
Anemia in Sports: A Narrative Review

 , ,
, ,
Abstract
:1. Introduction
2. Sports Anemia
3. Iron Deficiency
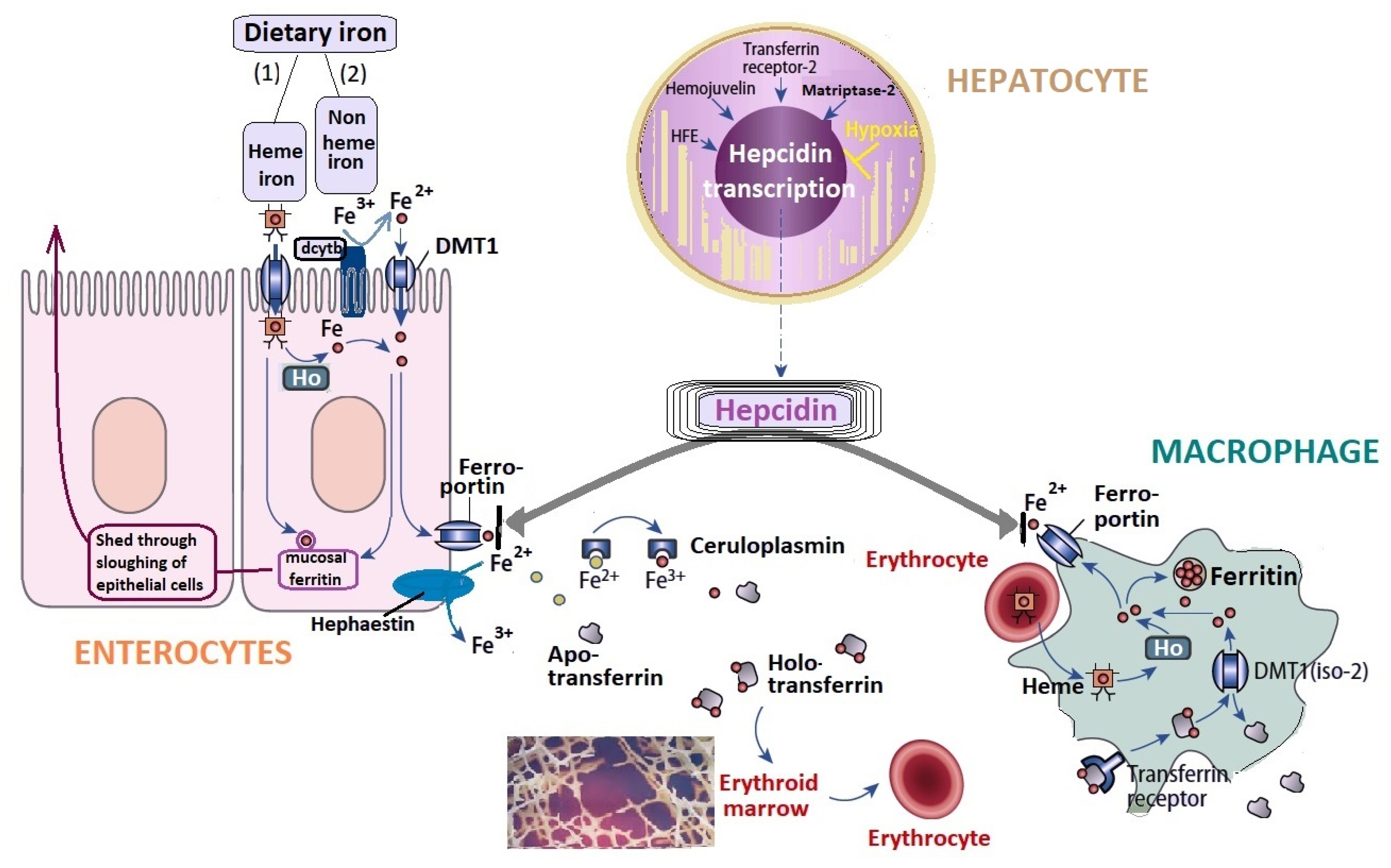
3.1. Iron Metabolism
3.2. Non Genetic Factors That Influence Iron Metabolism
3.2.1. Iron Absorption
3.2.2. Iron Loss during Exercise
3.2.3. Gastrointestinal Diseases
3.2.4. Inflammation
3.2.5. Other Losses
4. Sport-Related Hemolytic Anemia
5. Genetics, Sport, and Anemia
6. Other Considerations
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Banfi, G. Biochemical and Haematological Parameters in Football Players. In Football Traumatology, 2nd ed.; Volpi, P., Ed.; Springer International Publishing: Cham, Switzerland, 2006; pp. 43–52. [Google Scholar] [CrossRef]
- Eichner, E.R. Sports anemia, iron supplements, and blood doping. Med. Sci. Sports Exerc. 1992, 9, S315–S318. [Google Scholar] [CrossRef]
- Silva, A.S.R.; Santhiago, V.; Papoti, M.; Gobatto, C. Hematological parameters and anaerobic threshold in Brazilian soccer players throughout a training program. Int. J. Lab. Hematol. 2008, 30, 158–166. [Google Scholar] [CrossRef]
- Schumacher, Y.O.; Schmid, A.; Grathwohl, D.; Bültermann, D.; Berg, A. Hematological indices and iron status in athletes of various sports and performances. Med. Sci. Sports Exerc. 2002, 34, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Weight, L.M.; Klein, M.; Noakes, T.D.; Jacobs, P. “Sports anemia”—A real or apparent phenomenon in endurance-trained athletes? Int. J. Sports Med. 1992, 13, 344–347. [Google Scholar] [CrossRef]
- Clénin, G.; Cordes, M.; Huber, A.; Schumacher, Y.O.; Noack, P.; Scales, J.; Kriemler, S. Iron deficiency in sports—Definition, influence on performance and therapy. Schweiz. Z. Fur. Sport Und Sport 2016, 64, 6–18. [Google Scholar] [CrossRef] [Green Version]
- DellaValle, D.M. Iron supplementation for female athletes: Effects on iron status and performance outcomes. Curr Sports Med. Rep. 2013, 12, 234–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielik, V.; Kolisek, M. Bioaccessibility and bioavailability of minerals in relation to a healthy gut microbiome. Int. J. Mol. Sci. 2021, 22, 6803. [Google Scholar] [CrossRef] [PubMed]
- Varamenti, E.; Nikolovski, Z.; Elgingo, M.I.; Jamurtas, A.Z.; Cardinale, M. Training-Induced Variations in Haematological and Biochemical Variables in Adolescent Athletes of Arab Origin Throughout an Entire Athletic Season. J. Hum. Kinet 2018, 64, 123–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joksimović, A.; Stanković, D.; Ilić, D.; Joksimović, I.; Jerkan, M. Hematological profile of Serbian Youth National Soccer Teams. J. Hum. Kinet 2009, 22, 51–59. [Google Scholar] [CrossRef]
- Koc, H.; Tekin, A.; Ozturk, A.; Sarayemen, R.; Gokdemir, K.E.M. The effect of acute exercises on blood hematological parameters in handball players. Afr. J. Microbiol Res. 2012, 6, 2027–2032. [Google Scholar] [CrossRef]
