
Subcellular Localization of Fad1p in Saccharomyces cerevisiae: A Choice at Post-Transcriptional Level?

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Yeast Strains and Recombinant Multicopy Plasmids
2.3. Media and Growth Conditions
2.4. Preparation of Cellular Extracts
2.5. Preparation of Spheroplasts, Cytoplasm, Mitochondria, and Submitochondrial Fractions
2.6. Western Blotting
2.7. RNA Isolation
2.8. 3′RACE Analysis
2.9. Semiquantitative RT-PCR Assay
2.10. Other Assays
3. Results
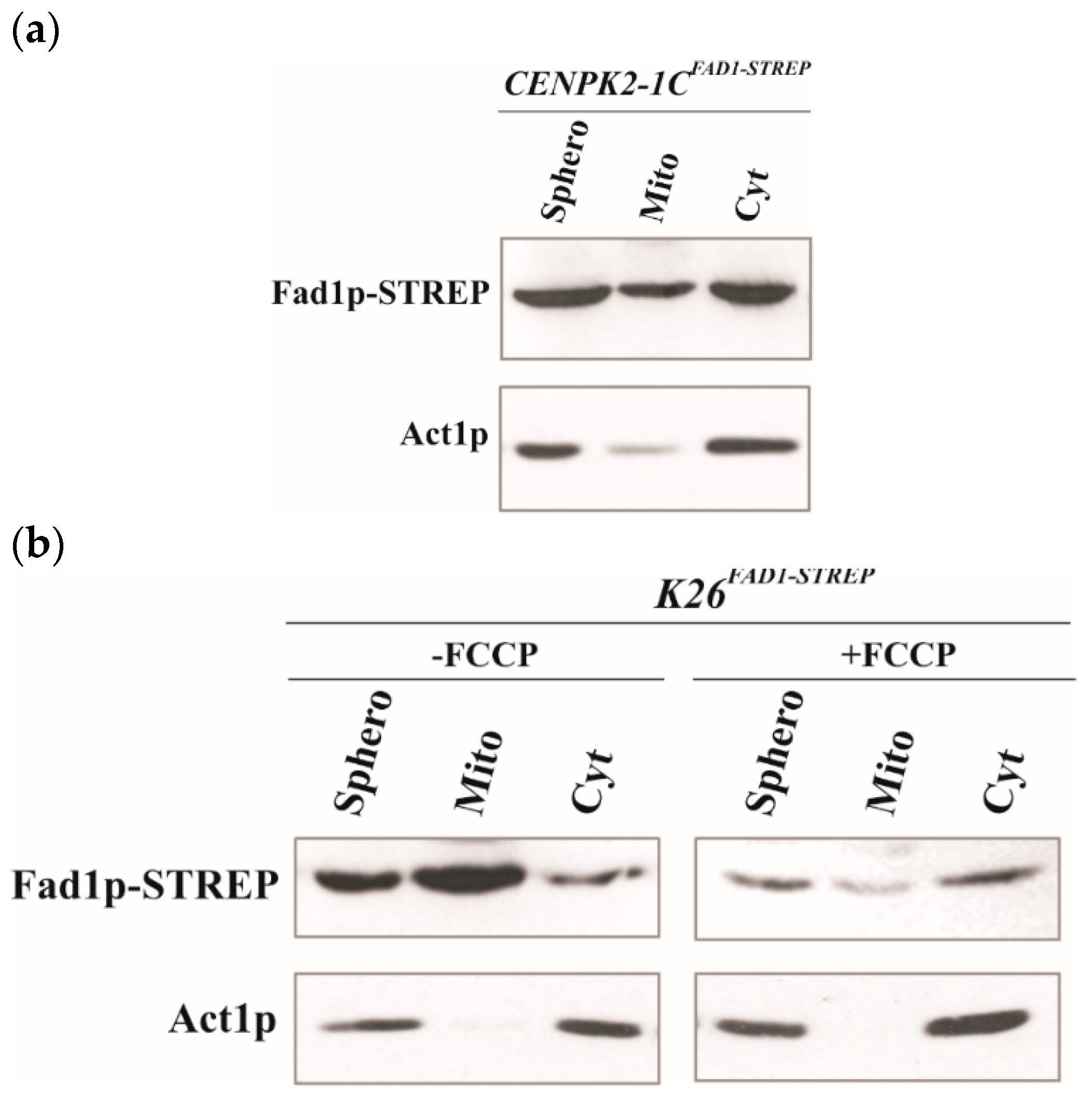
3.1. Mitochondrial Localization of Recombinant Fad1p
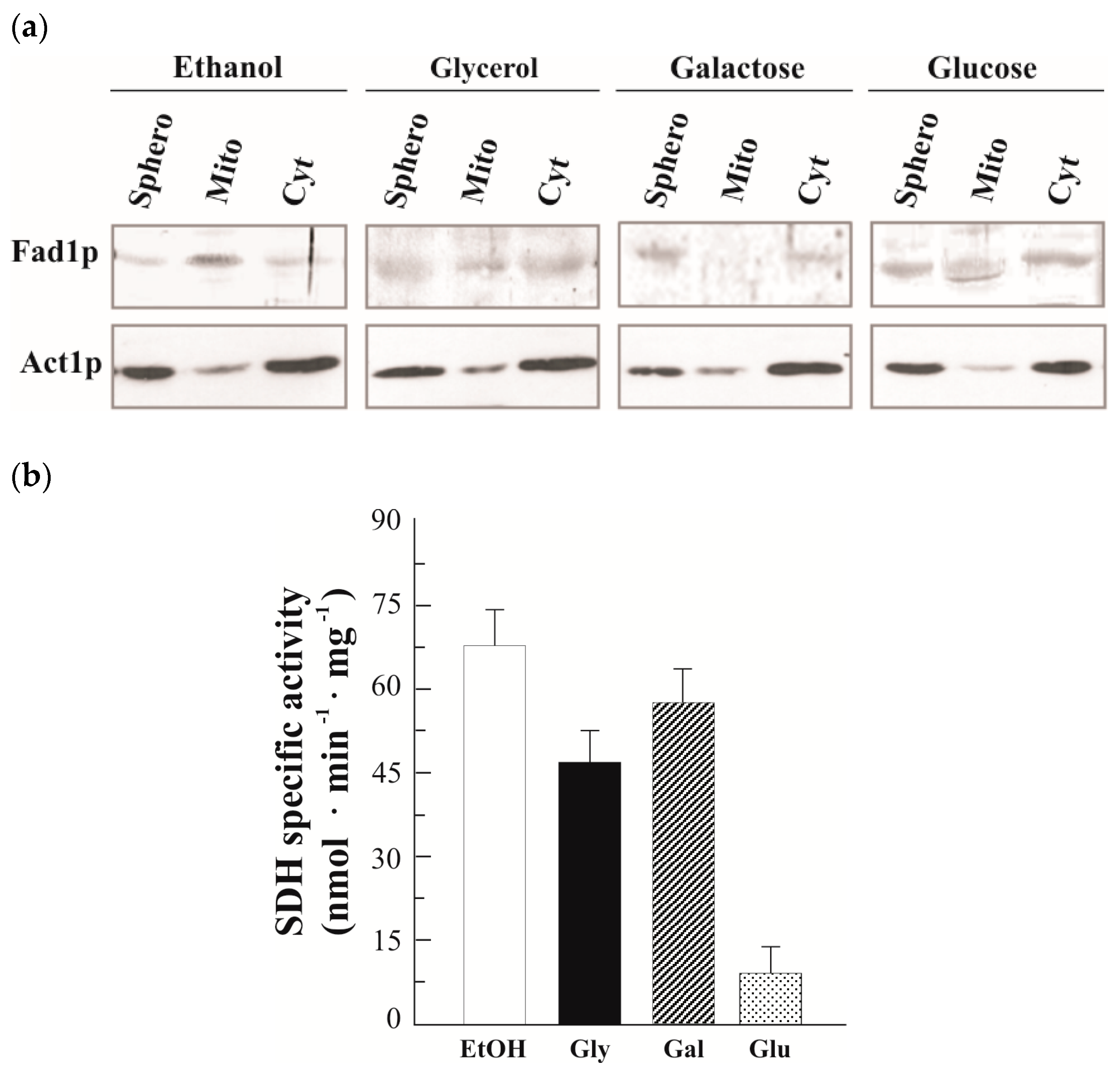
3.2. Natural Fad1p Exists in Mitochondria
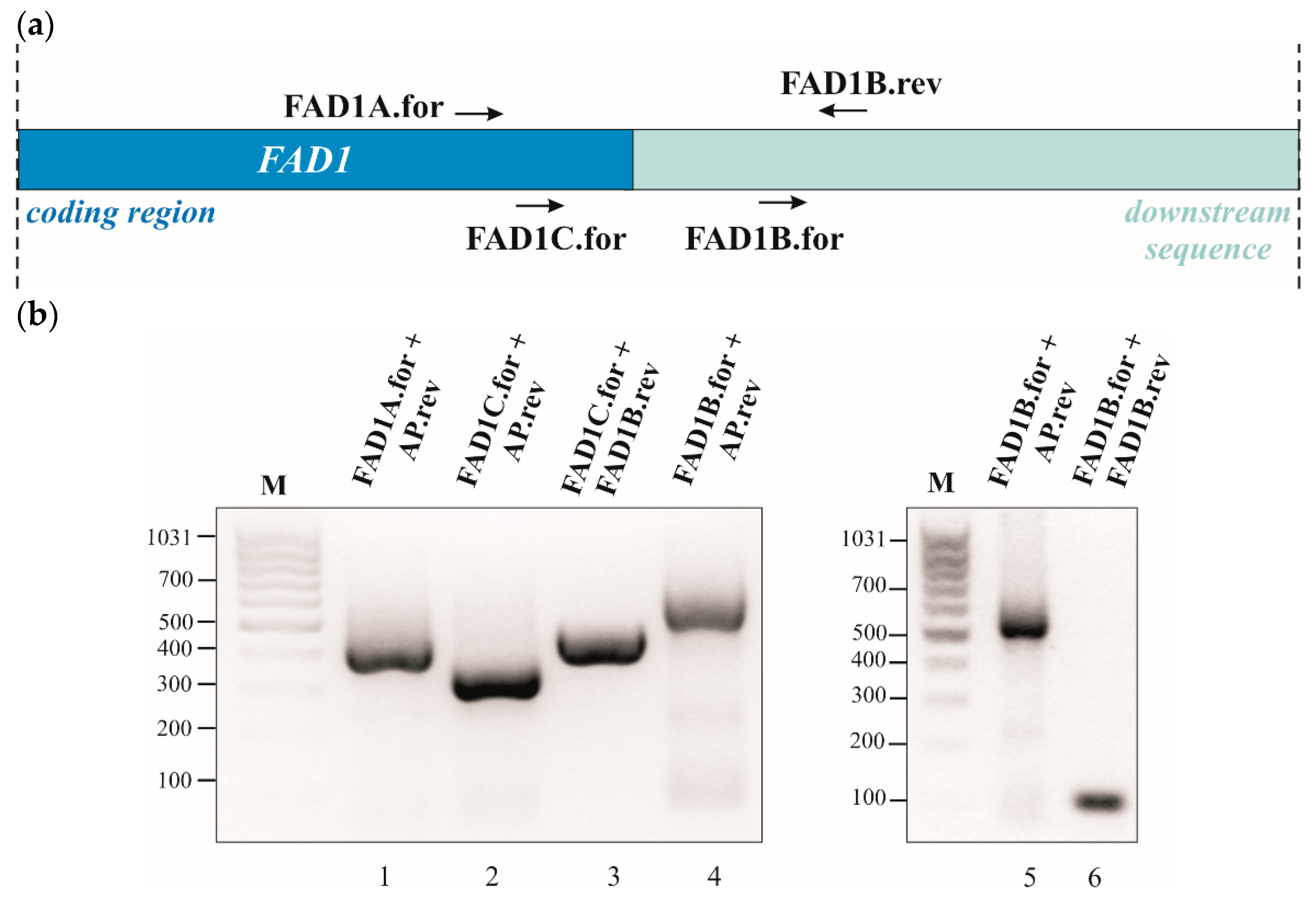
3.3. Detection of Two FAD1 mRNAs with Different 3′UTR and Their Carbon Source Dependence Profile
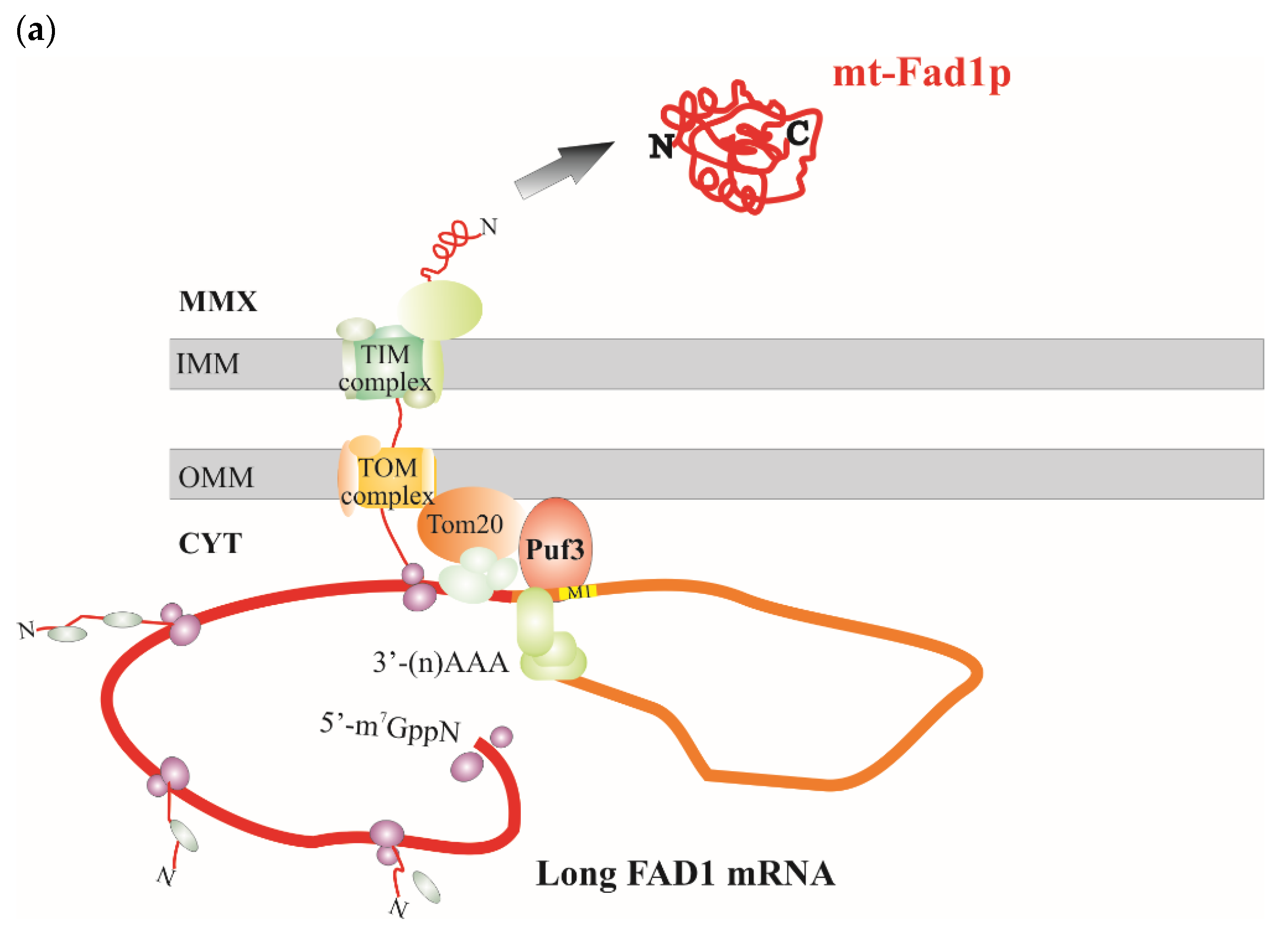
3.4. A Mitochondrial Localization Motif in FAD1 3′UTRs
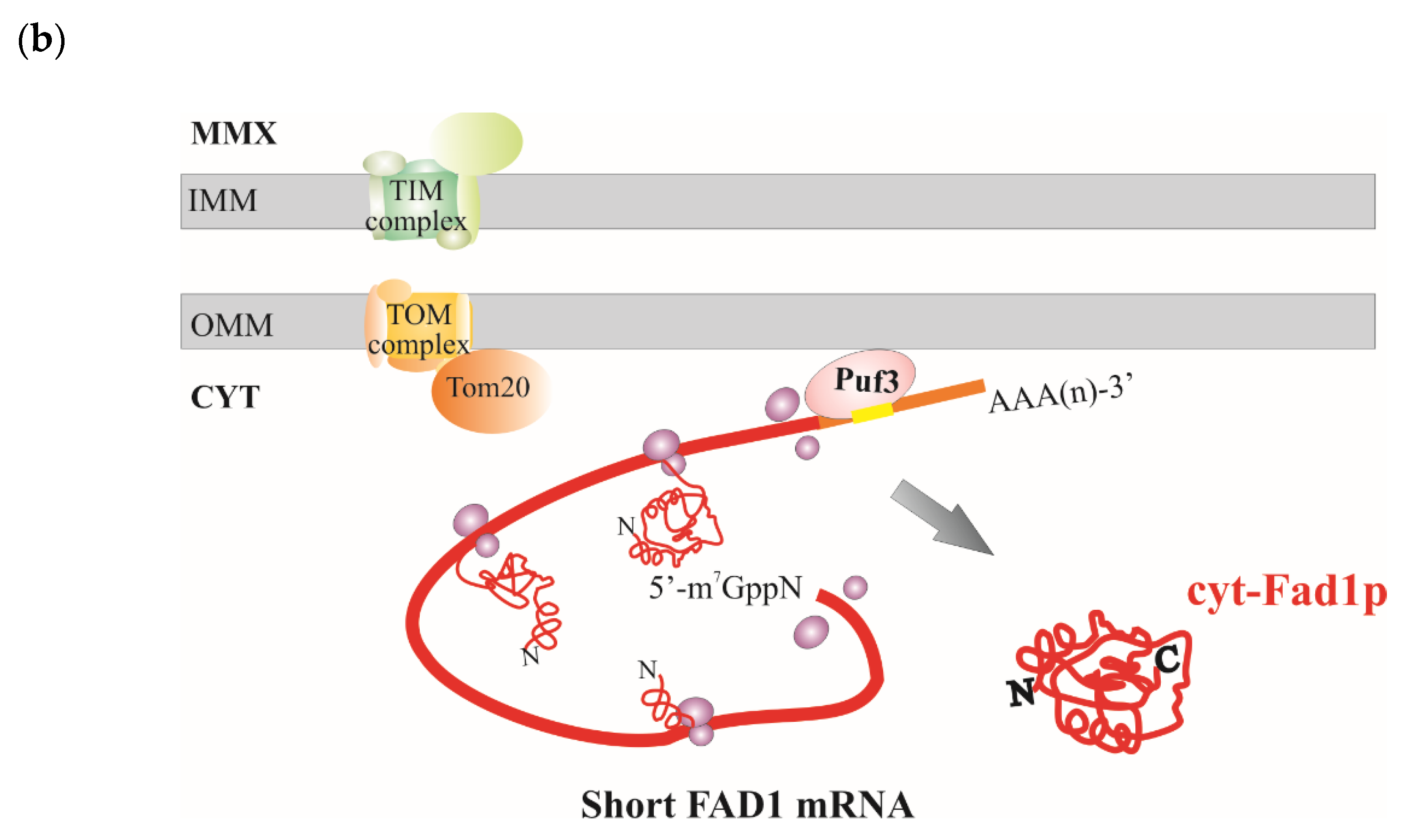
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tolomeo, M.; Nisco, A.; Leone, P.; Barile, M. Development of Novel Experimental Models to Study Flavoproteome Alterations in Human Neuromuscular Diseases: The Effect of Rf Therapy. Int. J. Mol. Sci. 2020, 21, 5310. [Google Scholar] [CrossRef]
- Joosten, V.; Van Berkel, W.J. Flavoenzymes. Curr. Opin. Chem. Biol. 2007, 11, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Macheroux, P.; Kappes, B.; Ealick, S.E. Flavogenomics—A genomic and structural view of flavin-dependent proteins. FEBS J. 2011, 278, 2625–2634. [Google Scholar] [CrossRef] [PubMed]
- Barile, M.; Giancaspero, T.A.; Leone, P.; Galluccio, M.; Indiveri, C. Riboflavin transport and metabolism in humans. J. Inherit. Metab. Dis. 2016, 39, 545–557. [Google Scholar] [CrossRef]
- Pimentel, C.; Batista-Nascimento, L.; Rodrigues-Pousada, C.; Menezes, R.A. Oxidative stress in Alzheimer’s and Parkinson’s diseases: Insights from the yeast Saccharomyces cerevisiae. Oxidative Med. Cell. Longev. 2012, 2012, 132146. [Google Scholar] [CrossRef] [Green Version]
- Botstein, D.; Fink, G.R. Yeast: An experimental organism for 21st Century biology. Genetics 2011, 189, 695–704. [Google Scholar] [CrossRef] [Green Version]
- Tenreiro, S.; Outeiro, T.F. Simple is good: Yeast models of neurodegeneration. FEMS Yeast Res. 2010, 10, 970–979. [Google Scholar] [CrossRef] [PubMed]
- Bastow, E.L.; Gourlay, C.W.; Tuite, M.F. Using yeast models to probe the molecular basis of amyotrophic lateral sclerosis. Biochem. Soc. Trans. 2012, 39, 1482–1487. [Google Scholar] [CrossRef] [PubMed]
