
Bim Expression Promotes the Clearance of Mononuclear Phagocytes during Choroidal Neovascularization, Mitigating Scar Formation in Mice

 and
and 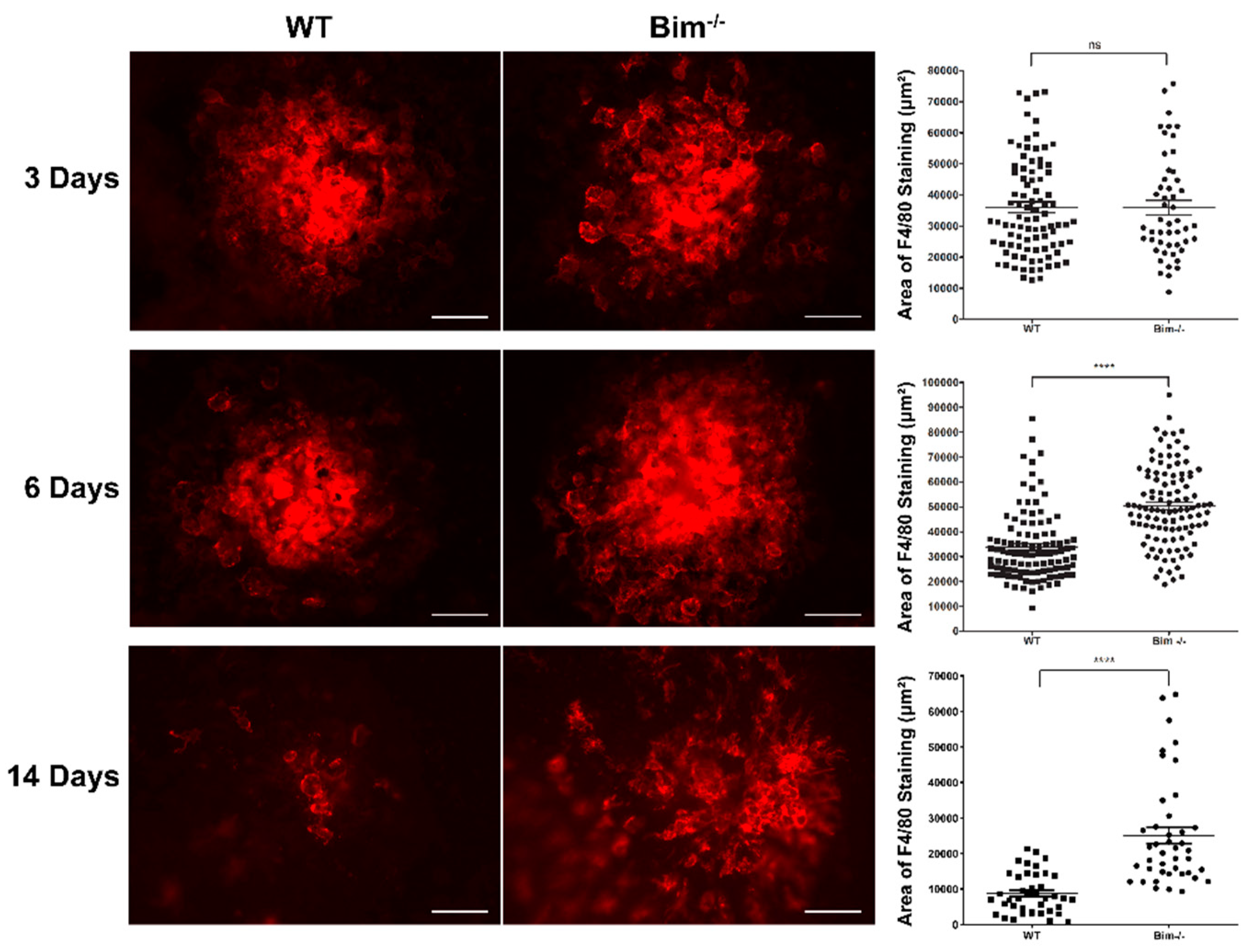{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
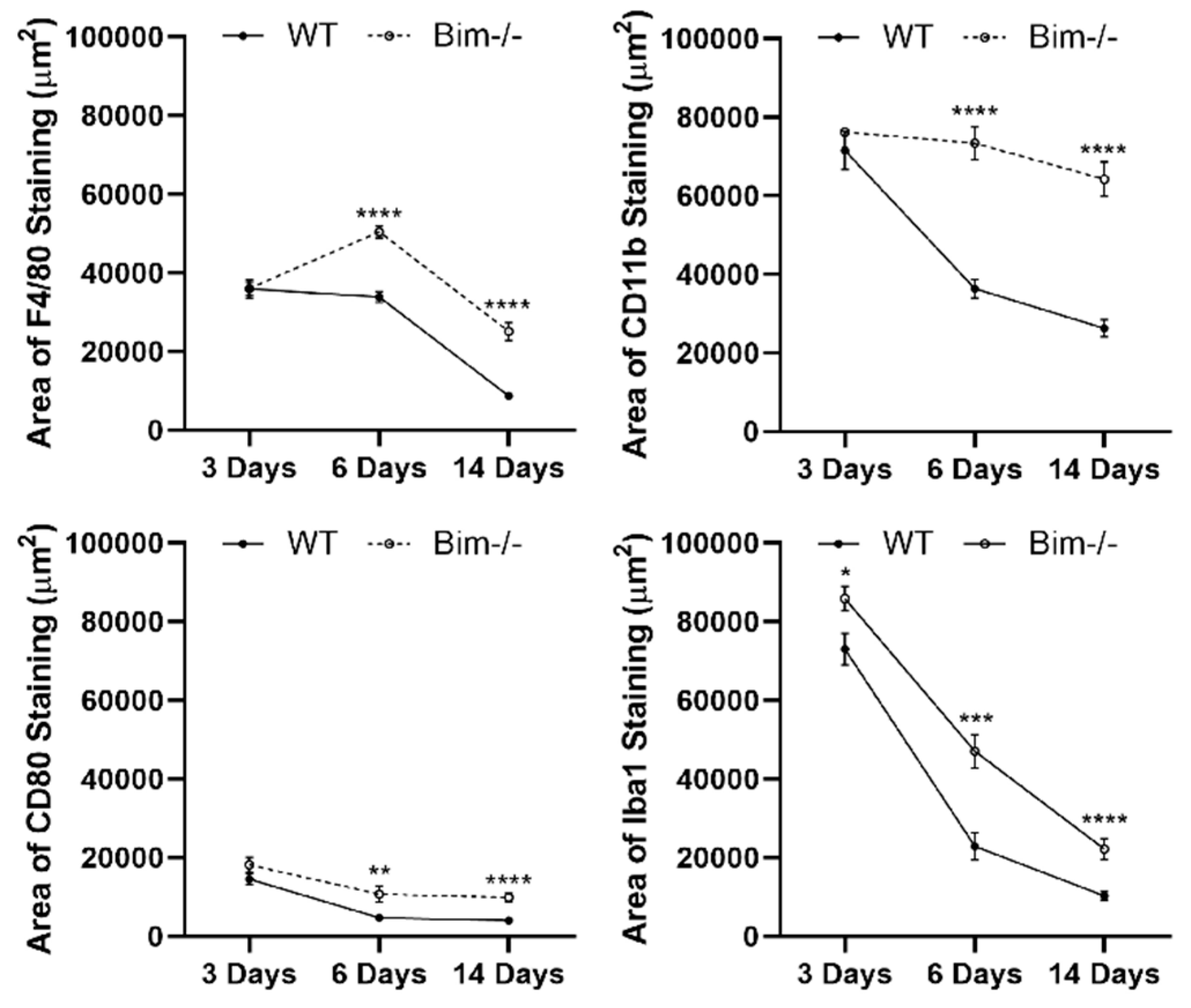
2.1. Bim Modulates Resolution of F4/80-Positive Staining Cells
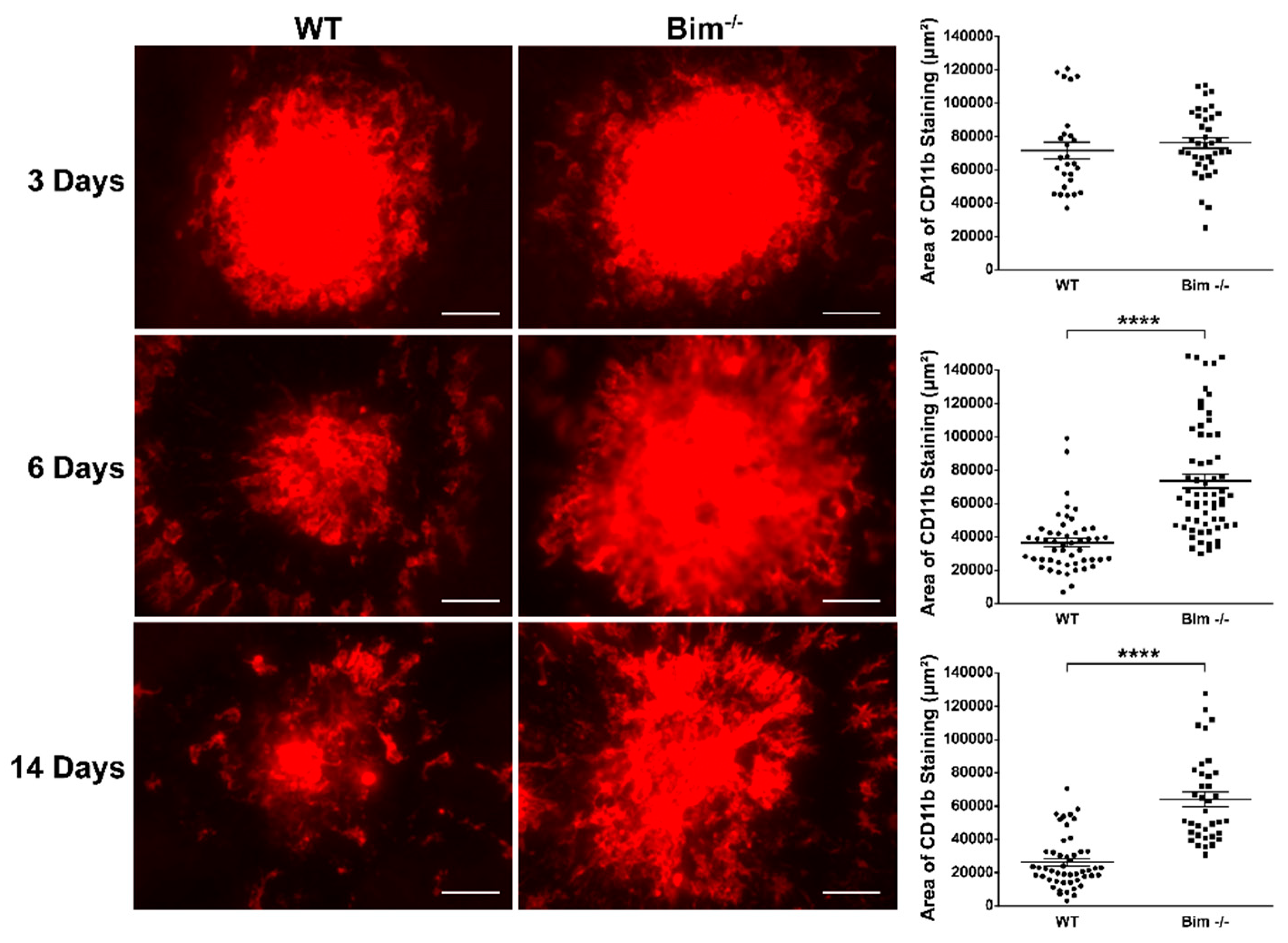
2.2. Bim Expression Aids Resolution of CD11b-Positive Staining Cells
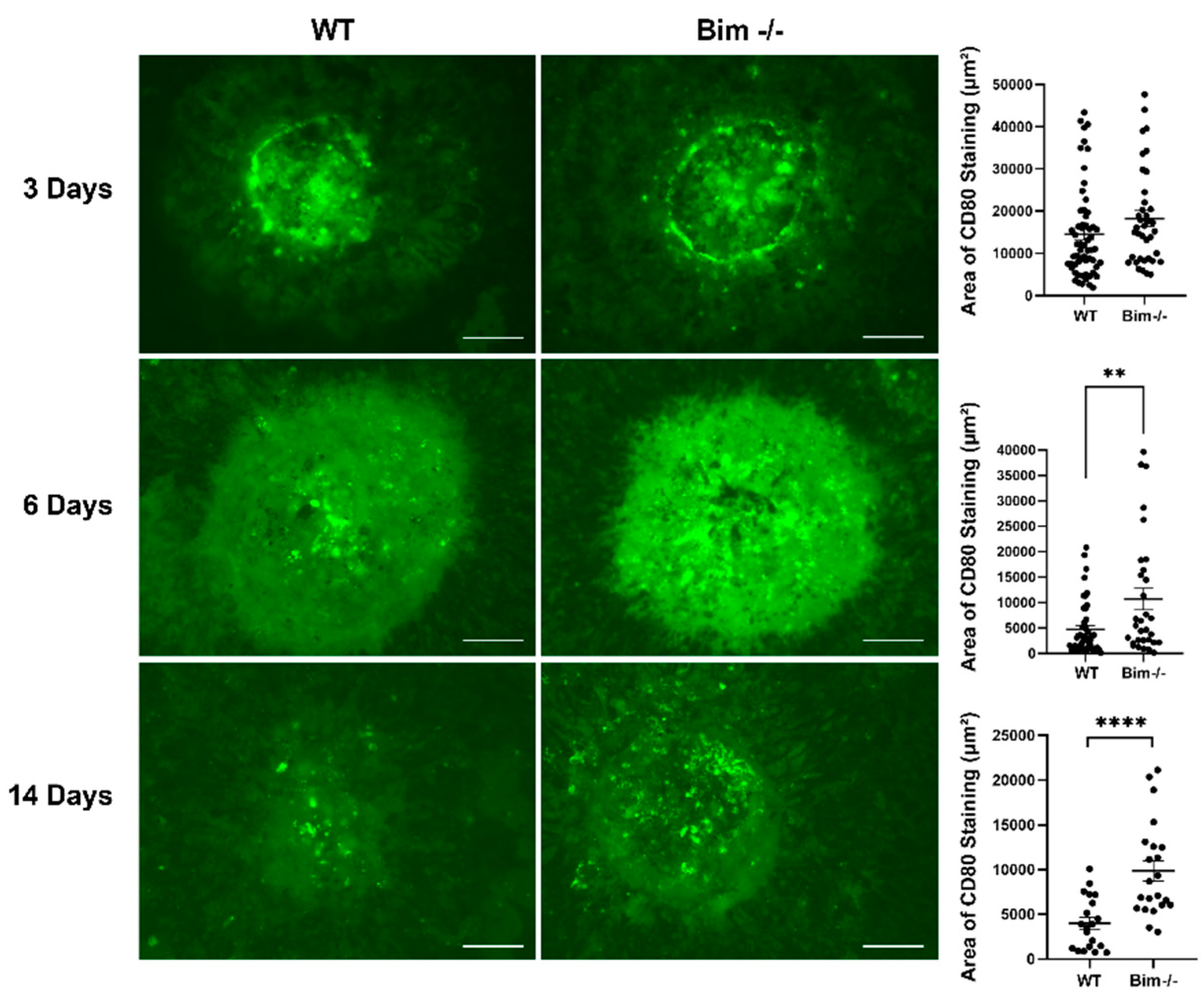
2.3. Delayed Clearing of CD80-Positive Staining Cells in the Absence of Bim
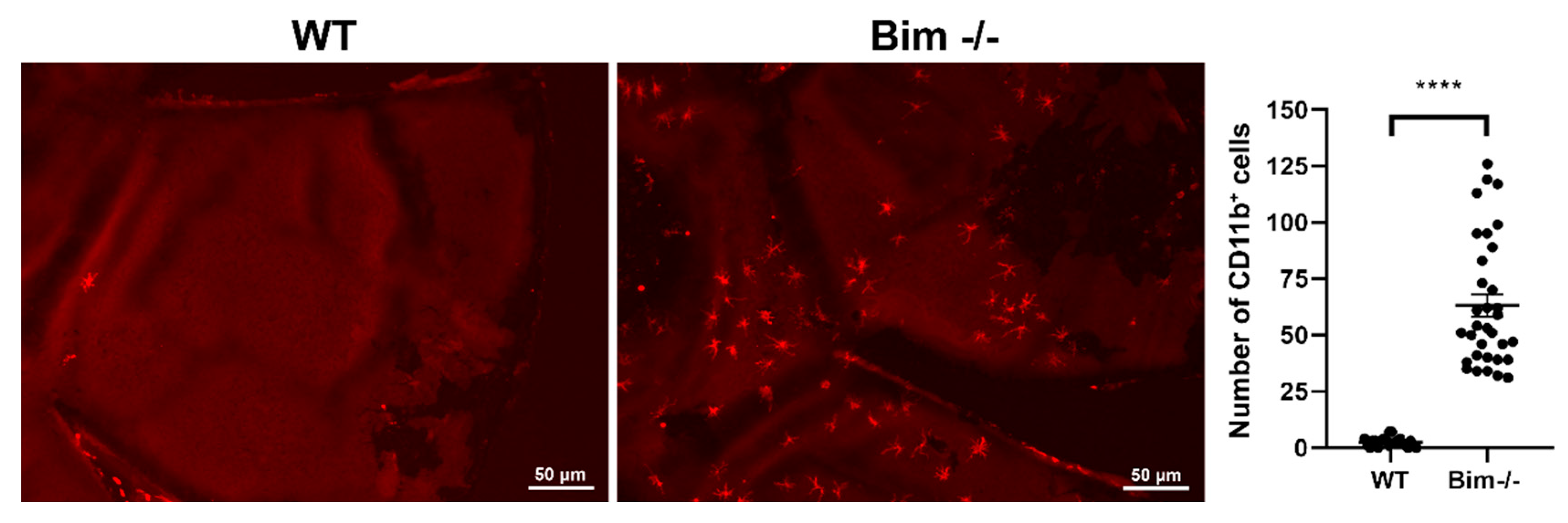
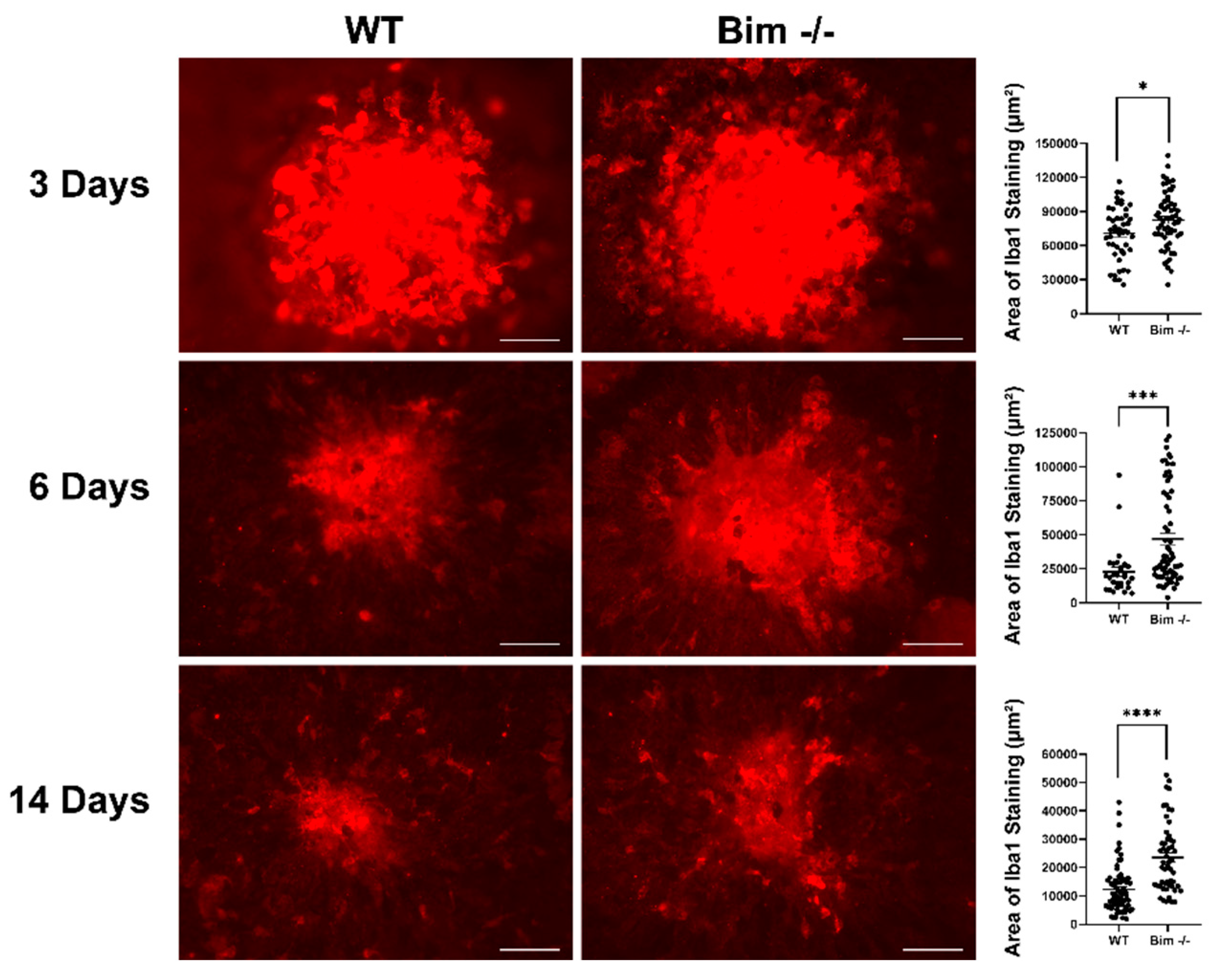
2.4. Delayed Resolution of Iba1-Positive Cells in the Absence of Bim
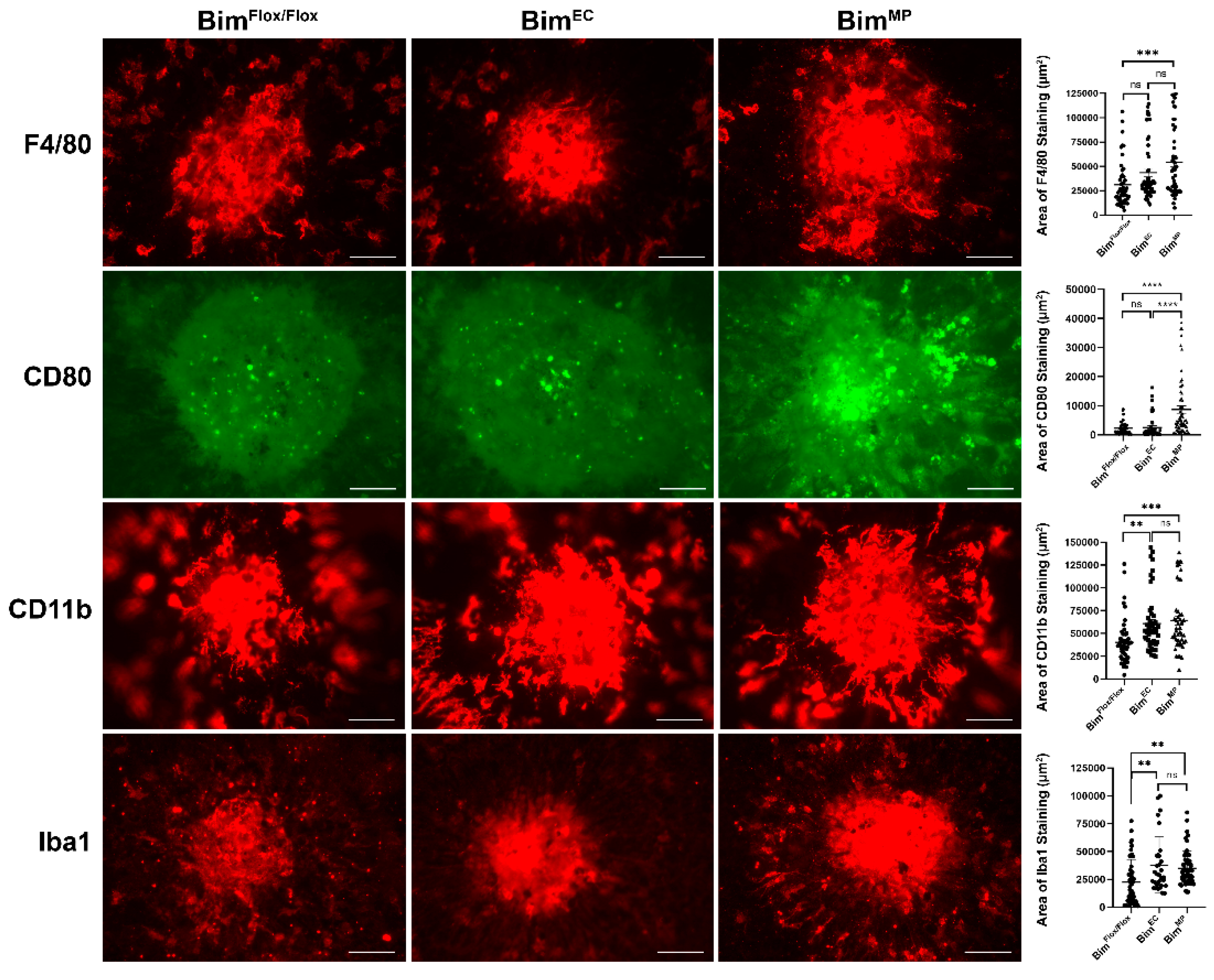
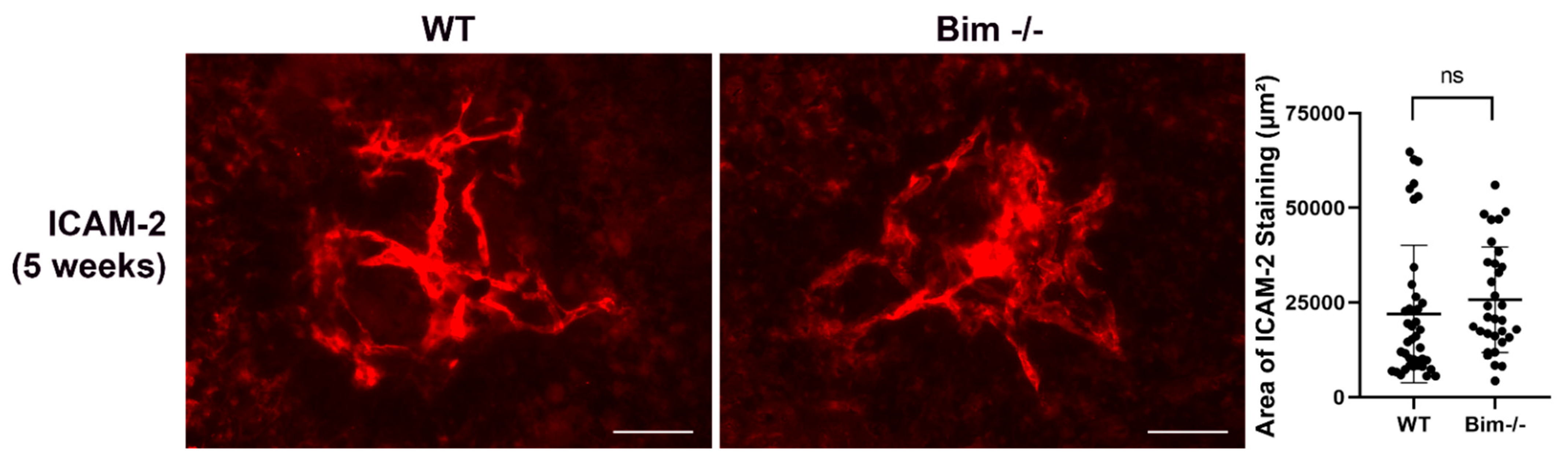
2.5. Lack of Bim Expression in Endothelial Cells or Mononuclear Phagocytes
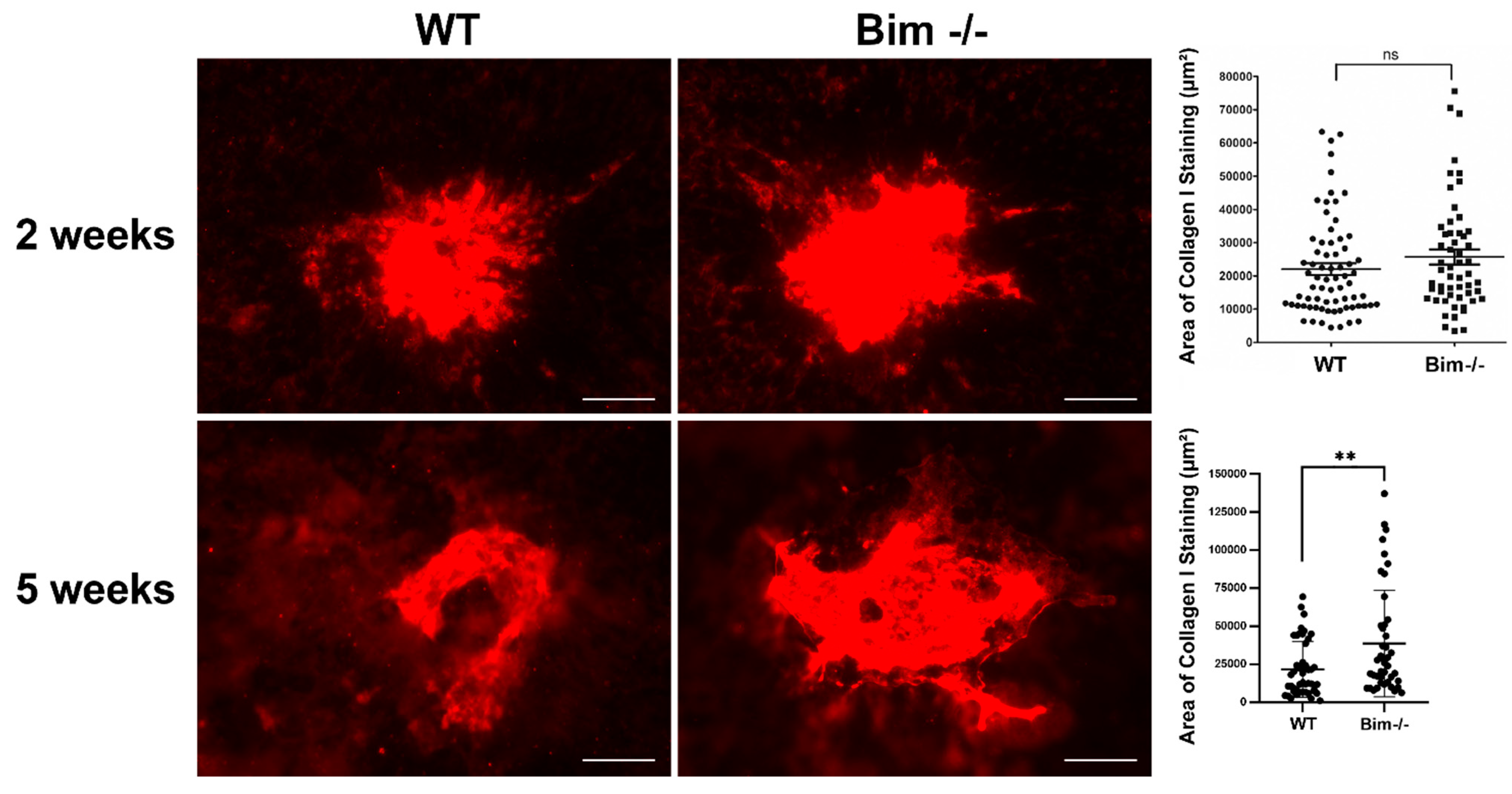
2.6. Increased Scar Formation in the Global Absence of Bim
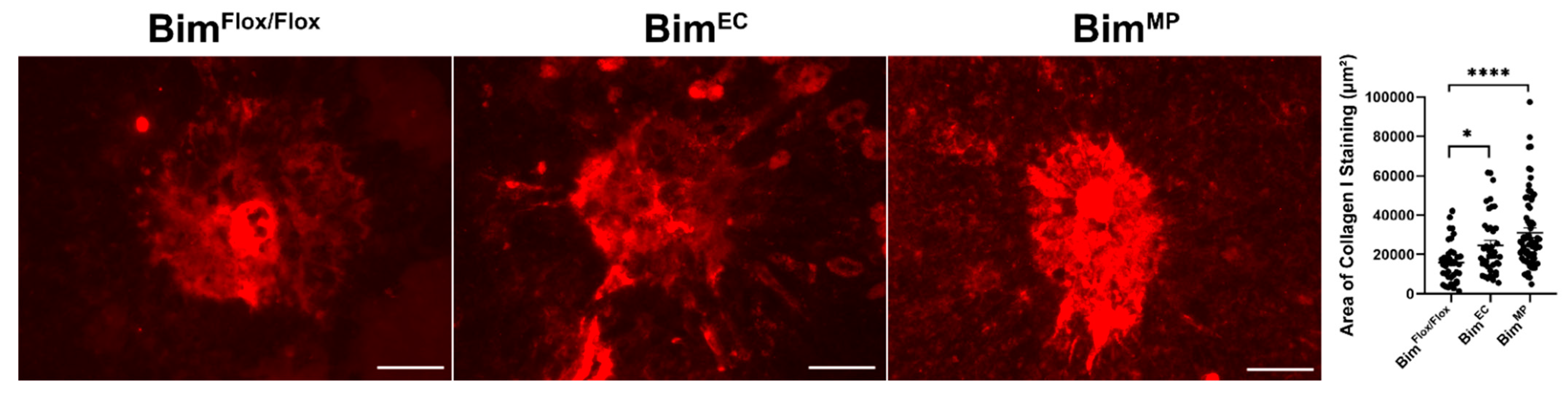
2.7. Lack of Endothelial Cell or MP Bim Expression Influences Scar Accumulation
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Laser-Induced Choroidal Neovascularization (CNV)
4.3. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jager, R.D.; Mieler, W.F.; Miller, J.W. Age-related macular degeneration. N. Engl. J. Med. 2008, 358, 2606–2617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pemp, B.; Schmetterer, L. Ocular blood flow in diabetes and age-related macular degeneration. Can. J. Ophthalmol. 2008, 43, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Farnoodian, M.; Wang, S.; Dietz, J.; Nickells, R.W.; Sorenson, C.M.; Sheibani, N. Negative regulators of angiogenesis: Important targets for treatment of exudative amd. Clin. Sci. 2017, 131, 