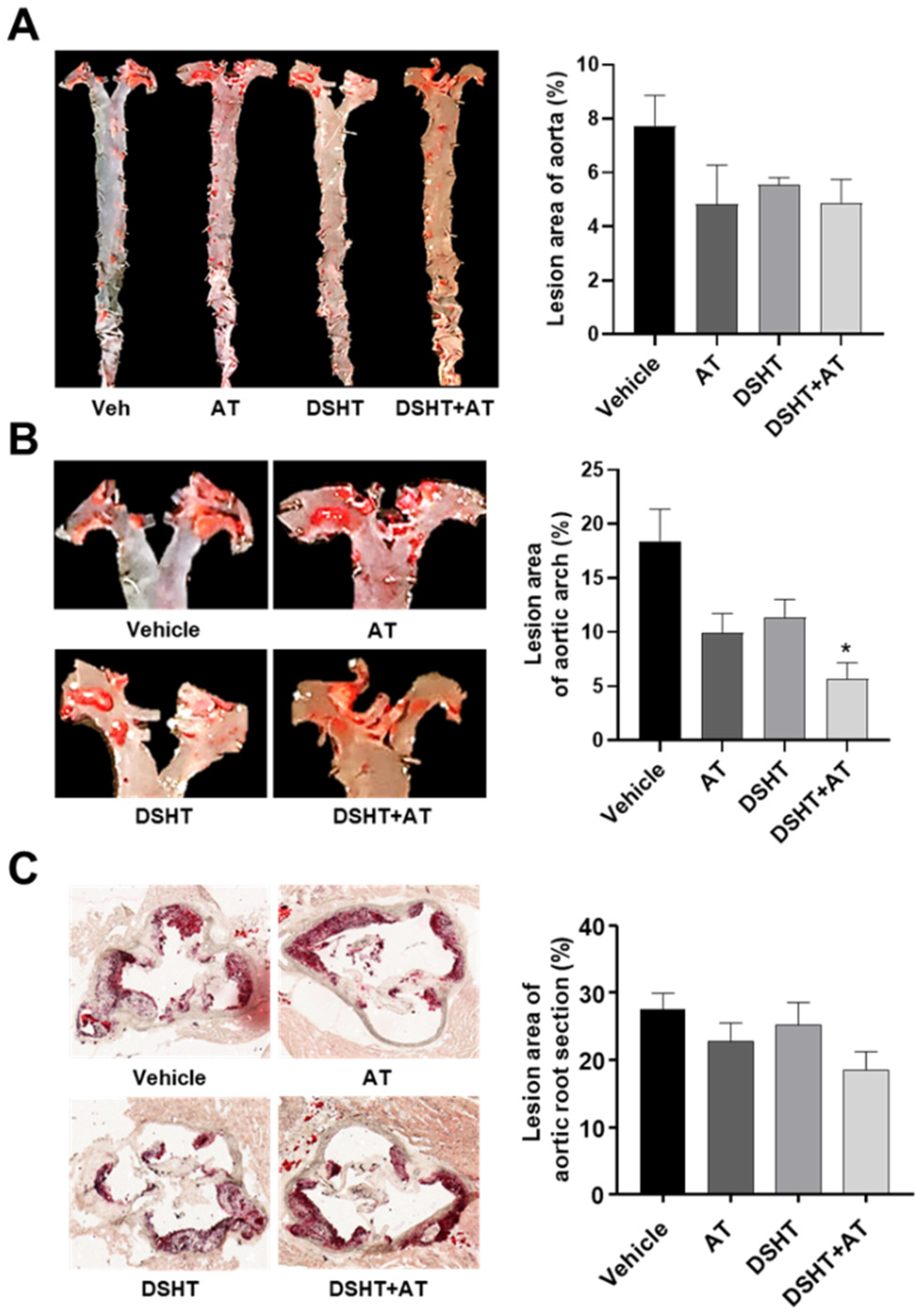
2.2. Quantification of Atherosclerotic Lesions
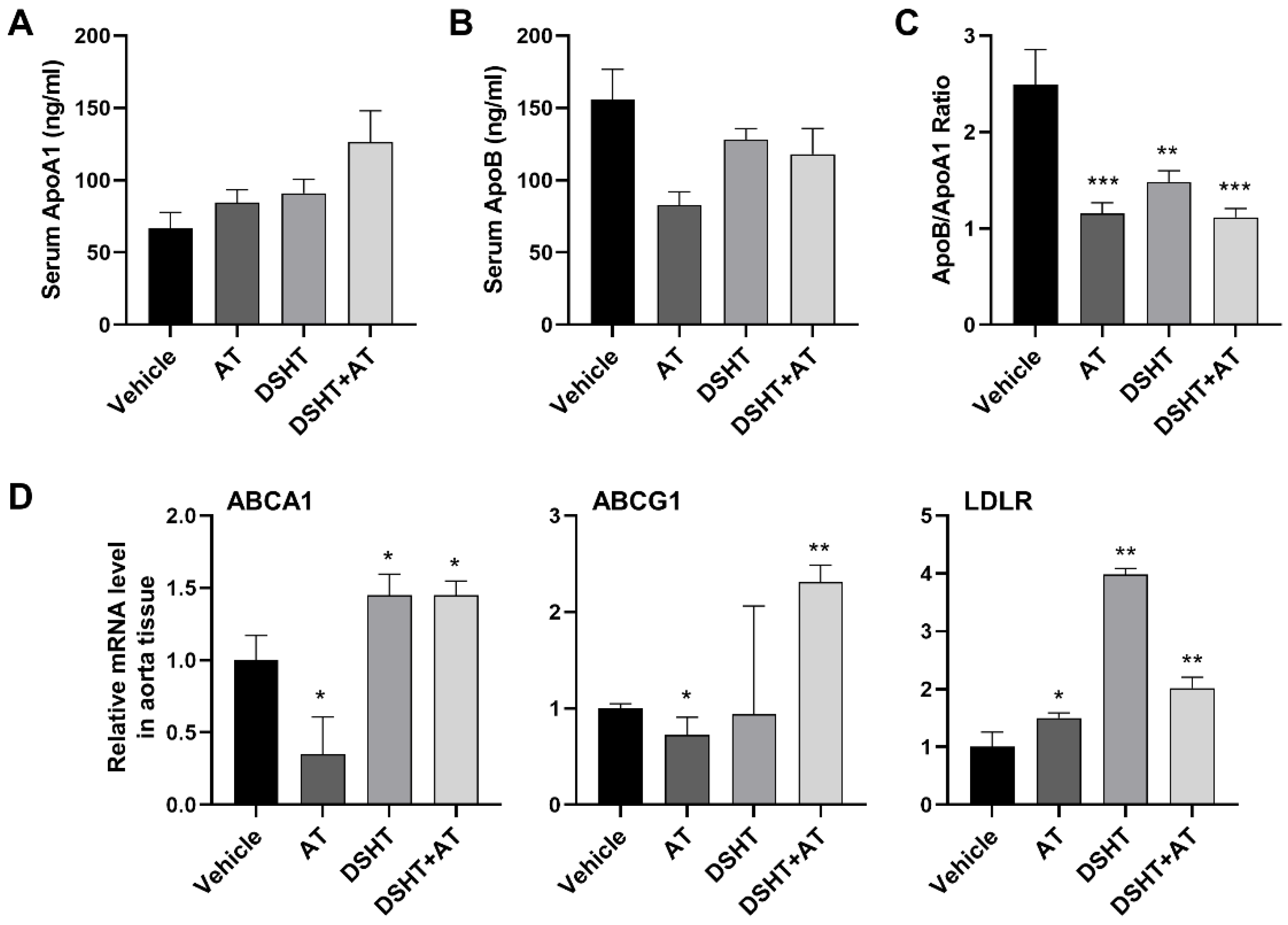
2.3. Measurement of Serum Cholesterol Parameters
2.4. RNA Isolation and Quantitative qPCR Analysis
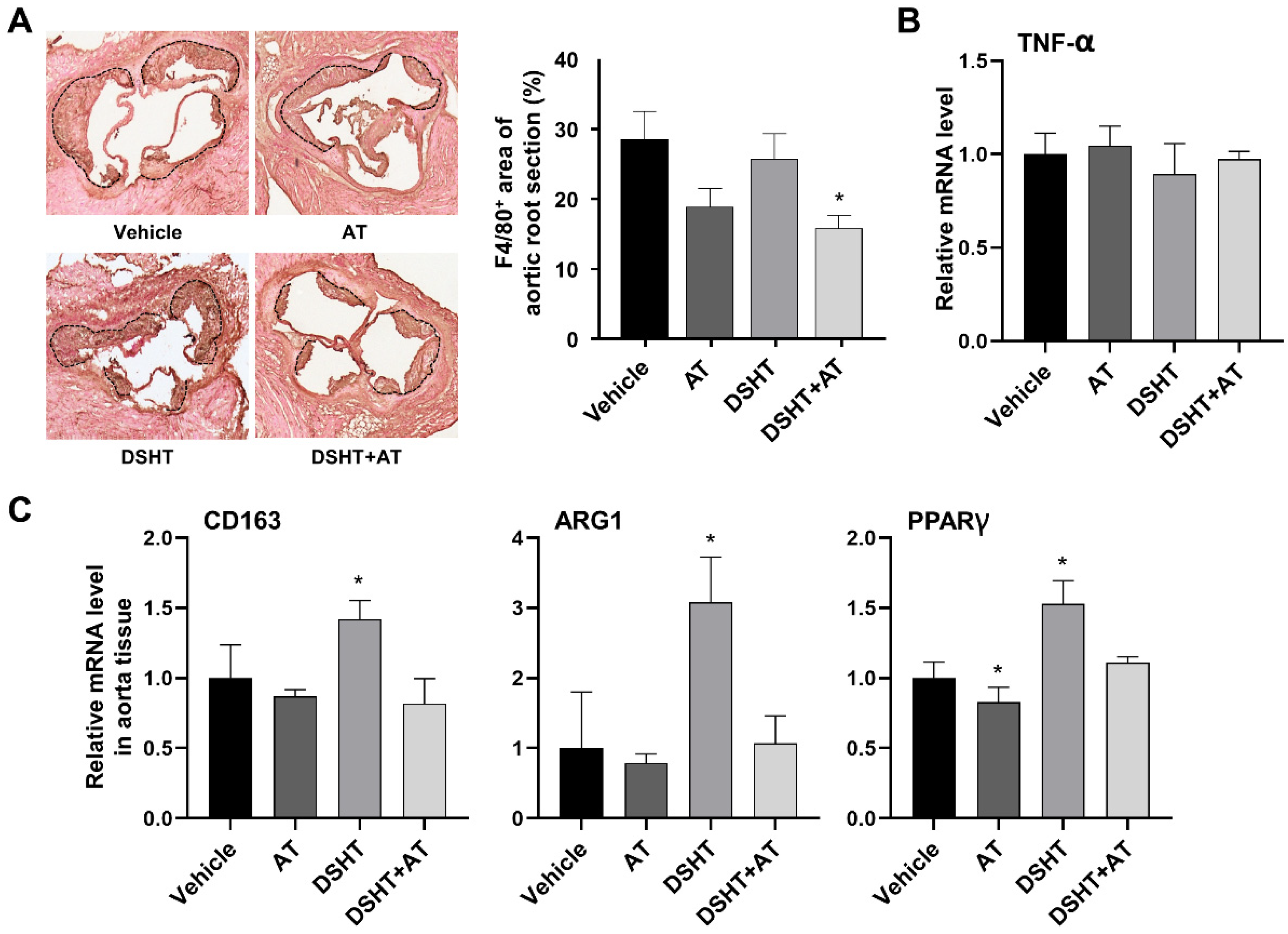
2.5. Immunohistochemistry for Macrophages
2.6. Statistical Analysis
3. Results
3.1. General Observations
3.2. DSHT Reduces Atherosclerotic Plaques in the Aortic Arch
3.3. DSHT Decreases Blood Lipid Level
3.4. DSHT May Influence M2 Macrophage Polarization in Atherosclerosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| LDL-C | Low-density lipoprotein cholesterol |
| HDL-C | High-density lipoprotein cholesterol |
| DSHT | Daeshiho-tang |
| AT | Atorvastatin |
| Apo | Apolipoprotein |
| HFD | High-fat diet |
References
- Bäck, M.; Yurdagul, A.; Tabas, I.; Öörni, K.; Kovanen, P.T. Inflammation and its resolution in atherosclerosis: Mediators and therapeutic opportunities. Nat. Rev. Cardiol. 2019, 16, 389–406. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.J.; Sheedy, F.J.; Fisher, E.A. Macrophages in atherosclerosis: A dynamic balance. Nat. Rev. Immunol. 2013, 13, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.J.; Tabas, I. Macrophages in the pathogenesis of atherosclerosis. Cell 2011, 145, 341–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, N.J.; Robinson, J.G.; Lichtenstein, A.H.; Merz, C.N.B.; Blum, C.B.; Eckel, R.H.; Goldberg, A.C.; Gordon, D.; Levy, D.; Lloyd-Jones, D.M.; et al. 2013 ACC/AHA guideline on the treatment of blood cholesterol to reduce atherosclerotic cardiovascular risk in adults: A report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines. J. Am. Coll. Cardiol. 2014, 63, 2889–2934. [Google Scholar] [CrossRef] [Green Version]