- Reihl, P.; Stolz, J. The monocarboxylate transporter homolog Mch5p catalyzes riboflavin (vitamin B2) uptake in Saccharomyces cerevisiae. J. Biol. Chem. 2005, 280, 39809–39817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriyama, Y. Riboflavin transporter is finally identified. J. Biochem. 2011, 150, 341–343. [Google Scholar] [CrossRef]
- Yonezawa, A.; Inui, K. Novel riboflavin transporter family RFVT/SLC52: Identification, nomenclature, functional characterization and genetic diseases of RFVT/SLC52. Mol. Asp. Med. 2013, 34, 693–701. [Google Scholar] [CrossRef]
- O’Callaghan, B.; Bosch, A.M.; Houlden, H. An update on the genetics, clinical presentation, and pathomechanisms of human riboflavin transporter deficiency. J. Inherit. Metab. Dis. 2019, 42, 598–607. [Google Scholar] [CrossRef]
- Yruela, I.; Arilla-Luna, S.; Medina, M.; Contreras-Moreira, B. Evolutionary divergence of chloroplast FAD synthetase proteins. BMC Evol. Biol. 2010, 10, 311. [Google Scholar] [CrossRef] [Green Version]
- Giancaspero, T.A.; Colella, M.; Brizio, C.; Difonzo, G.; Fiorino, G.M.; Leone, P.; Brandsch, R.; Bonomi, F.; Iametti, S.; Barile, M. Remaining challenges in cellular flavin cofactor homeostasis and flavoprotein biogenesis. Front. Chem. 2015, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.A.; Jimenez, A.; Revuelta, J.L. Molecular characterization of FMN1, the structural gene for the monofunctional flavokinase of Saccharomyces cerevisiae. J. Biol. Chem. 2000, 275, 28618–28624. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Repetto, B.; Glerum, D.M.; Tzagoloff, A. Cloning and characterization of FAD1, the structural gene for flavin adenine dinucleotide synthetase of Saccharomyces cerevisiae. Mol. Cell. Biol. 1995, 15, 264–271. [Google Scholar] [CrossRef] [Green Version]
- Bauer, S.; Kemter, K.; Bacher, A.; Huber, R.; Fischer, M.; Steinbacher, S. Crystal structure of Schizosaccharomyces pombe riboflavin kinase reveals a novel ATP and riboflavin-binding fold. J. Mol. Biol. 2003, 326, 1463–1473. [Google Scholar] [CrossRef]
- Karthikeyan, S.; Zhou, Q.; Mseeh, F.; Grishin, N.V.; Osterman, A.L.; Zhang, H. Crystal structure of human riboflavin kinase reveals a beta barrel fold and a novel active site arch. Structure 2003, 11, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Karthikeyan, S.; Zhou, Q.; Osterman, A.L.; Zhang, H. Ligand binding-induced conformational changes in riboflavin kinase: Structural basis for the ordered mechanism. Biochemistry 2003, 42, 12532–12538. [Google Scholar] [CrossRef] [PubMed]