- Buyukyazi, G.; Turgay, F. Acute and chronic effects of continuous and extensive interval running exercises on some haematalogical parameters. Turk. J. Sports Med. 2000, 35, 103–113. [Google Scholar]
- Bussollaro, A.K.; de Freitas, B.; Fadel, R.M.; Lara, G.S.; Barbosa, P.D.; Mezzomo, T.R. Hematological parameters and body composition of soccer players. Rev. Bras. Med. Esporte 2018, 24, 202–205. [Google Scholar] [CrossRef] [Green Version]
- Liao, P.; He, Q.; Zhou, X.; Ma, K.; Wen, J.; Chen, H.; Li, Q.; Qin, D.; Wang, H. Repetitive Bouts of Exhaustive Exercise Induces a Systemic Inflammatory Response and Multi-Organ Damage in Rats. Front. Physiol 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Alves, A.J.; Viana, J.; Cavalcante, S.L.; Oliveira, N.L.; Duarte, J.A.; Mota, J.; Oliveira, J.; Ribeiro, F. Physical activity in primary and secondary prevention of cardiovascular disease: Overview updated. World J. Cardiol. 2016, 8, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Semenova, E.A.; Miyamoto-Mikami, E.; Akimov, E.B.; Al-Khelaifi, F.; Murakami, H.; Zempo, H.; Kostryukova, E.S.; Kulemin, N.A.; Larin, A.K.; Borisov, O.V.; et al. The association of HFE gene H63D polymorphism with endurance athlete status and aerobic capacity: Novel findings and a meta-analysis. Graefe’s Arch. Clin. Exp. Ophthalmol. 2020, 120, 665–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsuphrom, P.; Latunde-Dada, G.O. Association of serum hepcidin levels with aerobic and resistance exercise: A systematic review. Nutrients 2021, 13, 393. [Google Scholar] [CrossRef] [PubMed]
- Hinton, P.S. Iron and the endurance athlete. Appl. Physiol. Nutr. Metab. 2014, 39, 1012–1018. [Google Scholar] [CrossRef]
- Kong, W.N.; Gao, G.; Chang, Y.Z. Hepcidin and sports anemia. Cell Biosci. 2014, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Sim, M.; Garvican-Lewis, L.A.; Cox, G.; Govus, A.; McKay, A.K.A.; Stellingwerff, T.; Peeling, P. Iron considerations for the athlete: A narrative review. Graefe’s Arch. Clin. Exp. Ophthalmol. 2019, 119, 1463–1478. [Google Scholar] [CrossRef]
- Kawamori, N.; Haff, G. The optimal training load for the development of muscular power. J. Strength Cond. Res. 2004, 18, 675–684. [Google Scholar] [PubMed]
- Domínguez, R.; Sánchez-Oliver, A.J.; Mata-Ordoñez, F.; Feria-Madueño, A.; Grimaldi-Puyana, M.; López-Samanes, Á.; Pérez-López, A. Effects of an Acute Exercise Bout on Serum Hepcidin Levels. Nutrients 2018, 10, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hasselt, P.M.; Clayton, P.H.R. Disorders in the Transport of Copper, Iron, Magnesium. In Inborn Metabolic Diseases, 6th ed.; Saudubray, J.M., Baumgartner, M.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 531–548. [Google Scholar]
- Kowdley, K.V.; Brown, K.E.; Ahn, J.; Sundaram, V. ACG Clinical Guideline: Hereditary Hemochromatosis. Clin. Liver Dis. 2020, 16, 177. [Google Scholar] [CrossRef]
- Rose, R.M. Overview of endocrinology of stress. In Neuroendocrinology and Psychiatric Disorder; Brown, G.M., Koslow, S.H., Reichlin, S., Eds.; Raven Press: New York, NY, USA, 1984; pp. 95–1223. [Google Scholar]
