
The Causal Relationship between Endothelin-1 and Hypertension: Focusing on Endothelial Dysfunction, Arterial Stiffness, Vascular Remodeling, and Blood Pressure Regulation


Abstract
:1. Introduction
2. Data on the Participation of ET-1 in the Development of HTN
2.1. ET-1 in Experimental HTN
2.2. ET-1 in Human HTN
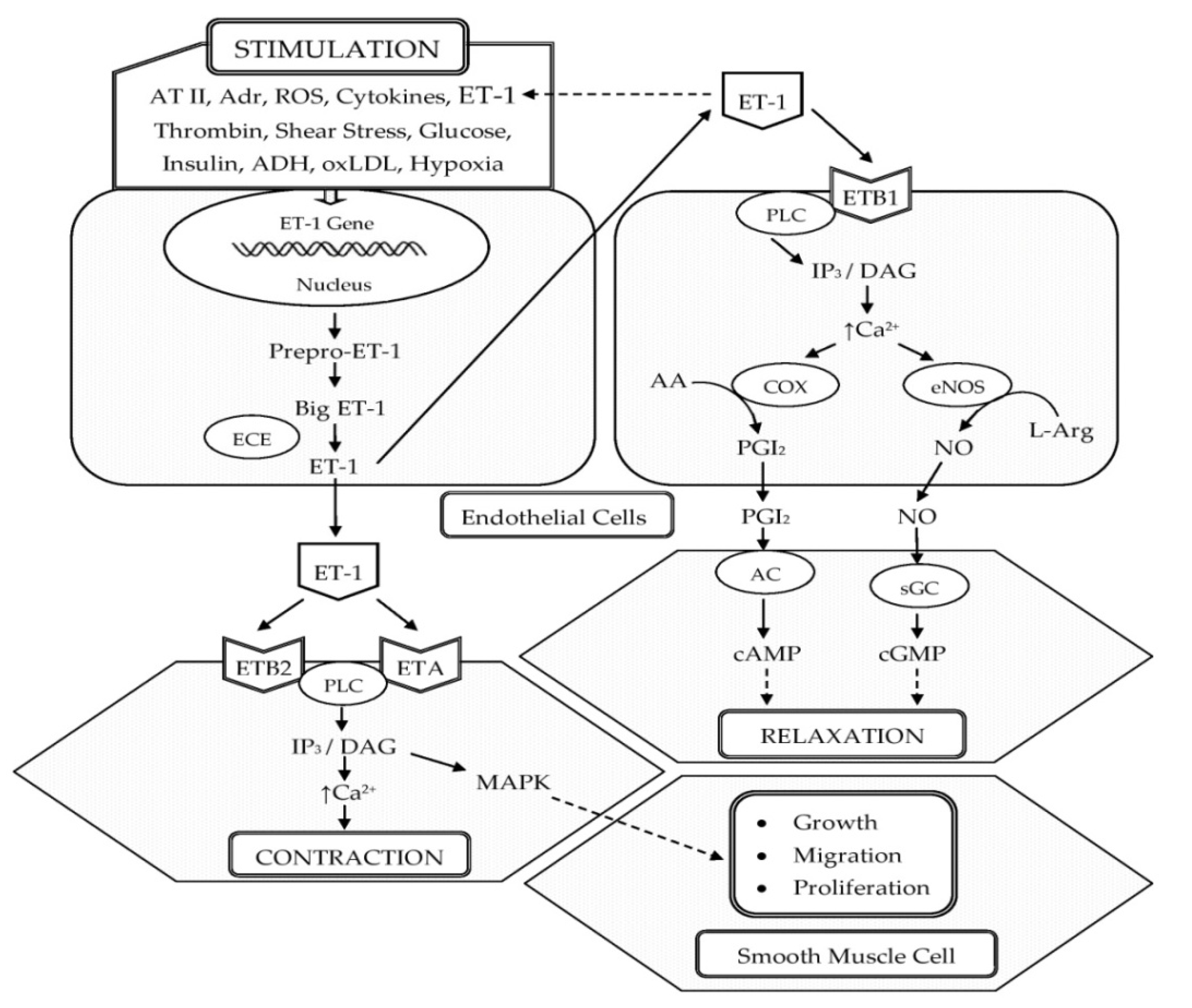
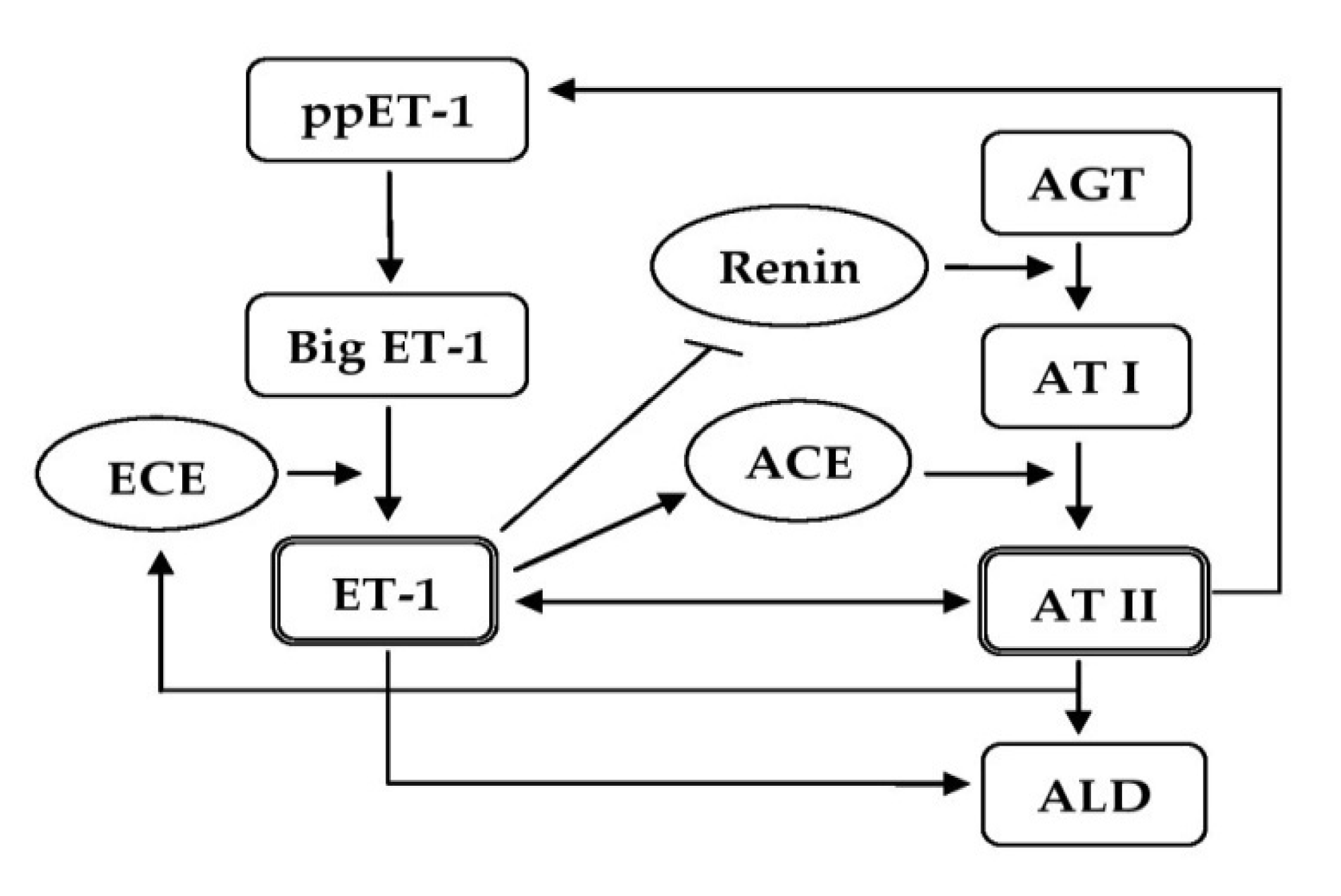
3. Role of ET-1 in the Pathogenesis of HTN
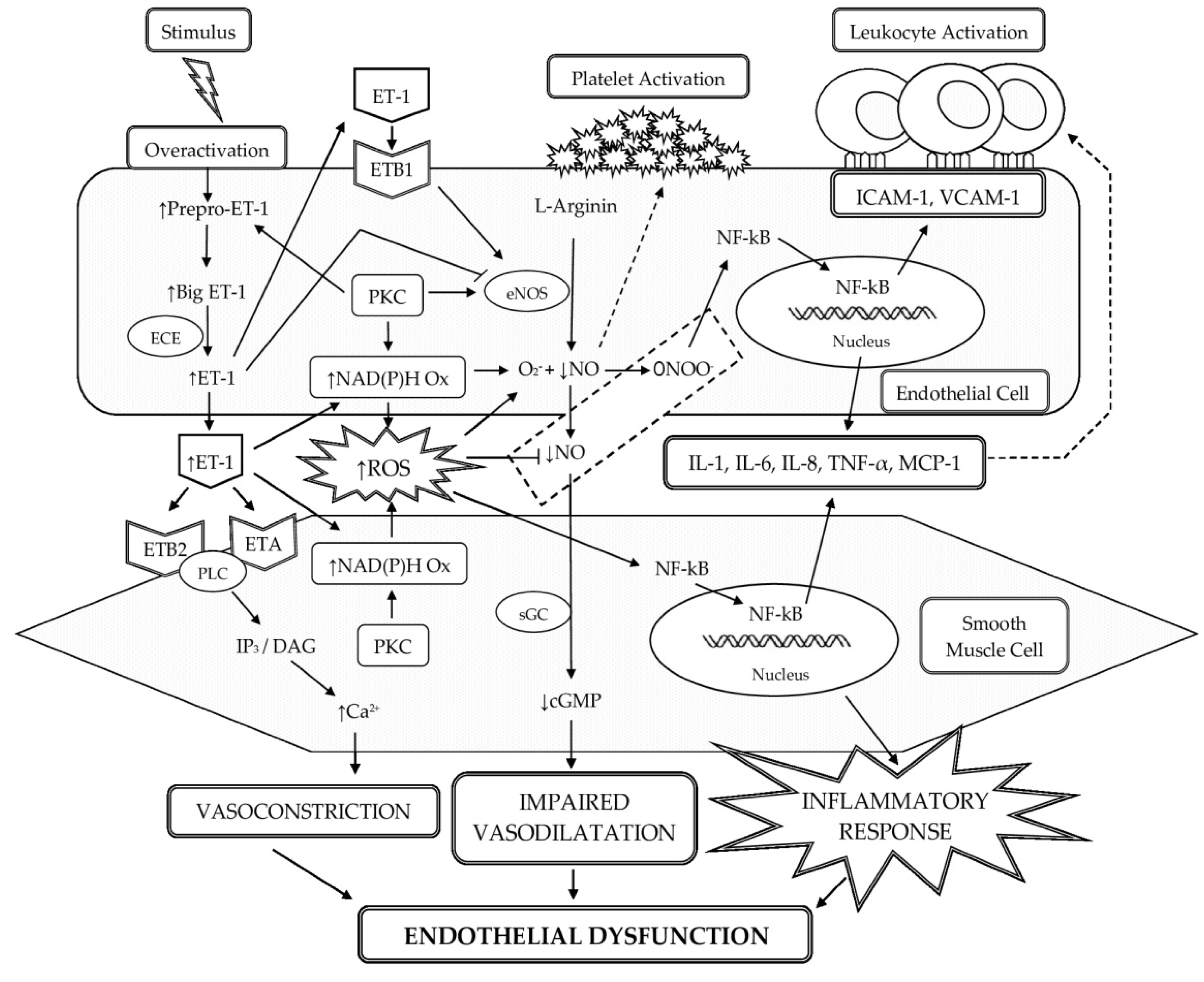
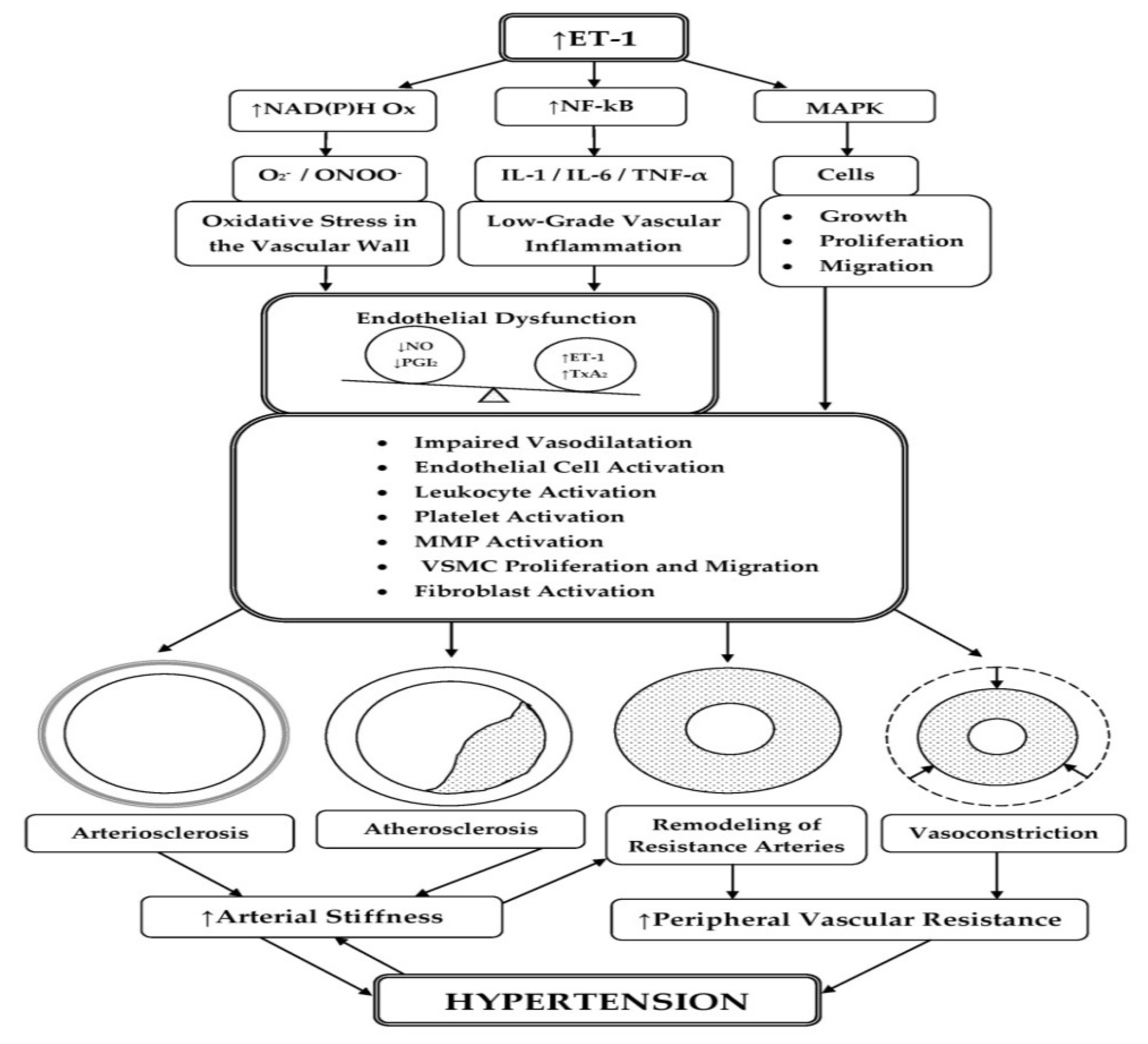
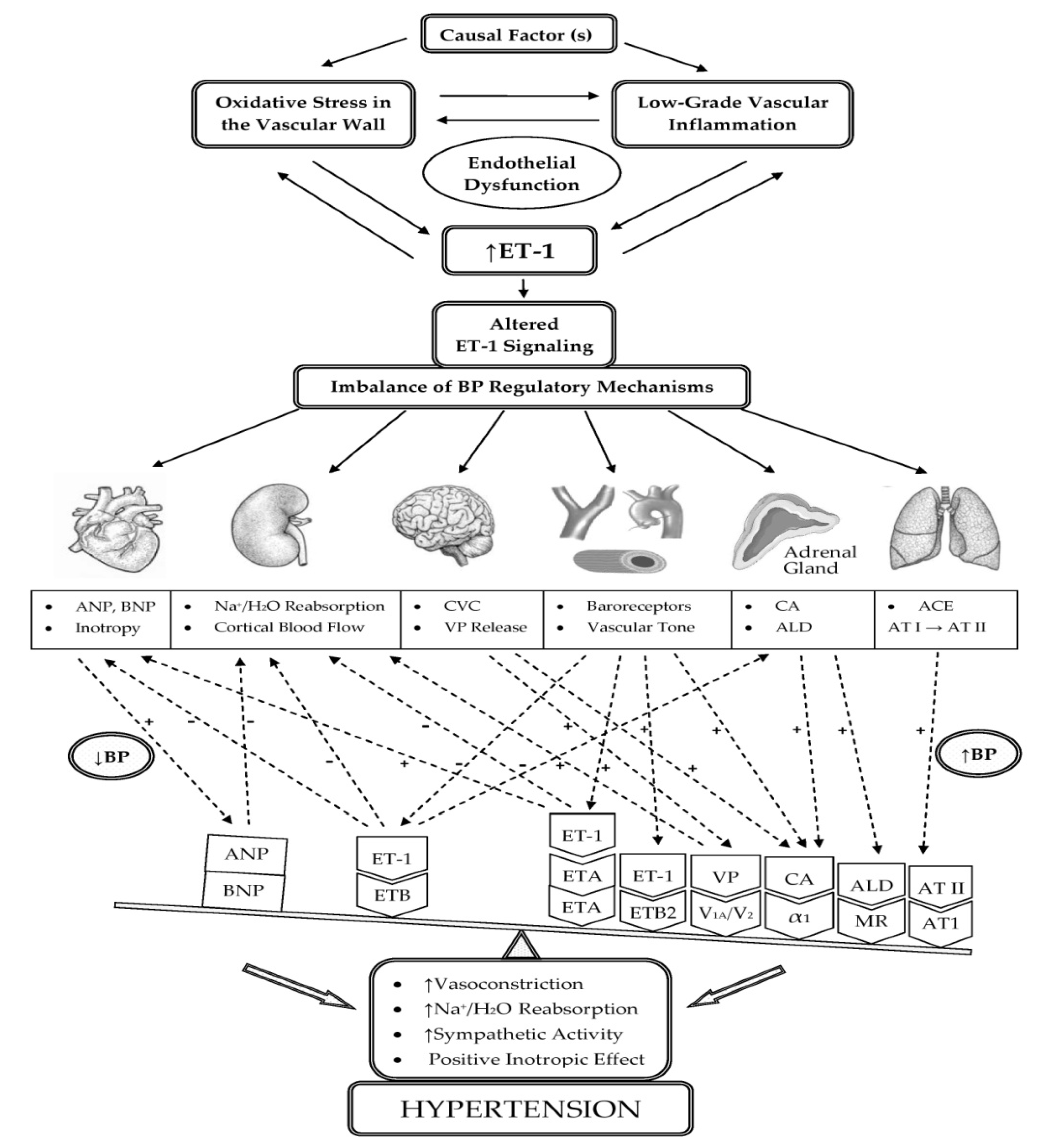
3.1. Participation of ET-1 in the Development of Oxidative Stress and Low-Grade Inflammation in the Vascular Wall with the Occurrence of ED
3.2. Participation of ET-1 in the Pathogenesis of Arterial Stiffness
3.2.1. Role of ET-1 in Arteriosclerosis
3.2.2. Role of ET-1 in Atherosclerosis
3.3. Participation of ET-1 in the Processes of Arterial Remodeling
3.4. Participation of ET-1 in the Mechanisms of BP Regulation
4. ET-1 as a Potential Therapeutic Target in HTN
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oparil, S.; Acelajado, M.C.; Bakris, G.L.; Berlowitz, D.R.; Cífková, R.; Dominiczak, A.F.; Grassi, G.; Jordan, J.; Poulter, N.R.; Rodgers, A.; et al. Hypertension. Nat. Rev. Dis. Primers 2018, 4, 18014. [Google Scholar] [CrossRef] [Green Version]
- Mensah, G.A.; Croft, J.B.; Giles, W.H. The heart, kidney, and brain as target organs in hypertension. Curr. Probl. Cardiol. 2003, 28, 156–193. [Google Scholar] [CrossRef]
- Mills, K.T.; Stefanescu, A.; He, J. The global epidemiology of hypertension. Nat. Rev. Nephrol. 2020, 16, 223–237. [Google Scholar] [CrossRef]
- Touyz, R.M.; Feldman, R.D.; Harrison, D.G.; Schiffrin, E.L. A new look at the mosaic theory of hypertension. Can. J. Cardiol. 2020, 36, 591–592. [Google Scholar] [CrossRef]
- Félétou, M.; Köhler, R.; Vanhoutte, P.M. Endothelium-derived vasoactive factors and hypertension: Possible roles in pathogenesis and as treatment targets. Curr. Hypertens. Rep. 2010, 12, 267–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahill, P.A.; Redmond, E.M. Vascular endothelium–gatekeeper of vessel health. Atherosclerosis 2016, 248, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, G.E.; Senger, D.R. Endothelial extracellular matrix: Biosynthesis, remodeling, and functions during vascular morphogenesis and neovessel stabilization. Circ. Res. 2005, 97, 1093–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoo, A.; van Zanten, J.J.V.; Metsios, G.S.; Carroll, D.; Kitas, G.D. The endothelium and its role in regulating vascular tone. Open Cardiovasc. Med. J. 2010, 4, 302–312. [Google Scholar] [CrossRef]
- Konukoglu, D.; Uzun, H. Endothelial dysfunction and hypertension. Hypertension 2016, 2, 511–540. [Google Scholar]
- Masaki, T.; Sawamura, T. Endothelin and endothelial dysfunction. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2006, 82, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Yanagisawa, M.; Kurihara, H.; Kimura, S.; Tomobe, Y.; Kobayashi, M.; Mitsui, Y.; Yazaki, Y.; Goto, K.; Masaki, T. A Novel potent vasoconstrictor peptide produced by vascular endothelial cells. Nature 1988, 332, 411–415. [Google Scholar] [CrossRef] [Green Version]
- Inoue, A.; Yanagisawa, M.; Kimura, S.; Kasuya, Y.; Miyauchi, T.; Goto, K.; Masaki, T. The human endothelin family: Three structurally and pharmacologically distinct isopeptides predicted by three separate genes. Proc. Natl. Acad. Sci. USA 1989, 86, 2863–2867. [Google Scholar] [CrossRef] [Green Version]
- Davenport, A.P.; Hyndman, K.A.; Dhaun, N.; Southan, C.; Kohan, D.E.; Pollock, J.S.; Pollock, D.M.; Webb, D.J.; Maguire, J.J. Endothelin. Pharmacol. Rev. 2016, 68, 357–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emoto, N. Endothelin receptor antagonist. In Diagnosis and Treatment of Pulmonary Hypertension; Springer: Berlin/Heidelberg, Germany, 2017; pp. 153–169. [Google Scholar]
- Pollock, D.M.; Keith, T.L.; Highsmith, R.F. Endothelin receptors and calcium signaling. FASEB J. 1995, 9, 1196–1204. [Google Scholar] [CrossRef] [Green Version]
- Miyagawa, K.; Emoto, N. Current state of endothelin receptor antagonism in hypertension and pulmonary hypertension. Ther. Adv. Cardiovasc. Dis. 2014, 8, 202–216. [Google Scholar] [CrossRef] [PubMed]
- Hynynen, M.M.; Khalil, R.A. The vascular endothelin system in hypertension–recent patents and discoveries. Recent Pat. Cardiovasc. Drug Discov. 2006, 1, 95–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maguire, J.J.; Davenport, A.P. Endothelin receptors and their antagonists. Semin. Nephrol. 2015, 35, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Schiffrin, E.L. Does endothelin-1 raise or lower blood pressure in humans? Nephron 2018, 139, 47–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiowski, W.; Lüscher, T.; Linder, L.; Bühler, F. Endothelin-1-induced vasoconstriction in humans. Reversal by calcium channel blockade but not by nitrovasodilators or endothelium-derived relaxing factor. Circulation 1991, 83, 469–475. [Google Scholar] [CrossRef] [Green Version]