- Cholesterol Treatment Trialists’ (CTT) Collaboration; Baigent, C.; Blackwell, L.; Emberson, J.; Holland, L.E.; Reith, C.; Bhala, N.; Peto, R.; Barnes, E.H.; Keech, A.; et al. Efficacy and safety of more intensive lowering of LDL cholesterol: A meta-analysis of data from 170,000 participants in 26 randomised trials. Lancet 2010, 376, 1670–1681. [Google Scholar] [CrossRef] [Green Version]
- Cannon, C.P.; Blazing, M.A.; Giugliano, R.P.; McCagg, A.; White, J.A.; Theroux, P.; Darius, H.; Lewis, B.S.; Ophuis, T.O.; Jukema, J.W.; et al. Ezetimibe Added to Statin Therapy after Acute Coronary Syndromes. N. Engl. J. Med. 2015, 372, 2387–2397. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Shim, S.H. Medicinal Herbs Effective Against Atherosclerosis: Classification According to Mechanism of Action. Biomol. Ther. 2019, 27, 254–264. [Google Scholar] [CrossRef]
- Torres, N.; Guevara-Cruz, M.; Velazquez-Villegas, L.A.; Tovar, A.R. Nutrition and Atherosclerosis. Arch. Med. Res. 2015, 46, 408–426. [Google Scholar] [CrossRef]
- Jeong, S.-M.; Noh, J.-W.; Lee, M.-S.; Yang, H.-G.; Ahn, Y.-M.; Ahn, S.-Y.; Lee, B.-C. A Retrospective Study on the Effect of Herbal Extracts Combined with Conventional Therapy on Blood Glucose in Type 2 Diabetes Mellitus. J. Intern. Korean Med. 2020, 41, 1231–1244. [Google Scholar] [CrossRef]
- Kim, C.-H.; Kwon, Y.-M.; Lee, Y.-T.; Park, S.-D. The preventive effect of Daeshiho-tang on liver damage induced by acetaminophen in the rats. Herb. Formula Sci. 2004, 12, 139–154. [Google Scholar]
- Yeo, U.-H.; Jo, H.-J.; Kim, H.-H. Effects of Daesiho-tang extract on Hypertension and Arterial Contraction. J. Physiol. Pathol. Korean Med. 2005, 19, 1573–1579. [Google Scholar]
- Lee, J.-C.; Lee, E. Effects of Siho (Bupleuri Radix) extracts on serum lipid composition and antioxidant system in rat fed high fat diet. Herb. Formula Sci. 2003, 11, 151–159. [Google Scholar]