- González-Domínguez, Á.; Visiedo-García, F.M.; Domínguez-Riscart, J.; González-Domínguez, R.; Mateos, R.M.; Lechuga-Sancho, A.M. Iron metabolism in obesity and metabolic syndrome. Int. J. Mol. Sci. 2020, 21, 5529. [Google Scholar] [CrossRef]
- Pietrangelo, A. Genetics, Genetic Testing, and Management of Hemochromatosis: 15 Years since Hepcidin. Gastroenterology 2015, 149, 1240–1251.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jończy, A.; Mazgaj, R.; Starzyński, R.R.; Poznański, P.; Szudzik, M.; Smuda, E.; Kamyczek, M.; Lipiński, P. Relationship between down-regulation of copper-related genes and decreased ferroportin protein level in the duodenum of iron-deficient piglets. Nutrients 2021, 13, 104. [Google Scholar] [CrossRef]
- Yang, Q.; Liu, W.; Zhang, S.; Liu, S. The cardinal roles of ferroportin and its partners in controlling cellular iron in and out. Life Sci. 2020, 258, 118135. [Google Scholar] [CrossRef] [PubMed]
- McKay, A.K.A.; Pyne, D.B.; Burke, L.M.; Peeling, P. Iron metabolism: Interactions with energy and carbohydrate availability. Nutr. 2020, 12, 3692. [Google Scholar] [CrossRef]
- Hoppe, M.; Hulthén, L.; Hallberg, L. The importance of bioavailability of dietary iron in relation to the expected effect from iron fortification. Eur. J. Clin. Nutr. 2008, 62, 761–769. [Google Scholar] [CrossRef] [Green Version]
- Gibson, R.S.; Heath, A.L.M.; Szymlek-Gay, E.A. Is iron and zinc nutrition a concern for vegetarian infants and young children in industrialized countries? Am. J. Clin. Nutr. 2014, 100, 459S–468S. [Google Scholar] [CrossRef] [Green Version]
- Rusu, I.G.; Vodnar, D.C.; Pop, C.R.; Sonia, A.S.; Vulturar, R.; Istrati, M.; Morosan, I.; Farcas, A.C.; Kerezsi, A.D.; Muresan, C.I.; et al. Iron Supplementation Influence on the Gut Microbiota and Probiotic Intake Effect in Iron Deficiency—A Literature-Based Review. Nutrients 2020, 12, 1993. [Google Scholar] [CrossRef] [PubMed]
- Sierra, A.P.R.; Oliveira, R.A.; Silva, E.D.; Lima, G.; Benetti, M.P.; Kiss, M.A.P.; Sierra, C.A.; Ghorayeb, N.; Seto, J.T.; Pesquero, J.B.; et al. Association between hematological parameters and iron metabolism response after marathon race and ACTN3 genotype. Front. Physiol. 2019, 10, 697. [Google Scholar] [CrossRef] [Green Version]
- Ottomano, C.; Franchini, M. Sports anaemia: Facts or fiction? Blood Transfus. 2012, 10, 252–254. [Google Scholar] [CrossRef]
- Waller, M.F. The effects of heat and exercise on sweat iron loss. Med. Sci. Sport Exerc. 1996, 28, 197–203. [Google Scholar] [CrossRef]
- Brune, M.; Magnusson, B.; Persson, H.; Hallberg, L. Iron losses in sweat. Am. J. Clin. Nutr. 1986, 43, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Urakami, S.; Ogawa, K.; Oka, S.; Hayashida, M.; Hagiwara, K.; Nagamoto, S.; Sakaguchi, K.; Yano, A.; Kurosawa, K.; Okaneya, T. Macroscopic hematuria caused by running-induced traumatic bladder mucosal contusions. IJU Case Rep. 2019, 2, 27–29. [Google Scholar] [CrossRef]