1763–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmeliet, P.; Ferreira, V.; Breier, G.; Pollefeyt, S.; Kieckens, L.; Gertsenstein, M.; Fahrig, M.; Vandenhoeck, A.; Harpal, K.; Eberhardt, C.; et al. Abnormal blood vessel development and lethality in embryos lacking a single vegf allele. Nature 1996, 380, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Carver-Moore, K.; Chen, H.; Dowd, M.; Lu, L.; O’Shea, K.S.; Powell-Braxton, L.; Hillan, K.J.; Moore, M.W. Heterozygous embryonic lethality induced by targeted inactivation of the vegf gene. Nature 1996, 380, 439–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonoda, K.H.; Sasa, Y.; Qiao, H.; Tsutsumi, C.; Hisatomi, T.; Komiyama, S.; Kubota, T.; Sakamoto, T.; Kawano, Y.; Ishibashi, T. Immunoregulatory role of ocular macrophages: The macrophages produce rantes to suppress experimental autoimmune uveitis. J. Immunol. 2003, 171, 2652–2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz-Guilloty, F.; Saeed, A.M.; Echegaray, J.J.; Duffort, S.; Ballmick, A.; Tan, Y.; Betancourt, M.; Viteri, E.; Ramkhellawan, G.C.; Ewald, E.; et al. Infiltration of proinflammatory m1 macrophages into the outer retina precedes damage in a mouse model of age-related macular degeneration. Int. J. Inflamm. 2013, 2013, 503725. [Google Scholar] [CrossRef] [PubMed]
- Sennlaub, F.; Auvynet, C.; Calippe, B.; Lavalette, S.; Poupel, L.; Hu, S.J.; Dominguez, E.; Camelo, S.; Levy, O.; Guyon, E.; et al. Ccr2(+) monocytes infiltrate atrophic lesions in age-related macular disease and mediate photoreceptor degeneration in experimental subretinal inflammation in cx3cr1 deficient mice. EMBO Mol. Med. 2013, 5, 1775–1793. [Google Scholar] [CrossRef]