- Lee, H.; Kang, R.; Hahn, Y.; Yang, Y.; Kim, S.S.; Cho, S.H.; Chung, S.I.; Yoon, Y. Antiobesity effect of baicalin involves the modulations of proadipogenic and antiadipogenic regulators of the adipogenesis pathway. Phytother. Res. 2009, 23, 1615–1623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fan, S.; Hu, N.; Gu, M.; Chu, C.; Li, Y.; Lu, X.; Huang, C. Rhein Reduces Fat Weight in db/db Mouse and Prevents Diet-Induced Obesity in C57Bl/6 Mouse through the Inhibition of PPARgamma Signaling. PPAR Res. 2012, 2012, 374936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.L.; Wang, Y.J.; Yang, Y.; Yang, R.C.; Zhu, B.; Zhang, Y.; Lin, Y.; Lu, Y.; Li, X.F.; O’Byrne, K.T. Suppression of lipopolysaccharide-induced upregulation of toll-like receptor 4 by emodin in mouse proximal tubular epithelial cells. Mol. Med. Rep. 2012, 6, 493–500. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.M.; Choi, S.W.; Cho, S.H.; Lee, S. Effect of Seeds Extract of Paeonia Lactiflora on Antioxidative System and Lipid Peroxidation of Liver in Rats Fed High-Cholesterol Dietentration Using a FIA Biosensor. Korean J. Nutr. 2003, 36, 793–800. [Google Scholar]
- Liu, J.C.; Chan, P.; Hsu, F.L.; Chen, Y.J.; Hsieh, M.H.; Lo, M.Y.; Lin, J.Y. The in vitro inhibitory effects of crude extracts of traditional Chinese herbs on 3-hydroxy-3-methylglutaryl-coenzyme A reductase on Vero cells. Am. J. Chin. Med. 2002, 30, 629–636. [Google Scholar] [CrossRef]
- Noh, J.-W.; Jeong, S.-M.; Kim, D.-H.; Yoo, J.-H.; Ahn, Y.-M.; Ahn, S.-Y.; Lee, B.-C. A Retrospective Study on the Effect of Daeshiho-tang on the Lipid Profile in Patients with Uncontrolled Dyslipidemia by Statins. J. Intern. Korean Med. 2019, 40, 1026–1034. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Lin, S.; Vanhoutte, P.M.; Woo, C.W.; Xu, A. Akkermansia muciniphila protects against atherosclerosis by preventing metabolic endotoxemia-induced inflammation in Apoe−/− mice. Circulation 2016, 133, 2434–2446. [Google Scholar] [CrossRef] [Green Version]