- Shephard, R.J. Exercise proteinuria and hematuria: Current knowledge and future directions. J. Sport Med. Phys. Fit. 2016, 56, 1060–1076. [Google Scholar]
- Rojas-Valverde, D.; Olcina, G.; Sánchez-Ureña, B.; Pino-Ortega, J.; Martínez-Guardado, I.; Timón, R. Proteinuria and bilirubinuria as potential risk indicators of acute kidney injury during running in outpatient settings. Medicina 2020, 56, 562. [Google Scholar] [CrossRef] [PubMed]
- Packer, N.; Hoffman-Goetz, L. Does physical activity affect quality of life, disease symptoms and immune measures in patients with inflammatory bowel disease? A systematic review. J. Sport Med. Phys. Fit. 2010, 50, 1–18. [Google Scholar]
- Eckert, K.G.; Abbasi-Neureither, I.; Köppel, M.; Huber, G. Structured physical activity interventions as a complementary therapy for patients with inflammatory bowel disease—A scoping review and practical implications. BMC Gastroenterol. 2019, 19, 1–12. [Google Scholar] [CrossRef]
- Costa, R.J.S.; Snipe, R.M.J.; Kitic, C.M.; Gibson, P.R. Systematic review: Exercise-induced gastrointestinal syndrome—implications for health and intestinal disease. Aliment. Pharmacol. Ther. 2017, 46, 246–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lis, D.; Ahuja, K.; Stellingwerff, T.; Kitic, C.M. Case Study: Utilizing a Low FODMAP Diet to Combat Exercise-Induced Gastrointestinal Symptoms. Int. J. Sport Nutr. Exerc. Metab. 2016, 26, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Castanier, C.; Bougault, V.; Teulier, C.; Jaffré, C.; Schiano-Lomoriello, S.; Vibarel-Rebot, N.; Villemain, A.; Rieth, N.; Le-Scanff, C.; Buisson, C.; et al. The Specificities of Elite Female Athletes: A Multidisciplinary Approach. Life 2021, 11, 622. [Google Scholar] [CrossRef] [PubMed]
- Diduch, B.K. Gastrointestinal Conditions in the Female Athlete. Clin. Sports Med. 2017, 36, 655–669. [Google Scholar] [CrossRef] [PubMed]
- D’Hondt, S.; Van Damme, T.; Malfait, F. Vascular phenotypes in nonvascular subtypes of the Ehlers-Danlos syndrome: A systematic review. Genet. Med. 2018, 20, 562–573. [Google Scholar] [CrossRef] [Green Version]
- Casey, M.C.; Robertson, I.; Waters, P.S.; Hanaghan, J.; Khan, W.; Barry, K. Non-operative management of diverticular perforation in a patient with suspected Ehlers-Danlos syndrome. Int. J. Surg. Case Rep. 2014, 5, 135–137. [Google Scholar] [CrossRef] [Green Version]
- Fikree, A.; Chelimsky, G.; Collins, H.; Kovacic, K.; Aziz, Q. Gastrointestinal involvement in the Ehlers–Danlos syndromes. Am. J. Med. Genet. Part. C Semin Med. Genet. 2017, 175, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Sim, M.; Dawson, B.; Landers, G.; Swinkels, D.W.; Tjalsma, H.; Trinder, D. Effect of exercise modality and intensity on post-exercise interleukin-6 and hepcidin levels. Int. J. Sport Nutr. Exerc. Metab. 2013, 23, 178–186. [Google Scholar] [CrossRef]