- Tsutsumi, C.; Sonoda, K.H.; Egashira, K.; Qiao, H.; Hisatomi, T.; Nakao, S.; Ishibashi, M.; Charo, I.F.; Sakamoto, T.; Murata, T.; et al. The critical role of ocular-infiltrating macrophages in the development of choroidal neovascularization. J. Leukoc. Biol. 2003, 74, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Sorenson, C.M.; Sheibani, N. Lack of thrombospondin 1 and exacerbation of choroidal neovascularization. Arch. Ophthalmol. 2012, 130, 615–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Park, S.; Fei, P.; Sorenson, C.M. Bim is responsible for the inherent sensitivity of the developing retinal vasculature to hyperoxia. Dev. Biol. 2011, 349, 296–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Zaitoun, I.S.; Johnson, R.P.; Jamali, N.; Gurel, Z.; Wintheiser, C.M.; Strasser, A.; Lindner, V.; Sheibani, N.; Sorenson, C.M. Bim expression in endothelial cells and pericytes is essential for regression of the fetal ocular vasculature. PLoS ONE 2017, 12, e0178198. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, L.M.; Nassin, M.L.; Hadji, A.; LaBelle, J.L. Killing two cells with one stone: Pharmacologic bcl-2 family targeting for cancer cell death and immune modulation. Front. Pediatrics 2016, 4, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herold, M.J.; Stuchbery, R.; Merino, D.; Willson, T.; Strasser, A.; Hildeman, D.; Bouillet, P. Impact of conditional deletion of the pro-apoptotic bcl-2 family member bim in mice. Cell Death Dis. 2014, 5, e1446. [Google Scholar] [CrossRef] [Green Version]
- Alfredsson, J.; Puthalakath, H.; Martin, H.; Strasser, A.; Nilsson, G. Proapoptotic bcl-2 family member bim is involved in the control of mast cell survival and is induced together with bcl-xl upon ige-receptor activation. Cell Death Differ. 2005, 12, 136–144. [Google Scholar] [CrossRef] [Green Version]
- Tsai, F.; Homan, P.J.; Agrawal, H.; Misharin, A.V.; Abdala-Valencia, H.; Haines, G.K., 3rd; Dominguez, S.; Bloomfield, C.L.; Saber, R.; Chang, A.; et al. Bim suppresses the development of sle by limiting myeloid inflammatory responses. J. Exp. Med. 2017, 214, 3753–3773. [Google Scholar] [CrossRef] [PubMed]