- Leulliot, N.; Blondeau, K.; Keller, J.; Ulryck, N.; Quevillon-Cheruel, S.; Van Tilbeurgh, H. Crystal structure of yeast FAD synthetase (Fad1) in complex with FAD. J. Mol. Biol. 2010, 398, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Brizio, C.; Galluccio, M.; Wait, R.; Torchetti, E.M.; Bafunno, V.; Accardi, R.; Gianazza, E.; Indiveri, C.; Barile, M. Over-expression in Escherichia coli and characterization of two recombinant isoforms of human FAD synthetase. Biochem. Biophys. Res. Commun. 2006, 344, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Leone, P.; Galluccio, M.; Brizio, C.; Barbiroli, A.; Iametti, S.; Indiveri, C.; Barile, M. The hidden side of the human FAD synthase 2. Int. J. Biol. Macromol. 2019, 138, 986–995. [Google Scholar] [CrossRef]
- Giancaspero, T.A.; Locato, V.; Barile, M. A regulatory role of NAD redox status on flavin cofactor homeostasis in S. cerevisiae mitochondria. Oxidative Med. Cell. Longev. 2013, 2013, 612784. [Google Scholar] [CrossRef] [Green Version]
- Vergani, L.; Barile, M.; Angelini, C.; Burlina, A.B.; Nijtmans, L.; Freda, M.P.; Brizio, C.; Zerbetto, E.; Dabbeni-Sala, F. Riboflavin therapy. Biochemical heterogeneity in two adult lipid storage myopathies. Brain A J. Neurol. 1999, 122 Pt 12, 2401–2411. [Google Scholar] [CrossRef] [Green Version]
- Gianazza, E.; Vergani, L.; Wait, R.; Brizio, C.; Brambilla, D.; Begum, S.; Giancaspero, T.A.; Conserva, F.; Eberini, I.; Bufano, D.; et al. Coordinated and reversible reduction of enzymes involved in terminal oxidative metabolism in skeletal muscle mitochondria from a riboflavin-responsive, multiple acyl-CoA dehydrogenase deficiency patient. Electrophoresis 2006, 27, 1182–1198. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.K.J.; Konarikova, E.; Giancaspero, T.A.; Mosegaard, S.; Boczonadi, V.; Matakovic, L.; Veauville-Merllie, A.; Terrile, C.; Schwarzmayr, T.; Haack, T.B.; et al. Riboflavin-Responsive and -Non-responsive Mutations in FAD Synthase Cause Multiple Acyl-CoA Dehydrogenase and Combined Respiratory-Chain Deficiency. Am. J. Hum. Genet. 2016, 98, 1130–1145. [Google Scholar] [CrossRef] [Green Version]
- Gregersen, N.; Andresen, B.S.; Pedersen, C.B.; Olsen, R.K.; Corydon, T.J.; Bross, P. Mitochondrial fatty acid oxidation defects--remaining challenges. J. Inherit. Metab. Dis. 2008, 31, 643–657. [Google Scholar] [CrossRef]