- Dahlquist, D.T.; Stellingwerff, T.; Dieter, B.P. Effects of macro- and micronutrients on exercise-induced hepcidin response in highly trained endurance athletes. Appl. Physiol. Nutr. Metab. Eff. 2017, 42, 1036–1043. [Google Scholar] [CrossRef] [Green Version]
- Peeling, P.; Dawson, B.; Goodman, C.; Landers, G.; Trinder, D. Athletic induced iron deficiency: New insights into the role of inflammation, cytokines and hormones. Eur. J. Appl. Physiol. 2008, 103, 381–391. [Google Scholar] [CrossRef]
- Bruinvels, G.; Burden, R.; Brown, N.; Richards, T.; Pedlar, C. The prevalence and impact of heavy menstrual bleeding (Menorrhagia) in elite and non-elite athletes. PLoS ONE 2016, 11, 1–8. [Google Scholar] [CrossRef]
- Carmichael, M.A.; Thomson, R.L.; Moran, L.J.; Wycherley, T.P. The impact of menstrual cycle phase on athletes’ performance: A narrative review. Int. J. Environ. Res. Public Health 2021, 18, 1667. [Google Scholar] [CrossRef] [PubMed]
- Larsen, B.; Morris, K.; Quinn, K.; Osborne, M.; Minahan, C. Practice does not make perfect: A brief view of athletes’ knowledge on the menstrual cycle and oral contraceptives. J. Sci. Med. Sport 2020, 23, 690–694. [Google Scholar] [CrossRef] [PubMed]
- Quinn, K.M.; Cox, A.J.; Roberts, L.; Pennell, E.N.; McKeating, D.R.; Fisher, J.J.; Perkins, A.V.; Minahan, C. Temporal changes in blood oxidative stress biomarkers across the menstrual cycle and with oral contraceptive use in active women. Graefe’s Arch. Clin. Exp. Ophthalmol. 2021, 121, 2607–2620. [Google Scholar]
- Oxfeldt, M.; Dalgaard, L.B.; Jørgensen, A.A. Hormonal Contraceptive Use, Menstrual Dysfunctions, and Self-Reported Side Effects in Elite Athletes in Denmark. Int. J. Sport Physiol. Perform. 2020, 15, 1377–1384. [Google Scholar] [CrossRef]
- Lippi, G.; Sanchis-Gomar, F. Epidemiological, biological and clinical update on exercise-induced hemolysis. Ann. Transl. Med. 2019, 7, 270. [Google Scholar] [CrossRef]
- Telford, R.D.; Sly, G.J.; Hahn, A.G.; Cunningham, R.B.; Bryant, C.; Smith, J.A. Footstrike is the major cause of hemolysis during running. J. Appl. Physiol. 2003, 94, 38–42. [Google Scholar] [CrossRef] [Green Version]
- Lippi, G.; Schena, F.; Salvagno, G.L.; Aloe, R.; Banfi, G.; Guidi, G.C. Foot-strike haemolysis after a 60-km ultramarathon. Blood Transfus. 2012, 10, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Tohori, M.; Ashida, T.; Kajiwara, N. Comparison of effects of vegetable protein diet and animal protein diet on the initiation of anemia during vigorous physical training (sports anemia) in dogs and rats. J. Nutr. Sci. Vitaminol. 1987, 33, 129–149. [Google Scholar] [CrossRef]
- Hoffman, M.D.; Stuempfle, K.J.; Fogard, K.; Hew-Butler, T.; Winger, J.; Weiss, R.H. Urine dipstick analysis for identification of runners susceptible to acute kidney injury following an ultramarathon. J. Sports Sci. 2013, 31, 20–31. [Google Scholar] [CrossRef]