- Gautier, E.L.; Ivanov, S.; Lesnik, P.; Randolph, G.J. Local apoptosis mediates clearance of macrophages from resolving inflammation in mice. Blood 2013, 122, 2714–2722. [Google Scholar] [CrossRef] [Green Version]
- Dupuy, A.G.; Caron, E. Integrin-dependent phagocytosis: Spreading from microadhesion to new concepts. J. Cell Sci. 2008, 121, 1773–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faridi, M.H.; Khan, S.Q.; Zhao, W.; Lee, H.W.; Altintas, M.M.; Zhang, K.; Kumar, V.; Armstrong, A.R.; Carmona-Rivera, C.; Dorschner, J.M.; et al. Cd11b activation suppresses tlr-dependent inflammation and autoimmunity in systemic lupus erythematosus. J. Clin. Investig. 2017, 127, 1271–1283. [Google Scholar] [CrossRef] [Green Version]
- Han, C.; Jin, J.; Xu, S.; Liu, H.; Li, N.; Cao, X. Integrin cd11b negatively regulates tlr-triggered inflammatory responses by activating syk and promoting degradation of myd88 and trif via cbl-b. Nat. Immunol. 2010, 11, 734–742. [Google Scholar] [CrossRef]
- Means, T.K.; Luster, A.D. Integrins limit the toll. Nat. Immunol. 2010, 11, 691–693. [Google Scholar] [CrossRef] [PubMed]
- Phillipson, M.; Heit, B.; Colarusso, P.; Liu, L.; Ballantyne, C.M.; Kubes, P. Intraluminal crawling of neutrophils to emigration sites: A molecularly distinct process from adhesion in the recruitment cascade. J. Exp. Med. 2006, 203, 2569–2575. [Google Scholar] [CrossRef] [PubMed]
- Gahmberg, C.G. Leukocyte adhesion: Cd11/cd18 integrins and intercellular adhesion molecules. Curr. Opin. Cell Biol. 1997, 9, 643–650. [Google Scholar] [CrossRef]