- Song, M.-Y.; Kim, E.-K.; Kong, J.-C.; Lee, J.-H.; Shin, B.-C.; Ryu, D.-G.; Kwon, K.-B. Inhibitory effect of Daesiho-tang (Dachaihu-tang) extracts on high-fat diet-induced obesity. J. Korean Med. Rehabil. 2008, 18, 41–49. [Google Scholar]
- Roth, L.; Rombouts, M.; Schrijvers, D.M.; Martinet, W.; De Meyer, G.R. Cholesterol-independent effects of atorvastatin prevent cardiovascular morbidity and mortality in a mouse model of atherosclerotic plaque rupture. Vasc. Pharmacol. 2016, 80, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Peled, M.; Fisher, E.A. Dynamic Aspects of Macrophage Polarization during Atherosclerosis Progression and Regression. Front. Immunol. 2014, 5, 579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randolph, G.J. Mechanisms that regulate macrophage burden in atherosclerosis. Circ. Res. 2014, 114, 1757–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawashima, T.; Ogata, M.; Fujita, N.; Takahashi, R. Daisaikoto Prevents Post-dieting Weight Regain by Reversing Dysbiosis and Reducing Serum Corticosterone in Mice. Front. Physiol. 2019, 10, 1483. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Yuan, H.Q.; Hao, Y.M.; Ren, Z.; Qu, S.L.; Liu, L.S.; Wei, D.H.; Tang, Z.H.; Zhang, J.F.; Jiang, Z.S. Macrophage polarization in atherosclerosis. Clin. Chim. Acta 2020, 501, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Tomita, H.; Zhilicheva, S.; Kim, S.; Maeda, N. Aortic arch curvature and atherosclerosis have overlapping quantitative trait loci in a cross between 129S6/SvEvTac and C57BL/6J apolipoprotein E-null mice. Circ. Res. 2010, 106, 1052–1060. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Xu, P.; Xie, X.; Li, J.; Zhang, J.; Wang, J.; Hong, F.; Li, J.; Zhang, Y.; Song, Y. DBZ (Danshensu Bingpian Zhi), a Novel Natural Compound Derivative, Attenuates Atherosclerosis in Apolipoprotein E–Deficient Mice. J. Am. Heart Assoc. 2017, 6, e006297. [Google Scholar] [CrossRef] [Green Version]