- Leone, P.; Galluccio, M.; Barbiroli, A.; Eberini, I.; Tolomeo, M.; Vrenna, F.; Gianazza, E.; Iametti, S.; Bonomi, F.; Indiveri, C.; et al. Bacterial Production, Characterization and Protein Modeling of a Novel Monofuctional Isoform of FAD Synthase in Humans: An Emergency Protein? Molecules 2018, 23, 116. [Google Scholar] [CrossRef] [Green Version]
- Torchetti, E.M.; Brizio, C.; Colella, M.; Galluccio, M.; Giancaspero, T.A.; Indiveri, C.; Roberti, M.; Barile, M. Mitochondrial localization of human FAD synthetase isoform 1. Mitochondrion 2010, 10, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Giancaspero, T.A.; Busco, G.; Panebianco, C.; Carmone, C.; Miccolis, A.; Liuzzi, G.M.; Colella, M.; Barile, M. FAD synthesis and degradation in the nucleus create a local flavin cofactor pool. J. Biol. Chem. 2013, 288, 29069–29080. [Google Scholar] [CrossRef] [Green Version]
- Gudipati, V.; Koch, K.; Lienhart, W.D.; Macheroux, P. The flavoproteome of the yeast Saccharomyces cerevisiae. Biochim. Et Biophys. Acta 2014, 1844, 535–544. [Google Scholar] [CrossRef] [Green Version]
- Hao, H.X.; Khalimonchuk, O.; Schraders, M.; Dephoure, N.; Bayley, J.P.; Kunst, H.; Devilee, P.; Cremers, C.W.; Schiffman, J.D.; Bentz, B.G.; et al. SDH5, a gene required for flavination of succinate dehydrogenase, is mutated in paraganglioma. Science 2009, 325, 1139–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghezzi, D.; Goffrini, P.; Uziel, G.; Horvath, R.; Klopstock, T.; Lochmuller, H.; D’Adamo, P.; Gasparini, P.; Strom, T.M.; Prokisch, H.; et al. SDHAF1, encoding a LYR complex-II specific assembly factor, is mutated in SDH-defective infantile leukoencephalopathy. Nat. Genet. 2009, 41, 654–656. [Google Scholar] [CrossRef]
- Robinson, K.M.; Lemire, B.D. Covalent attachment of FAD to the yeast succinate dehydrogenase flavoprotein requires import into mitochondria, presequence removal, and folding. J. Biol. Chem. 1996, 271, 4055–4060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Winge, D.R. Emerging concepts in the flavinylation of succinate dehydrogenase. Biochim. Biophys. Acta 2013, 1827, 627–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutter, J.; Winge, D.R.; Schiffman, J.D. Succinate dehydrogenase-Assembly, regulation and role in human disease. Mitochondrion 2010, 10, 393–401. [Google Scholar] [CrossRef] [Green Version]
- Schiff, M.; Veauville-Merllie, A.; Su, C.H.; Tzagoloff, A.; Rak, M.; Ogier de Baulny, H.; Boutron, A.; Smedts-Walters, H.; Romero, N.B.; Rigal, O.; et al. SLC25A32 Mutations and Riboflavin-Responsive Exercise Intolerance. N. Engl. J. Med. 2016, 374, 795–797. [Google Scholar] [CrossRef] [Green Version]