- Schmid, M.C.; Avraamides, C.J.; Foubert, P.; Shaked, Y.; Kang, S.W.; Kerbel, R.S.; Varner, J.A. Combined blockade of integrin-α4β1 plus cytokines sdf-1α or il-1β potently inhibits tumor inflammation and growth. Cancer Res. 2011, 71, 6965–6975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Mills, C.D.; Kincaid, K.; Alt, J.M.; Heilman, M.J.; Hill, A.M. M-1/m-2 macrophages and the th1/th2 paradigm. J. Immunol. 2000, 164, 6166–6173. [Google Scholar] [CrossRef] [Green Version]
- Scholz, R.; Caramoy, A.; Bhuckory, M.B.; Rashid, K.; Chen, M.; Xu, H.; Grimm, C.; Langmann, T. Targeting translocator protein (18 kda) (tspo) dampens pro-inflammatory microglia reactivity in the retina and protects from degeneration. J. Neuroinflamm. 2015, 12, 201. [Google Scholar] [CrossRef] [Green Version]
- Lull, M.E.; Block, M.L. Microglial activation and chronic neurodegeneration. Neurother. J. Am. Soc. Exp. Neurother. 2010, 7, 354–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirschnek, S.; Ying, S.; Fischer, S.F.; Häcker, H.; Villunger, A.; Hochrein, H.; Häcker, G. Phagocytosis-induced apoptosis in macrophages is mediated by up-regulation and activation of the bcl-2 homology domain 3-only protein bim. J. Immunol. 2005, 174, 671–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scatizzi, J.C.; Hutcheson, J.; Pope, R.M.; Firestein, G.S.; Koch, A.E.; Mavers, M.; Smason, A.; Agrawal, H.; Haines, G.K., 3rd; Chandel, N.S.; et al. Bim-bcl-2 homology 3 mimetic therapy is effective at suppressing inflammatory arthritis through the activation of myeloid cell apoptosis. Arthritis Rheum. 2010, 62, 441–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeretssian, G.; Correa, R.G.; Doiron, K.; Fitzgerald, P.; Dillon, C.P.; Green, D.R.; Reed, J.C.; Saleh, M. Non-apoptotic role of bid in inflammation and innate immunity. Nature 2011, 474, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Hutcheson, J.; Perlman, H. Bh3-only proteins in rheumatoid arthritis: Potential targets for therapeutic intervention. Oncogene 2008, 27 (Suppl. 1), S168–S175. [Google Scholar] [CrossRef] [Green Version]