- He, X.W.; Yu, D.; Li, W.L.; Zheng, Z.; Lv, C.L.; Li, C.; Liu, P.; Xu, C.Q.; Hu, X.F.; Jin, X.P. Anti-atherosclerotic potential of baicalin mediated by promoting cholesterol efflux from macrophages via the PPARgamma-LXRalpha-ABCA1/ABCG1 pathway. Biomed. Pharmacother. 2016, 83, 257–264. [Google Scholar] [CrossRef]
- He, D.; Wang, H.; Xu, L.; Wang, X.; Peng, K.; Wang, L.; Liu, P.; Qu, P. Saikosaponin-a Attenuates Oxidized LDL Uptake and Prompts Cholesterol Efflux in THP-1 Cells. J. Cardiovasc. Pharmacol. 2016, 67, 510–518. [Google Scholar] [CrossRef]
- Sukhorukov, V.N.; Khotina, V.A.; Chegodaev, Y.S.; Ivanova, E.; Sobenin, I.A.; Orekhov, A.N. Lipid metabolism in macrophages: Focus on atherosclerosis. Biomedicines 2020, 8, 262. [Google Scholar] [CrossRef]
- Fotakis, P.; Kothari, V.; Thomas, D.G.; Westerterp, M.; Molusky, M.M.; Altin, E.; Abramowicz, S.; Wang, N.; He, Y.; Heinecke, J.W. Anti-inflammatory effects of HDL (high-density lipoprotein) in macrophages predominate over proinflammatory effects in atherosclerotic plaques. Arterioscler. Thromb. Vasc. Biol. 2019, 39, e253–e272. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.J.; Distel, E.; Murphy, A.J.; Hu, J.; Garshick, M.S.; Ogando, Y.; Liu, J.; Vaisar, T.; Heinecke, J.W.; Berger, J.S. Apolipoprotein AI) promotes atherosclerosis regression in diabetic mice by suppressing myelopoiesis and plaque inflammation. Circulation 2019, 140, 1170–1184. [Google Scholar] [CrossRef] [PubMed]
- Yvan-Charvet, L.; Wang, N.; Tall, A.R. Role of HDL, ABCA1, and ABCG1 transporters in cholesterol efflux and immune responses. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 139–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.M.; Moon, J.; Cho, Y.; Chung, J.H.; Shin, M.J. Quercetin up-regulates expressions of peroxisome proliferator-activated receptor gamma, liver X receptor alpha, and ATP binding cassette transporter A1 genes and increases cholesterol efflux in human macrophage cell line. Nutr. Res. 2013, 33, 136–143. [Google Scholar] [CrossRef]