- Hellebrekers, D.; Sallevelt, S.; Theunissen, T.E.J.; Hendrickx, A.T.M.; Gottschalk, R.W.; Hoeijmakers, J.G.J.; Habets, D.D.; Bierau, J.; Schoonderwoerd, K.G.; Smeets, H.J.M. Novel SLC25A32 mutation in a patient with a severe neuromuscular phenotype. Eur. J. Hum. Genet. 2017, 25, 886–888. [Google Scholar] [CrossRef]
- Tzagoloff, A.; Jang, J.; Glerum, D.M.; Wu, M. FLX1 codes for a carrier protein involved in maintaining a proper balance of flavin nucleotides in yeast mitochondria. J. Biol. Chem. 1996, 271, 7392–7397. [Google Scholar] [CrossRef] [Green Version]
- Bafunno, V.; Giancaspero, T.A.; Brizio, C.; Bufano, D.; Passarella, S.; Boles, E.; Barile, M. Riboflavin uptake and FAD synthesis in Saccharomyces cerevisiae mitochondria: Involvement of the Flx1p carrier in FAD export. J. Biol. Chem. 2004, 279, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Barile, M.; Passarella, S.; Bertoldi, A.; Quagliariello, E. Flavin adenine dinucleotide synthesis in isolated rat liver mitochondria caused by imported flavin mononucleotide. Arch. Biochem. Biophys. 1993, 305, 442–447. [Google Scholar] [CrossRef]
- Giancaspero, T.A.; Wait, R.; Boles, E.; Barile, M. Succinate dehydrogenase flavoprotein subunit expression in Saccharomyces cerevisiae--involvement of the mitochondrial FAD transporter, Flx1p. FEBS J. 2008, 275, 1103–1117. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. Frozen competent yeast cells that can be transformed with high efficiency using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 1–4. [Google Scholar] [CrossRef]
- Giancaspero, T.A.; Locato, V.; De Pinto, M.C.; De Gara, L.; Barile, M. The occurrence of riboflavin kinase and FAD synthetase ensures FAD synthesis in tobacco mitochondria and maintenance of cellular redox status. FEBS J. 2009, 276, 219–231. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Pallotta, M.L.; Brizio, C.; Fratianni, A.; De Virgilio, C.; Barile, M.; Passarella, S. Saccharomyces cerevisiae mitochondria can synthesise FMN and FAD from externally added riboflavin and export them to the extramitochondrial phase. FEBS Lett. 1998, 428, 245–249. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.; Mahlke, K.; Pfanner, N. Role of an energized inner membrane in mitochondrial protein import. Delta psi drives the movement of presequences. J. Biol. Chem. 1991, 266, 18051–18057. [Google Scholar] [CrossRef]