- Hutcheson, J.; Scatizzi, J.C.; Siddiqui, A.M.; Haines, G.K., 3rd; Wu, T.; Li, Q.Z.; Davis, L.S.; Mohan, C.; Perlman, H. Combined deficiency of proapoptotic regulators bim and fas results in the early onset of systemic autoimmunity. Immunity 2008, 28, 206–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nopora, A.; Brocker, T. Bcl-2 controls dendritic cell longevity in vivo. J. Immunol. 2002, 169, 3006–3014. [Google Scholar] [CrossRef] [Green Version]
- Hou, W.S.; Van Parijs, L. A bcl-2-dependent molecular timer regulates the lifespan and immunogenicity of dendritic cells. Nat. Immunol. 2004, 5, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Gautier, E.L.; Huby, T.; Saint-Charles, F.; Ouzilleau, B.; Chapman, M.J.; Lesnik, P. Enhanced dendritic cell survival attenuates lipopolysaccharide-induced immunosuppression and increases resistance to lethal endotoxic shock. J. Immunol. 2008, 180, 6941–6946. [Google Scholar] [CrossRef] [Green Version]
- Sorenson, C.M.; Wang, S.; Darjatmoko, S.R.; Gurel, Z.; Liu, B.; Sheibani, N. Targeted thrombospondin-1 expression in ocular vascular development and neovascularization. Front. Cell Dev. Biol. 2021, 9, 671989. [Google Scholar] [CrossRef] [PubMed]
- Augis, V.; Airiau, K.; Josselin, M.; Turcq, B.; Mahon, F.X.; Belloc, F. A single nucleotide polymorphism in cbim is associated with a slower achievement of major molecular response in chronic myeloid leukaemia treated with imatinib. PLoS ONE 2013, 8, e78582. [Google Scholar] [CrossRef] [PubMed]
- Plesa, M.; Gagné, V.; Glisovic, S.; Younan, M.; Sharif-Askari, B.; Laverdière, C.; Alos, N.; Leclerc, J.M.; Sallan, S.E.; Neuberg, D.; et al. Influence of bcl2l11 polymorphism on osteonecrosis during treatment of childhood acute lymphoblastic leukemia. Pharm. J. 2019, 19, 33–41. [Google Scholar]