- Taddei, S.; Virdis, A.; Ghiadoni, L.; Sudano, I.; Magagna, A.; Salvetti, A. Role of endothelin in the control of peripheral vascular tone in human hypertension. Heart Fail. Rev. 2001, 6, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Webb, D.J. Endogenous endothelin generation maintains vascular tone in humans. J. Hum. Hypertens. 1995, 9, 459–463. [Google Scholar] [PubMed]
- Haynes, W.G.; Webb, D.J. Contribution of endogenous generation of endothelin-1 to basal vascular tone. Lancet 1994, 344, 852–854. [Google Scholar] [CrossRef]
- Kohan, D.E.; Rossi, N.F.; Inscho, E.W.; Pollock, D.M. Regulation of blood pressure and salt homeostasis by endothelin. Physiol. Rev. 2011, 91, 1–77. [Google Scholar] [CrossRef] [PubMed]
- Speed, J.S.; Fox, B.M.; Johnston, J.G.; Pollock, D.M. Endothelin and renal ion and water transport. Semin. Nephrol. 2015, 35, 137–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhaun, N.; Goddard, J.; Kohan, D.E.; Pollock, D.M.; Schiffrin, E.L.; Webb, D.J. Role of endothelin-1 in clinical hypertension: 20 years on. Hypertension 2008, 52, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Amiri, F.; Virdis, A.; Neves, M.F.; Iglarz, M.; Seidah, N.G.; Touyz, R.M.; Reudelhuber, T.L.; Schiffrin, E.L. Endothelium-restricted overexpression of human endothelin-1 causes vascular remodeling and endothelial dysfunction. Circulation 2004, 110, 2233–2240. [Google Scholar] [CrossRef] [Green Version]
- Leung, J.W.-C.; Wong, W.T.; Koon, H.W.; Mo, F.M.; Tam, S.; Huang, Y.; Vanhoutte, P.M.; Chung, S.S.M.; Chung, S.K. Transgenic mice over-expressing ET-1 in the endothelial cells develop systemic hypertension with altered vascular reactivity. PLoS ONE 2011, 6, e26994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, D.; Ge, Y.; Stricklett, P.K.; Gill, P.; Taylor, D.; Hughes, A.K.; Yanagisawa, M.; Miller, L.; Nelson, R.D.; Kohan, D.E. Collecting duct–specific knockout of endothelin-1 causes hypertension and sodium retention. J. Clin. Investig. 2004, 114, 504–511. [Google Scholar] [CrossRef] [Green Version]
- Ge, Y.; Bagnall, A.; Stricklett, P.K.; Strait, K.; Webb, D.J.; Kotelevtsev, Y.; Kohan, D.E. Collecting duct-specific knockout of the endothelin B receptor causes hypertension and sodium retention. Am. J. Physiol. Renal Physiol. 2006, 291, F1274–F1280. [Google Scholar] [CrossRef] [Green Version]
- Ge, Y.; Bagnall, A.; Stricklett, P.K.; Webb, D.; Kotelevtsev, Y.; Kohan, D.E. Combined knockout of collecting duct endothelin A and B receptors causes hypertension and sodium retention. Am. J. Physiol. Renal Physiol. 2008, 295, F1635–F1640. [Google Scholar] [CrossRef] [Green Version]
- Schiffrin, E.L. Endothelin: Role in experimental hypertension. J. Cardiovasc. Pharmacol. 2000, 35, S33–S35. [Google Scholar] [CrossRef]
- Schiffrin, E.L.; Turgeon, A.; Deng, L.Y. Effect of chronic ETA-selective endothelin receptor antagonism on blood pressure in experimental and genetic hypertension in rats. Br. J. Pharmacol. 1997, 121, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Oishi, S.; Sasaki, M.; Sato, T. Elevated immunoreactive endothelin levels in patients with pheochromocytoma. Am. J. Hypertens. 1994, 7, 717–722. [Google Scholar] [CrossRef]
- Yokokawa, K.; Tahara, H.; Kohno, M.; Murakawa, K.I.; Yasunari, K.; Nakagawa, K.; Hamada, T.; Otani, S.; Yanagisawa, M.; Takeda, T. Hypertension associated with endothelin-secreting malignant hemangioendothelioma. Ann. Intern. Med. 1991, 114, 213–215. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.; Yanagisawa, M. Endothelin: 20 years from discovery to therapy. Can. J. Physiol. Pharmacol. 2008, 86, 485–498. [Google Scholar] [CrossRef]
- Kumagae, S.I.; Adachi, H.; Jacobs, D.R., Jr.; Hirai, Y.; Enomoto, M.; Fukami, A.; Otsuka, M.; Nanjo, Y.; Esaki, E.; Kumagai, E.; et al. High level of plasma endothelin-1 predicts development of hypertension in normotensive subjects. Am. J. Hypertens. 2010, 23, 1103–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krum, H.; Viskoper, R.J.; Lacourciere, Y.; Budde, M.; Charlon, V. the effect of an endothelin-receptor antagonist, bosentan, on blood pressure in patients with essential hypertension. N. Engl. J. Med. 1998, 338, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Roumen, N.; Egon, P.; Siegfried, E. Darusentan: An effective endothelina receptor antagonist for treatment of hypertension. Am. J. Hypertens. 2002, 15, 583–589. [Google Scholar]
- Black, H.R.; Bakris, G.L.; Weber, M.A.; Weiss, R.; Shahawy, M.E.; Marple, R.; Tannoury, G.; Linas, S.; Wiens, B.L.; Linseman, J.V. Efficacy and safety of darusentan in patients with resistant hypertension: Results from a randomized, double-blind, placebo-controlled dose-ranging study. J. Clin. Hypertens. 2007, 9, 760–769. [Google Scholar] [CrossRef] [Green Version]