- Larrede, S.; Quinn, C.M.; Jessup, W.; Frisdal, E.; Olivier, M.; Hsieh, V.; Kim, M.J.; Van Eck, M.; Couvert, P.; Carrie, A.; et al. Stimulation of cholesterol efflux by LXR agonists in cholesterol-loaded human macrophages is ABCA1-dependent but ABCG1-independent. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1930–1936. [Google Scholar] [CrossRef]
- Ogata, M.; Tsujita, M.; Hossain, M.A.; Akita, N.; Gonzalez, F.J.; Staels, B.; Suzuki, S.; Fukutomi, T.; Kimura, G.; Yokoyama, S. On the mechanism for PPAR agonists to enhance ABCA1 gene expression. Atherosclerosis 2009, 205, 413–419. [Google Scholar] [CrossRef] [Green Version]
- Chawla, A.; Boisvert, W.A.; Lee, C.H.; Laffitte, B.A.; Barak, Y.; Joseph, S.B.; Liao, D.; Nagy, L.; Edwards, P.A.; Curtiss, L.K.; et al. A PPAR gamma-LXR-ABCA1 pathway in macrophages is involved in cholesterol efflux and atherogenesis. Mol. Cell 2001, 7, 161–171. [Google Scholar] [CrossRef]
- Pirahanchi, Y.; Huecker, M.R. Biochemistry, Ldl Cholesterol; StatPearls Publishing: Treasure Island, FL, USA, 2018. [Google Scholar]
- Feingold, K.R.; Grunfeld, C. Introduction to Lipids and Lipoproteins; MDText.com, Inc.: South Dartmouth, MA, USA, 2015. [Google Scholar]
- Pennings, M.; Meurs, I.; Ye, D.; Out, R.; Hoekstra, M.; Van Berkel, T.J.; Van Eck, M. Regulation of cholesterol homeostasis in macrophages and consequences for atherosclerotic lesion development. FEBS Lett. 2006, 580, 5588–5596. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.; Chen, Y.; Hu, W.; Li, X.; Yang, X.; Zhou, X.; Yin, Z.; Kong, D.; Yao, Z.; Hajjar, D.P.; et al. Peroxisome Proliferator-activated receptor gamma activation by ligands and dephosphorylation induces proprotein convertase subtilisin kexin type 9 and low density lipoprotein receptor expression. J. Biol. Chem. 2012, 287, 23667–23677. [Google Scholar] [CrossRef] [Green Version]