- Lee, F.X.Z.; Houweling, P.J.; North, K.N.; Quinlan, K.G.R. How does α-actinin-3 deficiency alter muscle function? Mechanistic insights into ACTN3, the “gene for speed”. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 686–693. [Google Scholar] [CrossRef]
- Friedlander, S.M.; Herrmann, A.L.; Lowry, D.; Mepham, E.R.; Lek, M.; North, K.N.; Organ, C.L. ACTN3 Allele Frequency in Humans Covaries with Global Latitudinal Gradient. PLoS ONE 2013, 8, e52282. [Google Scholar] [CrossRef] [Green Version]
- Hollerer, I.; Bachmann, A.; Muckenthaler, M.U. Pathophysiological consequences and benefits of HFE mutations: 20 years of research. Haematologica 2017, 102, 809–817. [Google Scholar] [CrossRef] [Green Version]
- Ishiguroa, K.; Linb, Z.P.; Penkethb, P.G.; Shyamb, K.; Zhub, R.; Baumannb, R.P.; Zhub, Y.-L.; Sartorellib, A.C.; Rutherforda, T.J.; Ratnera, E.S. Distinct mechanisms of cell-kill by triapine and its terminally dimethylated derivative Dp44mT due to a loss or gain of activity of their copper(II) complexes. Biochem. Pharmacol. 2014, 91, 312–322. [Google Scholar] [CrossRef] [Green Version]
- Dara, R.C.; Sharma, R.B.H. Severe drug-induced immune hemolysis due to ceftriaxone. Asian J. Transfus. Sci. 2020, 14, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Betensky, M.; Witmer, C.; Fisher, M.J.; Nance, S.; Weiss, M.J.; Sesok-Pizzini, D.A. Immune hemolytic anemia with drug-induced antibodies to carboplatin and vincristine in a pediatric patient with an optic pathway glioma. Transfusion 2014, 54, 2901–2905. [Google Scholar] [CrossRef] [PubMed]
- Salama, A. Drug-induced immune hemolytic anemia. Expert Opin. Drug Saf. 2009, 8, 73–79. [Google Scholar] [CrossRef]
- Kim, Y.; Park, J.; Kim, M. Diagnostic approaches for inherited hemolytic anemia in the genetic era. Blood Res. 2017, 52, 84–94. [Google Scholar] [CrossRef]
- Sankaran, V.G.; Weiss, M.J. Anemia: Progress in molecular mechanisms and therapy. Nat. Med. 2015, 21, 221–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stellingwerff, T.; Peeling, P.; Garvican-Lewis, L.A.; Hall, R.; Koivisto-Mørk, A.E.; Heikura, I.; Burke, L.M. Nutrition and Altitude: Strategies to Enhance Adaptation, Improve Performance and Maintain Health: A Narrative Review. Sport Med. 2019, 49, 169–184. [Google Scholar] [CrossRef] [Green Version]
- Gore, C.J.; Clark, S.A.; Saunders, P.U. Nonhematological mechanisms of improved sea-level performance after hypoxic exposure. Med. Sci. Sports Exerc. 2007, 39, 1600–1609. [Google Scholar] [CrossRef]
- Hannon, M.P.; Flueck, J.L.; Gremeaux, V.; Place, N.; Kayser, B.; Donnelly, C. Key Nutritional Considerations for Youth Winter Sports Athletes to Optimize Growth, Maturation and Sporting Development. Front. Sport Act. Living 2021, 3, 1–9. [Google Scholar] [CrossRef]
- Hall, R.; Peeling, P.; Nemeth, E.; Bergland, D.; Mccluskey, W.T.P.; Stellingwerff, T. Single versus Split Dose of Iron Optimizes Hemoglobin Mass Gains at 2106 m Altitude. Med. Sci. Sports Exerc. 2019, 51, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Saunders, P.U.; Garvican-Lewis, L.A.; Chapman, R.F.; Périard, J.D. Special environments: Altitude and heat. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 210–219. [Google Scholar] [CrossRef] [Green Version]