- Cardona, A.F.; Rojas, L.; Wills, B.; Arrieta, O.; Carranza, H.; Vargas, C.; Otero, J.; Corrales-Rodriguez, L.; Martín, C.; Reguart, N.; et al. Bim deletion polymorphisms in hispanic patients with non-small cell lung cancer carriers of egfr mutations. Oncotarget 2016, 7, 68933–68942. [Google Scholar] [CrossRef] [Green Version]
- Ying, H.Q.; Chen, J.; He, B.S.; Pan, Y.Q.; Wang, F.; Deng, Q.W.; Sun, H.L.; Liu, X.; Wang, S.K. The effect of bim deletion polymorphism on intrinsic resistance and clinical outcome of cancer patient with kinase inhibitor therapy. Sci. Rep. 2015, 5, 11348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alva, J.A.; Zovein, A.C.; Monvoisin, A.; Murphy, T.; Salazar, A.; Harvey, N.L.; Carmeliet, P.; Iruela-Arispe, M.L. Ve-cadherin-cre-recombinase transgenic mouse: A tool for lineage analysis and gene deletion in endothelial cells. Dev. Dyn. 2006, 235, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Corada, M.; Nyqvist, D.; Orsenigo, F.; Caprini, A.; Giampietro, C.; Taketo, M.M.; Iruela-Arispe, M.L.; Adams, R.H.; Dejana, E. The wnt/beta-catenin pathway modulates vascular remodeling and specification by upregulating dll4/notch signaling. Dev. Cell 2010, 18, 938–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuttler, A.S.; LeClair, R.J.; Stohn, J.P.; Wang, Q.; Sorenson, C.M.; Liaw, L.; Lindner, V. Characterization of pdgfrb-cre transgenic mice reveals reduction of rosa26 reporter activity in remodeling arteries. Genesis 2011, 49, 673–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckle, T.; Hughes, K.; Ehrentraut, H.; Brodsky, K.S.; Rosenberger, P.; Choi, D.S.; Ravid, K.; Weng, T.; Xia, Y.; Blackburn, M.R.; et al. Crosstalk between the