- Bakris, G.L.; Lindholm, L.H.; Black, H.R.; Krum, H.; Linas, S.; Linseman, J.V.; Arterburn, S.; Sager, P.; Weber, M. Divergent results using clinic and ambulatory blood pressures: Report of a darusentan-resistant hypertension trial. Hypertension 2010, 56, 824–830. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Lu, Y.P.; Hasan, A.A.; Hocher, B. Plasma ET-1 concentrations are elevated in patients with hypertension–meta-analysis of clinical studies. Kidney Blood Press. Res. 2017, 42, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.; Shaw, S.; d’Uscio, L.V.; Moreau, P.; Lüscher, T.F. Angiotensin II increases vascular and renal endothelin-1 and functional endothelin converting enzyme activity in vivo: Role of ETA receptors for endothelin regulation. Biochem. Biophys. Res. Commun. 1997, 238, 861–865. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Sietsma, S.J.; Paul, M. A role for endothelin in the pathogenesis of hypertension: Fact or fiction? Kidney Int. 1998, 54, S115–S121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Omari, M.A.; Khaleghi, M.; Mosley, T.H.; Morgenthaler, N.G.; Struck, J.; Bergmann, A.; Kullo, I.J. Plasma C-terminal pro-endothelin-1 is associated with left ventricular mass index and aortic root diameter in African-American adults with hypertension. J. Hum. Hypertens. 2011, 25, 106–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolpe, A.G.; Ruddiman, C.A.; Hall, P.J.; Isakson, B.E. Polarized proteins in endothelium and their contribution to function. J.Vasc. Res. 2021, 58, 65–91. [Google Scholar]
- Saito, Y.; Nakao, K.; Mukoyama, M.; Imura, H. Increased plasma endothelin level in patients with essential hypertension. N. Engl. J. Med. 1990, 322, 205. [Google Scholar]
- Shichiri, M.; Hirata, Y.; Ando, K.; Emori, T.; Ohta, K.; Kimoto, S.; Ogura, M.; Inoue, A.; Marumo, F. Plasma endothelin levels in hypertension and chronic renal failure. Hypertension 1990, 15, 493–496. [Google Scholar] [CrossRef] [Green Version]
- Parrinello, G.; Scaglione, R.; Pinto, A.; Corrao, S.; Cecala, M.; Di Silvestre, G.; Amato, P.; Licata, A.; Licata, G. Central obesity and hypertension: The role of plasma endothelin. Am. J. Hypertens. 1996, 9, 1186–1191. [Google Scholar] [CrossRef] [Green Version]
- Amoroso, A.; Cossu, M.; Mariotti, A.; Guido, F.; Ferri, G.; De Rosa, F.; Sportelli, G. Increased plasma levels of endothelin in patients with essential arterial hypertension. Eur. Rev. Med. Pharmacol. Sci. 1996, 18, 33–37. [Google Scholar]
- Schneider, M.P.; Hilgers, K.F.; Klingbeil, A.U.; John, S.; Veelken, R.; Schmieder, R.E. Plasma endothelin is increased in early essential hypertension. Am. J. Hypertens. 2000, 13, 579–585. [Google Scholar] [CrossRef] [Green Version]
- Parissis, J.T.; Venetsanou, K.F.; Mentzikof, D.G.; Kalantzi, M.V.; Georgopoulou, M.V.; Chrisopoulos, N.; Karas, S.M. Plasma levels of soluble cellular adhesion molecules in patients with arterial hypertension. Correlations with plasma endothelin-1. Eur. J. Intern. Med. 2001, 12, 350–356. [Google Scholar] [CrossRef]
- Kostov, K.; Dimitrova, A.; Grigoryan, A.; Tisheva, S.; Ruseva, A.; Atanasova, M.; Gospodinov, C.; Blazhev, A. Changes in the serum levels of endothelin-1, matrix metalloproteinases-2,-9 and C-reactive protein in patients with mild and severe degree of arterial hypertension. C. R. Acad. Bulg. Sci. 2014, 67, 427–434. [Google Scholar]
- Gu, X.; Li, H.; Zhu, X.; Gu, H.; Chen, J.; Wang, L.; Harding, P.; Xu, W. Inverse correlation between plasma adropin and ET-1 levels in essential hypertension: A cross-sectional study. Medicine 2015, 94, e1712. [Google Scholar] [CrossRef] [PubMed]
- Kostov, K.; Blazhev, A.; Atanasova, M.; Dimitrova, A. Serum concentrations of endothelin-1 and matrix metalloproteinases-2, -9 in pre-hypertensive and hypertensive patients with type 2 diabetes. Int. J. Mol. Sci. 2016, 17, 1182. [Google Scholar] [CrossRef] [PubMed]
- del Villar, C.P.; Alonso, C.J.G.; Feldstein, C.A.; Juncos, L.A.; Romero, J.C. Role of endothelin in the pathogenesis of hypertension. In Mayo Clinic Proceedings; Elsevier: Amsterdam, The Netherlands, 2005; Volume 80, pp. 84–96. [Google Scholar]
- Iglarz, M.; Schiffrin, E.L. Role of endothelin-1 in hypertension. Curr. Hypertens. Rep. 2003, 5, 144–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.A.; Jesmin, S.; Islam, M.M.; Sohael, F.; Hasan, A.S.H.; Zaedi, S.; Sultana, S.N.; Yamaguchi, N.; Kawano, S.; Okazaki, O. Circulatory level of endothelin-1 and hypertension in rural women in Bangladesh: A potential association evidenced from a community based cross-sectional study. J. Hypertens. 2015, 33, e17–e18. [Google Scholar] [CrossRef]