- Barrett, K.E.; Barman, S.M.; Brooks, H.L. Ganong’s Review of Medical Physiology; McGraw Hill Education: New York, NY, USA, 2019; p. 326. [Google Scholar]
- Stellingwerff, T.; Heikura, I.A.; Meeusen, R.; Bermon, S.; Seiler, S.; Mountjoy, M.L.; Burke, L.M. Overtraining Syndrome (OTS) and Relative Energy Deficiency in Sport (RED-S): Shared Pathways, Symptoms and Complexities. Sport Med. 2021. [Google Scholar] [CrossRef] [PubMed]
- Cadegiani, F.A.; Kater, C.E. Hormonal aspects of overtraining syndrome: A systematic review. BMC Sports Sci. Med. Rehabil. 2017, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Borkowska, A.; Tomczyk, M.; Żychowska, M.; Pilis, W.; Zych, M.; Antosiewicz, J. Effect of 8-day fasting on leukocytes expression of genes and proteins involved in iron metabolism in healthy men. Int. J. Mol. Sci. 2021, 22, 3248. [Google Scholar] [CrossRef]
- Maughan, R.J.; Leiper, J.B.; Bartagi, Z.; Zrifi, R.; Zerguini, Y.; Dvorak, J. Effect of Ramadan fasting on some biochemical and haematological parameters in Tunisian youth soccer players undertaking their usual training and competition schedule. J. Sports Sci. 2008, 26, 37–41. [Google Scholar] [CrossRef]
- Nisenbaum, E.J.; Novikov, D.; Lui, Y.W. The Presence and Role of Iron in Mild Traumatic Brain Injury: An Imaging Perspective. J. Neurotrauma 2014, 31, 301–307. [Google Scholar] [CrossRef]
- Cialdella-Kam, L.; Kulpins, D.; Manore, M.M. Vegetarian, Gluten-Free, and Energy Restricted Diets in Female Athletes. Sports 2016, 4, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mountjoy, M.; Sundgot-Borgen, J.K.; Burke, L.M.; Ackerman, K.E.; Blauwet, C.; Constantini, N.; Lebrun, C.; Lundy, B.; Melin, A.; Meyer, N.L.; et al. IOC consensus statement on relative energy deficiency in sport (RED-S): 2018 update. Br. J. Sports Med. 2018, 52, 687–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logue, D.M.; Madigan, S.M.; Melin, A.; Delahunt, E.; Heinen, M.; Mc Donnell, S.-J.; Corish, C.A. Low Energy Availability in Athletes 2020: An Updated Narrative Review of Prevalence, Risk, Within-Day Energy Balance, Knowledge, and Impact on Sports Performance. Nutrients 2020, 12, 835. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Type of Anemia | Cause | Observations | References |
|---|---|---|---|
| “Sports anemia” | Hemodilution Redistribution (plasma volume expansion) | Controversial; misnomer Increased Hb mass is outpaced by plasma expansion | [2,3,4,5] |
| Increased iron demands | Increased tissue remodeling | Increased erythropoiesis and muscle hypertrophy | [4,34] |
| Iron reduced intake | RED S | Low energy availability surrogates correlate with hematological dysfunction | [84] |
| Restrictive diets | Dancers, gymnasts, etc. | [85] | |