- Traglia, H.M.; O’Connor, J.P.; Tung, K.S.; Dallabrida, S.; Shen, W.C.; Hopper, A.K. Nucleus-associated pools of Rna1p, the Saccharomyces cerevisiae Ran/TC4 GTPse activating protein involved in nucleus/cytosol transit. Proc. Natl. Acad. Sci. USA 1996, 93, 7667–7672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leadsham, J.E.; Gourlay, C.W. Cytoskeletal induced apoptosis in yeast. Biochim. Biophys. Acta 2008, 1783, 1406–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardo, A.; Cereghino, G.P.; Scheffler, I.E. Control of mRNA turnover as a mechanism of glucose repression in Saccharomyces cerevisiae. Mol. Cell. Biol. 1992, 12, 2941–2948. [Google Scholar] [CrossRef] [Green Version]
- Prieto, S.; De la Cruz, B.J.; Scheffler, I.E. Glucose-regulated turnover of mRNA and the influence of poly(A) tail length on half-life. J. Biol. Chem. 2000, 275, 14155–14166. [Google Scholar] [CrossRef] [Green Version]
- Polakis, E.S.; Bartley, W. Changes in the enzyme activities of Saccharomyces cerevisiae during aerobic growth on different carbon sources. Biochem. J. 1965, 97, 284–297. [Google Scholar] [CrossRef] [Green Version]
- Zahedi, R.P.; Sickmann, A.; Boehm, A.M.; Winkler, C.; Zufall, N.; Schonfisch, B.; Guiard, B.; Pfanner, N.; Meisinger, C. Proteomic analysis of the yeast mitochondrial outer membrane reveals accumulation of a subclass of preproteins. Mol. Biol. Cell 2006, 17, 1436–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marc, P.; Margeot, A.; Devaux, F.; Blugeon, C.; Corral-Debrinski, M.; Jacq, C. Genome-wide analysis of mRNAs targeted to yeast mitochondria. EMBO Rep. 2002, 3, 159–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.U.; Fisher, P.R. Import of nuclear-encoded mitochondrial proteins: A cotranslational perspective. Int. Rev. Cell Mol. Biol. 2009, 273, 49–68. [Google Scholar]
- Kellems, R.E.; Allison, V.F.; Butow, R.A. Cytoplasmic type 80S ribosomes associated with yeast mitochondria. IV. Attachment of ribosomes to the outer membrane of isolated mitochondria. J. Cell Biol. 1975, 65, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Eliyahu, E.; Lesnik, C.; Arava, Y. The protein chaperone Ssa1 affects mRNA localization to the mitochondria. FEBS Lett. 2012, 586, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Dichtl, B.; Keller, W. Recognition of polyadenylation sites in yeast pre-mRNAs by cleavage and polyadenylation factor. The EMBO J. 2001, 20, 3197–3209. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Hyman, L.; Moore, C. Formation of mRNA 3’ ends in eukaryotes: Mechanism, regulation, and interrelationships with other steps in mRNA synthesis. Microbiol. Mol. Biol. Rev. 1999, 63, 405–445. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Han, H.; Li, J.; Wong, L. DNAFSMiner: A web-based software toolbox to recognize two types of functional sites in DNA sequences. Bioinformatics 2005, 21, 671–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shalgi, R.; Lapidot, M.; Shamir, R.; Pilpel, Y. A catalog of stability-associated sequence elements in 3’ UTRs of yeast mRNAs. Genome Biol. 2005, 6, R86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellis, M.; Patterson, N.; Endrizzi, M.; Birren, B.; Lander, E.S. Sequencing and comparison of yeast species to identify genes and regulatory elements. Nature 2003, 423, 241–254. [Google Scholar] [CrossRef]
- Jin, C.; Myers, A.M.; Tzagoloff, A. Cloning and characterization of MRP10, a yeast gene coding for a mitochondrial ribosomal protein. Curr. Genet. 1997, 31, 228–234. [Google Scholar] [CrossRef]
- Kitakawa, M.; Graack, H.R.; Grohmann, L.; Goldschmidt-Reisin, S.; Herfurth, E.; Wittmann-Liebold, B.; Nishimura, T.; Isono, K. Identification and characterization of the genes for mitochondrial ribosomal proteins of Saccharomyces cerevisiae. Eur. J. Biochem. 1997, 245, 449–456. [Google Scholar] [CrossRef]
- Yogev, O.; Pines, O. Dual targeting of mitochondrial proteins: Mechanism, regulation and function. Biochim. Et Biophys. Acta 2011, 1808, 1012–1020. [Google Scholar] [CrossRef] [Green Version]
- Bader, G.; Enkler, L.; Araiso, Y.; Hemmerle, M.; Binko, K.; Baranowska, E.; De Craene, J.O.; Ruer-Laventie, J.; Pieters, J.; Tribouillard-Tanvier, D.; et al. Assigning mitochondrial localization of dual localized proteins using a yeast Bi-Genomic Mitochondrial-Split-GFP. eLife 2020, 9, e56649. [Google Scholar] [CrossRef] [PubMed]
- Ben-Menachem, R.; Pines, O. Detection of Dual Targeting and Dual Function of Mitochondrial Proteins in Yeast. Methods Mol. Biol. 2017, 1567, 179–195. [Google Scholar] [CrossRef] [PubMed]
- Ben-Menachem, R.; Tal, M.; Shadur, T.; Pines, O. A third of the yeast mitochondrial proteome is dual localized: A question of evolution. Proteomics 2011, 11, 4468–4476. [Google Scholar] [CrossRef]
- Kisslov, I.; Naamati, A.; Shakarchy, N.; Pines, O. Dual-targeted proteins tend to be more evolutionarily conserved. Mol. Biol. Evol. 2014, 31, 2770–2779. [Google Scholar] [CrossRef] [Green Version]
- Dinur-Mills, M.; Tal, M.; Pines, O. Dual targeted mitochondrial proteins are characterized by lower MTS parameters and total net charge. PLoS ONE 2008, 3, e2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barile, M.; University of Bari ‘Aldo Moro’, Bari, Italy. Personal communication, 2021.