- Skalska, A.B.; Pietrzycka, A.; Stępniewski, M. Correlation of endothelin-1 plasma levels with plasma antioxidant capacity in elderly patients treated for hypertension. Clin. Biochem. 2009, 42, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.; Jiménez, R.; Sánchez, M.; López-Sepúlveda, R.; Zarzuelo, A.; Tamargo, J.; Pérez-Vizcaíno, F.; Duarte, J. Vascular superoxide production by endothelin-1 requires Src non-receptor protein tyrosine kinase and MAPK activation. Atherosclerosis 2010, 212, 78–85. [Google Scholar] [CrossRef]
- Piechota, A.; Polańczyk, A.; Gorąca, A. Role of endothelin-1 receptor blockers on hemodynamic parameters and oxidative stress. Pharmacol. Rep. 2010, 62, 28–34. [Google Scholar] [CrossRef]
- Touyz, R.M.; Rios, F.J.; Alves-Lopes, R.; Neves, K.B.; Camargo, L.L.; Montezano, A.C. Oxidative stress: A unifying paradigm in hypertension. Can. J. Cardiol. 2020, 36, 659–670. [Google Scholar] [CrossRef] [Green Version]
- Böhm, F.; Pernow, J. The importance of endothelin-1 for vascular dysfunction in cardiovascular disease. Cardiovasc. Res. 2007, 76, 8–18. [Google Scholar] [CrossRef] [Green Version]
- Pernow, J.; Shemyakin, A.; Böhm, F. New perspectives on endothelin-1 in atherosclerosis and diabetes mellitus. Life Sci. 2012, 91, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Tomiyama, H.; Shiina, K.; Matsumoto-Nakano, C.; Ninomiya, T.; Komatsu, S.; Kimura, K.; Chikamori, T.; Yamashina, A. The contribution of inflammation to the development of hypertension mediated by increased arterial stiffness. J. Am. Heart Assoc. 2017, 6, e005729. [Google Scholar] [CrossRef]
- Vaziri, N.D. Roles of oxidative stress and antioxidant therapy in chronic kidney disease and hypertension. Curr. Opin. Nephrol. Hypertens. 2004, 13, 93–99. [Google Scholar] [CrossRef]
- Peterson, J.R.; Sharma, R.V.; Davisson, R.L. Reactive oxygen species in the neuropathogenesis of hypertension. Curr. Hypertens. Rep. 2006, 8, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Harrison, D.G.; Gongora, M.C. Oxidative stress and hypertension. Med. Clin. N. Am. 2009, 93, 621–635. [Google Scholar] [CrossRef]
- Briones, A.M.; Touyz, R.M. Oxidative stress and hypertension: Current concepts. Curr. Hypertens. Rep. 2010, 12, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Zhang, X.; Wold, L.E.; Ren, Q.; Zhang, Z.; Ren, J. Endothelin-1 enhances oxidative stress, cell proliferation and reduces apoptosis in human umbilical vein endothelial cells: Role of ETB receptor, NADPH oxidase and caveolin-1. Br. J. Pharmacol. 2005, 145, 323–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duerrschmidt, N.; Wippich, N.; Goettsch, W.; Broemme, H.J.; Morawietz, H. Endothelin-1 induces NAD(P)H oxidase in human endothelial cells. Biochem. Biophys. Res. Commun. 2000, 269, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Galle, J.; Lehmann-Bodem, C.; Hübner, U.; Heinloth, A.; Wanner, C. CyA and OxLDL cause endothelial dysfunction in isolated arteries through endothelin-mediated stimulation of O2− formation. Nephrol. Dial. Transplant. 2000, 15, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Loomis, E.D.; Sullivan, J.C.; Osmond, D.A.; Pollock, D.M.; Pollock, J.S. Endothelin mediates superoxide production and vasoconstriction through activation of NADPH oxidase and uncoupled nitric-oxide synthase in the rat aorta. J. Pharmacol. Exp. Ther. 2005, 315, 1058–1064. [Google Scholar] [CrossRef]
- López-Sepúlveda, R.; Gómez-Guzmán, M.; Zarzuelo, M.J.; Romero, M.; Sánchez, M.; Quintela, A.M.; Galindo, P.; O’Valle, F.; Tamargo, J.; Pérez-Vizcaíno, F. Red wine polyphenols prevent endothelial dysfunction induced by endothelin-1 in rat aorta: Role of NADPH oxidase. Clin. Sci. 2011, 120, 321–333. [Google Scholar] [CrossRef] [Green Version]
- Mohazzab, K.; Kaminski, P.M.; Wolin, M.S. NADH oxidoreductase is a major source of superoxide anion in bovine coronary artery endothelium. Am. J. Physiol. Heart Circ. Physiol. 1994, 266, H2568–H2572. [Google Scholar] [CrossRef] [PubMed]
- Kamata, K.; Kanie, N.; Matsumoto, T.; Kobayashi, T. Endothelin-1-induced impairment of endothelium-dependent relaxation in aortas isolated from controls and diabetic rats. J. Cardiovasc. Pharmacol. 2004, 44, S186–S190. [Google Scholar] [CrossRef] [PubMed]
- Kanie, N.; Kamata, K. Effects of chronic administration of the novel endothelin antagonist J-104132 on endothelial dysfunction in streptozotocin-induced diabetic rat. Br. J. Pharmacol. 2002, 135, 1935–1942. [Google Scholar] [CrossRef] [Green Version]
- Camici, G.G.; Sudano, I.; Noll, G.; Tanner, F.C.; Lüscher, T.F. Molecular pathways of aging and hypertension. Curr. Opin. Nephrol. Hypertens. 2009, 18, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Savoia, C.; Sada, L.; Zezza, L.; Pucci, L.; Lauri, F.M.; Befani, A.; Alonzo, A.; Volpe, M. Vascular inflammation and endothelial dysfunction in experimental hypertension. Int. J. Hypertens. 2011, 2011, 281240. [Google Scholar] [CrossRef] [PubMed]
- Barrows, I.R.; Ramezani, A.; Raj, D.S. Inflammation, immunity, and oxidative stress in—partners in crime? Adv. Chronic Kidney Dis. 2019, 26, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Guzik, T.J.; Touyz, R.M. Oxidative stress, inflammation, and vascular aging in hypertension. Hypertension 2017, 70, 660–667. [Google Scholar] [CrossRef]