| Iron reduced absorption | Vegetarian diets | Complexes with phytate, oxalate, phosphate, polyphenols decreasing absorption | [31,32,33] |
| Gastrointestinal blood flow redistribution | May result in malabsorption | [43] | |
| Exercise-induced inflammation | Increased IL-6 triggers hepcidin, consequently lowering the iron absorption | [50,51] | |
| Celiac disease | Iron malabsorption | [45,46] | |
| Iron sequestration | Inflammation | IL-6 triggered hepcidin contribute to reduced iron availability in acute exercise | [17,43,52] |
| Iron loss | Sweating | During thermoregulation | [36,37] |
| Hematuria | Bladder posterior wall repeatedly kicked against the fixed bladder neck during running, catecholamines, hypoxia, oxidative stress, lactate accumulation, increased glomerular permeability, renal ischemia | [38,39] | |
| Gastrointestinal bleeding | Decreased gastrointestinal tract blood flow from redistribution to muscles results in fecal blood loss | [41,43] | |
| Inflammation | Decreased gastrointestinal blow flow leads to intestinal ischemia, increased permeability, endotoxemia, and systemic inflammatory responses | [43,44] | |
| Trauma | Hematoma, bleeding in contact sports (boxing, etc.) | [82] | |
| Polymenorrhea | Heavy menstrual bleeding in more than 1/3 of the female athletes Oral contraceptives increase oxidative stress and inflammation | [53,54,55] [45,56] | |
| Connective tissue fragility | Hematomas, hemorrhoids complicated with bleeding, diverticular perforation, or other vascular complications in hypermobility syndromes (dances, gymnasts, etc.) | [47,48] | |
| Hemolysis | Footstrike—foot vessels compression and trauma during running Vascular contraction and red blood cells extravascular compression by muscle contraction or by kidney vasoconstriction Lipid profile changes with decreased cholesterol and increased lysolecithin increase osmotic fragility | [58,59] [40,58,60] [58,61] | |
| Genetic causes | ACTN3 polymorphisms | 577X alleles homozygosity results in α-actinin-3 deficiency and improved iron metabolism Detrimental for power exercise and sprinting, but beneficial for endurance activities | [34,63] |
| HFE polymorphisms | HFE mutation (C282Y, H63D, or S65C) increases iron supply to the physical performance | [65] | |
| Hemoglobinopathies, RBC enzymopathies | β-thalassemia, sickle cell anemia, and others are frequent in certain populations; increased hemolysis | [70,71] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damian, M.-T.; Vulturar, R.; Login, C.C.; Damian, L.; Chis, A.; Bojan, A. Anemia in Sports: A Narrative Review. Life 2021, 11, 987. https://doi.org/10.3390/life11090987
Damian M-T, Vulturar R, Login CC, Damian L, Chis A, Bojan A. Anemia in Sports: A Narrative Review. Life. 2021; 11(9):987. https://doi.org/10.3390/life11090987
Chicago/Turabian StyleDamian, Marc-Tudor, Romana Vulturar, Cristian Cezar Login, Laura Damian, Adina Chis, and Anca Bojan. 2021. "Anemia in Sports: A Narrative Review" Life 11, no. 9: 987. https://doi.org/10.3390/life11090987