equilibrative nucleoside transporter ent2 and alveolar adora2b adenosine receptors dampens acute lung injury. FASEB J. 2013, 27, 3078–3089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Economopoulou, M.; Langer, H.F.; Celeste, A.; Orlova, V.V.; Choi, E.Y.; Ma, M.; Vassilopoulos, A.; Callen, E.; Deng, C.; Bassing, C.H.; et al. Histone h2ax is integral to hypoxia-driven neovascularization. Nat. Med. 2009, 15, 553–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, J.J.; Briot, A.; Enciso, J.; Zovein, A.C.; Ren, S.; Zhang, Z.W.; Radtke, F.; Simons, M.; Wang, Y.; Iruela-Arispe, M.L. Endothelial deletion of murine jag1 leads to valve calcification and congenital heart defects associated with alagille syndrome. Development 2012, 139, 4449–4460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Lee, D.K. What is the proper way to apply the multiple comparison test? Korean J. Anesthesiol. 2018, 71, 353–360. [Google Scholar] [CrossRef] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Zaitoun, I.S.; Darjatmoko, S.R.; Sheibani, N.; Sorenson, C.M. Bim Expression Promotes the Clearance of Mononuclear Phagocytes during Choroidal Neovascularization, Mitigating Scar Formation in Mice. Life 2022, 12, 208. https://doi.org/10.3390/life12020208
Wang S, Zaitoun IS, Darjatmoko SR, Sheibani N, Sorenson CM. Bim Expression Promotes the Clearance of Mononuclear Phagocytes during Choroidal Neovascularization, Mitigating Scar Formation in Mice. Life. 2022; 12(2):208. https://doi.org/10.3390/life12020208
Chicago/Turabian StyleWang, Shoujian, Ismail S. Zaitoun, Soesiawati R. Darjatmoko, Nader Sheibani, and Christine M. Sorenson. 2022. "Bim Expression Promotes the Clearance of Mononuclear Phagocytes during Choroidal Neovascularization, Mitigating Scar Formation in Mice" Life 12, no. 2: 208. https://doi.org/10.3390/life12020208