- Kähler, J.; Mendel, S.; Weckmüller, J.; Orzechowski, H.D.; Mittmann, C.; Köster, R.; Paul, M.; Meinertz, T.; Münzel, T. Oxidative stress increases synthesis of big endothelin-1 by activation of the endothelin-1 promoter. J. Mol. Cell. Cardiol. 2000, 32, 1429–1437. [Google Scholar] [CrossRef]
- Anggrahini, D.W.; Emoto, N.; Nakayama, K.; Widyantoro, B.; Adiarto, S.; Iwasa, N.; Nonaka, H.; Rikitake, Y.; Kisanuki, Y.Y.; Yanagisawa, M.; et al. Vascular endothelial cell-derived endothelin-1 mediates vascular inflammation and neointima formation following blood flow cessation. Cardiovasc. Res. 2009, 82, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruetten, H.; Thiemermann, C. Endothelin-1 stimulates the biosynthesis of tumour necrosis factor in macrophages: ET-receptors, signal transduction and inhibition by dexamethasone. J. Physiol. Pharmacol. 1997, 48, 675–688. [Google Scholar] [PubMed]
- Hofman, F.M.; Chen, P.; Jeyaseelan, R.; Incardona, F.; Fisher, M.; Zidovetzki, R. Endothelin-1 induces production of the neutrophil chemotactic factor interleukin-8 by human brain-derived endothelial cells. Blood 1998, 92, 3064–3072. [Google Scholar] [CrossRef] [PubMed]
- Browatzki, M.; Schmidt, J.; Kübler, W.; Kranzhöfer, R. Endothelin-1 induces interleukin-6 release via acctivation of the transcription factor NF-κB in human vascular smooth muscle cells. Basic Res. Cardiol. 2000, 95, 98–105. [Google Scholar] [CrossRef]
- Yang, L.L.; Gros, R.; Kabir, M.G.; Sadi, A.; Gotlieb, A.I.; Husain, M.; Stewart, D.J. Conditional cardiac overexpression of endothelin-1 induces inflammation and dilated cardiomyopathy in mice. Circulation 2004, 109, 255–261. [Google Scholar] [CrossRef] [Green Version]
- Virdis, A.; Schiffrin, E.L. Vascular Inflammation: A role in vascular disease in hypertension? Curr. Opin. Nephrol. Hypertens. 2003, 12, 181–187. [Google Scholar] [CrossRef]
- Vierhapper, H.; Wagner, O.; Nowotny, P.; Waldhäusl, W. Effect of endothelin-1 in man. Circulation 1990, 81, 1415–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letizia, C.; Celi, M.; Cerci, S.; Scuro, L.; Delfini, E.; Subioli, S.; Caliumi, C.; D’Erasmo, E. High circulating levels of adrenomedullin and endothelin-1 in obesity associated with arterial hypertension. Ital. Heart J. 2001, 2, 1011–1015. [Google Scholar]
- McEniery, C.M.; Qasem, A.; Schmitt, M.; Avolio, A.P.; Cockcroft, J.R.; Wilkinson, I.B. Endothelin-1 regulates arterial pulse wave velocity in vivo. J. Am. Coll. Cardiol. 2003, 42, 1975–1981. [Google Scholar] [CrossRef] [Green Version]
- Vuurmans, T.J.; Boer, P.; Koomans, H.A. Effects of endothelin-1 and endothelin-1 receptor blockade on cardiac output, aortic pressure, and pulse wave velocity in humans. Hypertension 2003, 41, 1253–1258. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.Y.; Oh, B.H. Aging and arterial stiffness. Circ. J. 2010, 74, 2257–2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z. Aging, arterial stiffness, and hypertension. Hypertension 2015, 65, 252–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Rourke, M.F. Arterial aging: Pathophysiological principles. Vasc. Med. 2007, 12, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Palombo, C.; Kozakova, M. Arterial stiffness, atherosclerosis and cardiovascular risk: Pathophysiologic mechanisms and emerging clinical indications. Vascul. Pharmacol. 2016, 77, 1–7. [Google Scholar] [CrossRef]
- Kim, H.L.; Kim, S.H. Pulse wave velocity in atherosclerosis. Front. Cardiovasc. Med. 2019, 6, 41. [Google Scholar] [CrossRef]
- Van Popele, N.M.; Grobbee, D.E.; Bots, M.L.; Asmar, R.; Topouchian, J.; Reneman, R.S.; Hoeks, A.P.; van der Kuip, D.A.; Hofman, A.; Witteman, J.C. Association between arterial stiffness and atherosclerosis: The Rotterdam Study. Stroke 2001, 32, 454–460. [Google Scholar] [CrossRef] [Green Version]
- Rogers, W.J.; Hu, Y.L.; Coast, D.; Vido, D.A.; Kramer, C.M.; Pyeritz, R.E.; Reichek, N. Age-associated changes in regional aortic pulse wave velocity. J. Am. Coll. Cardiol. 2001, 38, 1123–1129. [Google Scholar] [CrossRef] [Green Version]
- Sutton-Tyrrell, K.; Najjar, S.S.; Boudreau, R.M.; Venkitachalam, L.; Kupelian, V.; Simonsick, E.M.; Havlik, R.; Lakatta, E.G.; Spurgeon, H.; Kritchevsky, S.; et al. Elevated aortic pulse wave velocity, a marker of arterial stiffness, predicts cardiovascular events in well-functioning older adults. Circulation 2005, 111, 3384–3390. [Google Scholar] [CrossRef]
- Laurent, S.; Boutouyrie, P. Arterial stiffness and hypertension in the elderly. Front. Cardiovasc. Med. 2020, 7, 202. [Google Scholar] [CrossRef]
- Wallace, S.M.L.; Yasmin; McEniery, C.M.; Mäki-Petäjä, K.M.; Booth, A.D.; Cockcroft, J.R.; Wilkinson, I.B. Isolated systolic hypertension is characterized by increased aortic stiffness and endothelial dysfunction. Hypertension 2007, 50, 228–233. [Google Scholar] [CrossRef] [Green Version]