- Duffield, J.S. The inflammatory macrophage: A story of Jekyll and Hyde. Clin. Sci. 2003, 104, 27–38. [Google Scholar] [CrossRef]
- Cardilo-Reis, L.; Gruber, S.; Schreier, S.M.; Drechsler, M.; Papac-Milicevic, N.; Weber, C.; Wagner, O.; Stangl, H.; Soehnlein, O.; Binder, C.J. Interleukin-13 protects from atherosclerosis and modulates plaque composition by skewing the macrophage phenotype. EMBO Mol. Med. 2012, 4, 1072–1086. [Google Scholar] [CrossRef] [PubMed]
- Khallou-Laschet, J.; Varthaman, A.; Fornasa, G.; Compain, C.; Gaston, A.T.; Clement, M.; Dussiot, M.; Levillain, O.; Graff-Dubois, S.; Nicoletti, A.; et al. Macrophage plasticity in experimental atherosclerosis. PLoS ONE 2010, 5, e8852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Q.; Liu, J.; Zhang, Z.; Li, F.; Zhang, C.; Lai, B.; Xiao, L.; Wang, N. Peroxisome proliferator–Activated receptor γ (PPARγ) induces the gene expression of integrin αVβ5 to promote macrophage M2 polarization. J. Biol. Chem. 2018, 293, 16572–16582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.M.; Ni, X.X.; Xu, Q.Y.; Wang, Q.; Li, X.Y.; Hua, J. Regulation of lipid-induced macrophage polarization through modulating peroxisome proliferator-activated receptor-gamma activity affects hepatic lipid metabolism via a Toll-like receptor 4/NF-κB signaling pathway. J. Gastroenterol. Hepatol. 2020, 35, 1998–2008. [Google Scholar] [CrossRef] [PubMed]
- Sanson, M.; Distel, E.; Fisher, E.A. HDL induces the expression of the M2 macrophage markers arginase 1 and Fizz-1 in a STAT6-dependent process. PLoS ONE 2013, 8, e74676. [Google Scholar] [CrossRef] [PubMed]