- Sawabe, M. Vascular aging: From molecular mechanism to clinical significance. Geriatr. Gerontol. Int. 2010, 10, S213–S220. [Google Scholar] [CrossRef]
- Laurent, S.; Boutouyrie, P.; Lacolley, P. Structural and genetic bases of arterial stiffness. Hypertension 2005, 45, 1050–1055. [Google Scholar] [CrossRef]
- Harvey, A.; Montezano, A.C.; Lopes, R.A.; Rios, F.; Touyz, R.M. Vascular fibrosis in aging and hypertension: Molecular mechanisms and clinical implications. Can. J. Cardiol. 2016, 32, 659–668. [Google Scholar] [CrossRef] [Green Version]
- Trindade, M.; Oigman, W.; Fritsch Neves, M. Potential role of endothelin in early vascular aging. Curr. Hypertens. Rev. 2017, 13, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Guilder, G.P.V.; Westby, C.M.; Greiner, J.J.; Stauffer, B.L.; DeSouza, C.A. Endothelin-1 vasoconstrictor tone increases with age in healthy men but can be reduced by regular aerobic exercise. Hypertension 2007, 50, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Thijssen, D.H.J.; Rongen, G.A.; Van Dijk, A.; Smits, P.; Hopman, M.T.E. Enhanced endothelin-1-mediated leg vascular tone in healthy older subjects. J. Appl. Physiol. 2007, 103, 852–857. [Google Scholar] [CrossRef] [Green Version]
- Tokunaga, O.; Fan, J.; Watanabe, T.; Kobayashi, M.; Kumazaki, T.; Mitsui, Y. Endothelin. Immunohistologic localization in aorta and biosynthesis by cultured human aortic endothelial cells. Lab. Investig. 1992, 67, 210–217. [Google Scholar]
- Seals, D.R.; Jablonski, K.L.; Donato, A.J. Aging and vascular endothelial function in humans. Clin. Sci. 2011, 120, 357–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horstmeyer, A.; Licht, C.; Scherr, G.; Eckes, B.; Krieg, T. Signalling and regulation of collagen I synthesis by ET-1 and TGF-β1. FEBS J. 2005, 272, 6297–6309. [Google Scholar] [CrossRef] [PubMed]
- Hafizi, S.; Wharton, J.; Chester, A.H.; Yacoub, M.H. Profibrotic effects of endothelin-1 via the ETA receptor in cultured human cardiac fibroblasts. Cell. Physiol. Biochem. 2004, 14, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Clozel, M.; Salloukh, H. Role of endothelin in fibrosis and anti-fibrotic potential of bosentan. Ann. Med. 2005, 37, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Wermuth, P.J.; Li, Z.; Mendoza, F.A.; Jimenez, S.A. Stimulation of transforming growth factor-β1-induced endothelial-to-mesenchymal transition and tissue fibrosis by endothelin-1 (ET-1): A novel profibrotic effect of ET-1. PLoS ONE 2016, 11, e0161988. [Google Scholar]
- Barton, M.; Haudenschild, C.C.; d’Uscio, L.V.; Shaw, S.; Münter, K.; Lüscher, T.F. Endothelin ETA receptor blockade restores NO-mediated endothelial function and inhibits atherosclerosis in apolipoprotein E-deficient mice. Proc. Natl. Acad. Sci. USA 1998, 95, 14367–14372. [Google Scholar] [CrossRef] [Green Version]
- Lerman, A.; Webster, M.W.; Chesebro, J.H.; Edwards, W.D.; Wei, C.M.; Fuster, V.; Burnett, J.C., Jr. Circulating and tissue endothelin immunoreactivity in hypercholesterolemic pigs. Circulation 1993, 88, 2923–2928. [Google Scholar] [CrossRef] [Green Version]
- Lerman, A.; Edwards, B.S.; Hallett, J.W.; Heublein, D.M.; Sandberg, S.M.; Burnett, J.C., Jr. Circulating and tissue endothelin immunoreactivity in advanced atherosclerosis. N. Engl. J. Med. 1991, 325, 997–1001. [Google Scholar] [CrossRef] [Green Version]
- Zeiher, A.M.; Ihling, C.; Pistorius, K.; Schächinger, V.; Schaefer, H.E. Increased tissue endothelin immunoreactivity in atherosclerotic lesions associated with acute coronary syndromes. Lancet 1994, 344, 1405–1406. [Google Scholar] [CrossRef]
- Böhm, F.; Johansson, B.L.; Hedin, U.; Alving, K.; Pernow, J. Enhanced vasoconstrictor effect of big endothelin-1 in patients with atherosclerosis: Relation to conversion to endothelin-1. Atherosclerosis 2002, 160, 215–222. [Google Scholar] [CrossRef]
- Li, M.W.; Mian, M.O.R.; Barhoumi, T.; Rehman, A.; Mann, K.; Paradis, P.; Schiffrin, E.L. Endothelin-1 overexpression exacerbates atherosclerosis and induces aortic aneurysms in apolipoprotein E knockout mice. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2306–2315. [Google Scholar] [CrossRef] [Green Version]
- Lerman, A.; Holmes, D.R., Jr.; Bell, M.R.; Garratt, K.N.; Nishimura, R.A.; Burnett, J.C., Jr. Endothelin in coronary endothelial dysfunction and early atherosclerosis in humans. Circulation 1995, 92, 2426–2431. [Google Scholar] [CrossRef] [PubMed]
- Ihling, C.; Szombathy, T.; Bohrmann, B.; Brockhaus, M.; Schaefer, H.E.; Loeffler, B.M. Coexpression of endothelin-converting enzyme-1 and endothelin-1 in different stages of human atherosclerosis. Circulation 2001, 104, 864–869. [Google Scholar] [CrossRef] [Green Version]
- Maguire, J.J.; Davenport, A.P. Increased response to big endothelin-1 in atherosclerotic human coronary artery: Functional evidence for up-regulation of endothelin-converting enzyme activity in disease. Br. J. Pharmacol. 1998, 125, 238–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sica, D.A. Endothelin receptor antagonism: What does the future hold? Hypertension 2008, 52, 460–461. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Su, X.; Qin, Q.; Yu, Y.; Jia, M.; Zhang, H.; Li, H.; Pei, L. New insights into phenotypic switching of VSMCs induced by hyperhomocysteinemia: Role of endothelin-1 signaling. Biomed. Pharmacother. 2020, 123, 109758. [Google Scholar] [CrossRef]
- Lyle, A.N.; Taylor, W.R. The pathophysiological basis of vascular disease. Lab. Investig. 2019, 99, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Intengan, H.D.; Schiffrin, E.L. Structure and mechanical properties of resistance arteries in hypertension. Hypertension 2000, 36, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Schiffrin, E.L. Role of endothelin-1 in hypertension and vascular disease. Am. J. Hypertens. 2001, 14, 83S–89S. [Google Scholar] [CrossRef] [Green Version]
- Schiffrin, E.L. Role of endothelin-1 in hypertension. Hypertension 1999, 34, 876–881. [Google Scholar] [CrossRef] [Green Version]
- Li, J.S.; Larivière, R.; Schiffrin, E.L. Effect of a nonselective endothelin antagonist on vascular remodeling in deoxycorticosterone acetate-salt hypertensive rats. Evidence for a role of endothelin in vascular hypertrophy. Hypertension 1994, 24, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Sayed, S.; Nussberger, J.; Aubert, J.F.; Gohlke, P.; Brunner, H.R.; Brakch, N. Measurement of plasma endothelin-1 in experimental hypertension and in healthy subjects. Am. J. Hypertens. 2003, 16, 515–521. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, T.; Ohta, H.; Okada, M.; Kawai, N.; Nakao, R.; Siegl, P.K.S.; Kobayashi, T.; Maeda, S.; Miyauchi, T.; Nishikibe, M. Pathophysiological roles of endothelin-1 in Dahl salt-sensitive hypertension. Hypertension 1999, 34, 514–519. [Google Scholar] [CrossRef] [Green Version]
- Schiffrin, E.L. Vascular remodeling in hypertension. Hypertension 2012, 59, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Nishida, Y.; Tandai-Hiruma, M.; Kemuriyama, T.; Hagisawa, K. Long-term blood pressure control: Is there a set-point in the brain? J. Physiol. Sci. 2012, 62, 147–161. [Google Scholar] [CrossRef]
- Speed, J.S.; Heimlich, J.B.; Hyndman, K.A.; Fox, B.M.; Patel, V.; Yanagisawa, M.; Pollock, J.S.; Titze, J.M.; Pollock, D.M. Endothelin Endothelin-1 as a master regulator of whole-body Na+ homeostasis. FASEB J. 2015, 29, 4937–4944. [Google Scholar] [CrossRef] [Green Version]
- Nohria, A.; Garrett, L.; Johnson, W.; Kinlay, S.; Ganz, P.; Creager, M.A. Endothelin-1 and vascular tone in subjects with atherogenic risk factors. Hypertension 2003, 42, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Haynes, W.G.; Ferro, C.E.; Webb, D.J. Physiologic role of endothelin in maintenance of vascular tone in humans. J. Cardiovasc. Pharmacol. 1995, 26, S183–S185. [Google Scholar] [CrossRef]
- Drawnel, F.M.; Archer, C.R.; Roderick, H.L. The role of the paracrine/autocrine mediator endothelin-1 in regulation of cardiac contractility and growth. Br. J. Pharmacol. 2013, 168, 296–317. [Google Scholar] [CrossRef] [Green Version]
- Zolk, O.; Münzel, F.; Eschenhagen, T. Effects of chronic endothelin-1 stimulation on cardiac myocyte contractile function. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H1248–H1257. [Google Scholar] [CrossRef] [PubMed]
- Piuhola, J.; Mäkinen, M.; Szokodi, I.; Ruskoaho, H. Dual role of endothelin-1 via ETA and ETB receptors in regulation of cardiac contractile function in mice. Am. J. Physiol. Heart Circ. Physiol. 2003, 285, H112–H118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecci, A.; Gomez-Sanchez, C.E.; de Bedners, M.E.; Lantos, C.P.; Cozza, E.N. In vivo stimulation of aldosterone biosynthesis by endothelin: Loci of action and effects of doses and infusion rate. J. Steroid Biochem. Mol. Biol. 1993, 45, 555–561. [Google Scholar] [CrossRef]
- Andreis, P.G.; Neri, G.; Tortorella, C.; Aragona, F.; Rossi, G.P.; Nussdorfer, G.G. Mechanisms transducing the aldosterone secretagogue signal of endothelins in the human adrenal cortex. Peptides 2002, 23, 561–566. [Google Scholar] [CrossRef]
- Yamamoto, T.; Kimura, T.; Ota, K.; Shoji, M.; Inoue, M.; Sato, K.; Ohta, M.; Yoshinaga, K. Central effects of endothelin-1 on vasopressin and atrial natriuretic peptide release and cardiovascular and renal function in conscious rats. J. Cardiovasc. Pharmacol. 1991, 17, S316–S318. [Google Scholar] [CrossRef] [PubMed]
- Rossi, N.F. Regulation of vasopressin secretion by ETA and ETB receptors in compartmentalized rat hypothalamo-neurohypophysial explants. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E535–E541. [Google Scholar] [CrossRef] [Green Version]