- Huo, X.; Lu, F.; Qiao, L.; Li, G.; Zhang, Y. A component formula of Chinese medicine for hypercholesterolemia based on virtual screening and biology network. Evid.-Based Complement. Altern. Med. 2018, 2018, 1854972. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Botanic Name | Full Name | Relative Amount (g) | % of Total Weight |
|---|---|---|---|
| Bupleuri Radix | Bupleurum falcatum Linné | 2.00 | 30 |
| Pinelliae Tuber | Pinellia ternata Breitenbach | 1.33 | 20 |
| Scutellariae Radix | Scutellaria baicalensis Georgi | 1.00 | 15 |
| Paeoniae Radix | Paeonia lactiflora Pallas | 1.00 | 15 |
| Rhei Rhizoma | Rheum palmatum Linne | 0.67 | 10 |
| Ponciri Fructus | Poncirus trifoliata Rafinesque | 0.67 | 10 |
| Gene | Forward Primers | Reverse Primers |
|---|---|---|
| ABCA1 | AAAACCGCAGACATCCTTCAG | CATACCGAAACTCGTTCACCC |
| ABCG1 | GACACCGATGTGAACCCGTTTC | GCATGATGCTGAGGAAGGTCCT |
| LDLR | TGACTCAGACGAACAAGGCTG | ATCTAGGCAATCTCGGTCTCC |
| TNF-α | GCCTCTTCTCATTCCTGCTTG | CTGATGAGAGGGAGGCCATT |
| CD163 | TCCACACGTCCAGAACAGTC | CCTTGGAAACAGAGACAGGC |
| ARG1 | CTCCAAGCCAAAGTCCTTAGAG | AGGAGCTGTCATTAGGGACATC |
| PPARγ | TGGAATTAGATGACAGCGACTTGG | CTGGAGCAGCTTGGCAAACA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, M.-Y.; Cho, H.; Lee, S.; Lee, K.H.; Kim, W. Daeshiho-tang Attenuates Atherosclerosis by Regulating Cholesterol Metabolism and Inducing M2 Macrophage Polarization. Life 2022, 12, 197. https://doi.org/10.3390/life12020197
Song M-Y, Cho H, Lee S, Lee KH, Kim W. Daeshiho-tang Attenuates Atherosclerosis by Regulating Cholesterol Metabolism and Inducing M2 Macrophage Polarization. Life. 2022; 12(2):197. https://doi.org/10.3390/life12020197
Chicago/Turabian StyleSong, Min-Young, Haneul Cho, Sora Lee, Kyung Hye Lee, and Weon Kim. 2022. "Daeshiho-tang Attenuates Atherosclerosis by Regulating Cholesterol Metabolism and Inducing M2 Macrophage Polarization" Life 12, no. 2: 197. https://doi.org/10.3390/life12020197