- Ohara-Imaizumi, M.; Kumakura, K. Dynamics of the secretory response evoked by endothelin-1 in adrenal chromaffin cells. J. Cardiovasc. Pharmacol. 1991, 17, S156–S158. [Google Scholar] [CrossRef]
- Yoshida, K.; Yasujima, M.; Kohzuki, M.; Tsunoda, K.; Kudo, K.; Kanazawa, M.; Yabe, T.; Abe, K.; Yoshinaga, K. Chronic synergistic effect of endothelin-1 and angiotensin II on blood pressure in conscious rats. J. Cardiovasc. Pharmacol. 1991, 17, S514–S516. [Google Scholar] [CrossRef]
- Imai, T.; Hirata, Y.; Emori, T.; Yanagisawa, M.; Masaki, T.; Marumo, F. Induction of endothelin-1 gene by angiotensin and vasopressin in endothelial cells. Hypertension 1992, 19, 753–757. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, A.V.; Smith, P. Cardiovascular responses induced by endothelin microinjection into area postrema. Regul. Pept. 1990, 27, 75–85. [Google Scholar] [CrossRef]
- Li, D.P.; Fan, Z.Z.; He, R.R. Modulatory effects of endothelin on carotid baroreflex in anesthetized rats. Acta Physiol. Sin. 1998, 50, 169–175. [Google Scholar]
- Chen, A.D.; Xiong, X.Q.; Gan, X.B.; Zhang, F.; Zhou, Y.B.; Gao, X.Y.; Han, Y. Endothelin-1 in paraventricular nucleus modulates cardiac sympathetic afferent reflex and sympathetic activity in rats. PLoS ONE 2012, 7, e40748. [Google Scholar] [CrossRef] [Green Version]
- Rossi, N.F.; Chen, H. PVN lesions prevent the endothelin 1-induced increase in arterial pressure and vasopressin. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E349–E356. [Google Scholar] [CrossRef]
- Bruno, R.M.; Sudano, I.; Ghiadoni, L.; Masi, L.; Taddei, S. Interactions between sympathetic nervous system and endogenous endothelin in patients with essential hypertension. Hypertension 2011, 57, 79–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, M.; Yanagisawa, M. Endothelin: 30 years from discovery to therapy. Hypertension 2019, 74, 1232–1265. [Google Scholar] [CrossRef] [PubMed]
- McCoy, E.K.; Lisenby, K.M. Aprocitentan (a dual endothelin-receptor antagonist) for treatment-resistant hypertension. J. Cardiovasc. Pharmacol. 2021, 77, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Enevoldsen, F.C.; Sahana, J.; Wehland, M.; Grimm, D.; Infanger, M.; Krüger, M. Endothelin receptor antagonists: Status quo and future perspectives for targeted therapy. J. Clin. Med. 2020, 9, 824. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Results | Significance |
|---|---|---|
| Saito, 1990 [47] | Patients with essential HTN showed a significant elevation in the plasma ET-1 level compared with age-matched control subjects. | p < 0.01 |
| Shichiri, 1990 [48] | Patients with essential HTN had significantly higher plasma ET-1 levels than normal subjects. | p < 0.025 |
| Oishi, 1994 [34] | In patients with pheochromocytoma, the hypertensive group had higher ET-1 than the normotensive group. Elevated plasma ET-1 concentrations returned to normal levels after surgical resection of the tumor. | Higher, but NS |
| Parrinello, 1996 [49] | ET-1 levels were significantly higher in obese hypertensives and obese normotensives than in lean normotensives. In addition, ET-1 levels were significantly higher in obese hypertensives than in obese normotensives. | p < 0.05 |
| Amoroso, 1996 [50] | Patients with HTN had significantly higher plasma ET-1 concentration than normal subjects. | p < 0.02 |
| Schneider, 2000 [51] | Basal ET-1 was significantly higher in hypertensive than in normotensive subjects, both in venous and arterial samples. There was no significant difference between venous and arterial ET-1 concentrations. | p < 0.01 |
| Parissis, 2001 [52] | Patients with HTN showed significantly higher levels of ET-1 compared with normotensive controls. | p < 0.01 |
| Kostov, 2014 [53] | Serum levels of ET-1 are significantly higher in patients with mild and severe HTN compared to the control group. | p < 0.02 |
| Gu, 2015 [54] | Plasma ET-1 levels were higher in hypertensives than in controls. | p < 0.001 |
| Kostov, 2016 [55] | Serum ET-1 concentrations were significantly higher in hypertensive patients with type 2 diabetes than in prehypertensive patients with diabetes and healthy normotensive controls. | p < 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostov, K. The Causal Relationship between Endothelin-1 and Hypertension: Focusing on Endothelial Dysfunction, Arterial Stiffness, Vascular Remodeling, and Blood Pressure Regulation. Life 2021, 11, 986. https://doi.org/10.3390/life11090986
Kostov K. The Causal Relationship between Endothelin-1 and Hypertension: Focusing on Endothelial Dysfunction, Arterial Stiffness, Vascular Remodeling, and Blood Pressure Regulation. Life. 2021; 11(9):986. https://doi.org/10.3390/life11090986
Chicago/Turabian StyleKostov, Krasimir. 2021. "The Causal Relationship between Endothelin-1 and Hypertension: Focusing on Endothelial Dysfunction, Arterial Stiffness, Vascular Remodeling, and Blood Pressure Regulation" Life 11, no. 9: 986. https://doi.org/10.3390/life11090986