- Lashkevich, K.A.; Dmitriev, S.E. mRNA Targeting, Transport and Local Translation in Eukaryotic Cells: From the Classical View to a Diversity of New Concepts. Mol. Biol. 2021, 1–31. [Google Scholar] [CrossRef]
- Andreassi, C.; Riccio, A. To localize or not to localize: mRNA fate is in 3’UTR ends. Trends Cell Biol. 2009, 19, 465–474. [Google Scholar] [CrossRef]
- Sparks, K.A.; Dieckmann, C.L. Regulation of poly(A) site choice of several yeast mRNAs. Nucleic Acids Res. 1998, 26, 4676–4687. [Google Scholar] [CrossRef] [Green Version]
- Turner, R.E.; Pattison, A.D.; Beilharz, T.H. Alternative polyadenylation in the regulation and dysregulation of gene expression. Semin. Cell Dev. Biol. 2018, 75, 61–69. [Google Scholar] [CrossRef]
- Sylvestre, J.; Vialette, S.; Corral Debrinski, M.; Jacq, C. Long mRNAs coding for yeast mitochondrial proteins of prokaryotic origin preferentially localize to the vicinity of mitochondria. Genome Biol. 2003, 4, R44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horton, P. A simulation study of mRNA diffusion and mitochondrial localization. bioRxiv 2019. [Google Scholar] [CrossRef]
- Gerber, A.P.; Herschlag, D.; Brown, P.O. Extensive association of functionally and cytotopically related mRNAs with Puf family RNA-binding proteins in yeast. PLoS Biol. 2004, 2, e79. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rodriguez, L.J.; Gay, A.C.; Pon, L.A. Puf3p, a Pumilio family RNA binding protein, localizes to mitochondria and regulates mitochondrial biogenesis and motility in budding yeast. J. Cell Biol. 2007, 176, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Rowe, W.; Kershaw, C.J.; Castelli, L.M.; Costello, J.L.; Ashe, M.P.; Grant, C.M.; Sims, P.F.; Pavitt, G.D.; Hubbard, S.J. Puf3p induces translational repression of genes linked to oxidative stress. Nucleic Acids Res. 2014, 42, 1026–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quenault, T.; Lithgow, T.; Traven, A. PUF proteins: Repression, activation and mRNA localization. Trends Cell Biol. 2011, 21, 104–112. [Google Scholar] [CrossRef]
- Saint-Georges, Y.; Garcia, M.; Delaveau, T.; Jourdren, L.; Le Crom, S.; Lemoine, S.; Tanty, V.; Devaux, F.; Jacq, C. Yeast mitochondrial biogenesis: A role for the PUF RNA-binding protein Puf3p in mRNA localization. PLoS ONE 2008, 3, e2293. [Google Scholar] [CrossRef] [Green Version]
- Freeberg, M.A.; Han, T.; Moresco, J.J.; Kong, A.; Yang, Y.C.; Lu, Z.J.; Yates, J.R.; Kim, J.K. Pervasive and dynamic protein binding sites of the mRNA transcriptome in Saccharomyces cerevisiae. Genome Biol. 2013, 14, R13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapointe, C.P.; Stefely, J.A.; Jochem, A.; Hutchins, P.D.; Wilson, G.M.; Kwiecien, N.W.; Coon, J.J.; Wickens, M.; Pagliarini, D.J. Multi-omics Reveal Specific Targets of the RNA-Binding Protein Puf3p and Its Orchestration of Mitochondrial Biogenesis. Cell Syst. 2018, 6, 125–135.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chacinska, A.; Koehler, C.M.; Milenkovic, D.; Lithgow, T.; Pfanner, N. Importing mitochondrial proteins: Machineries and mechanisms. Cell 2009, 138, 628–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, F.; Yanofsky, C. Instruction of translating ribosome by nascent peptide. Science 2002, 297, 1864–1867. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Multicopy Plasmid |
|---|---|
| CEN.PK113-13D (EBY157, WT, K26) | / |
| K26 CTR | p426-HXT7-STREP |
| K26 FAD1-STREP | p426-HXT7-FAD1-STREP |
| CENPK2-1C FAD1-STREP | p426-MET25-FAD1-STREP |
| CENPK2-1C STREP-FAD1 | p426-MET25-STREP-FAD1 |
| Gene | Primer | Sequence |
|---|---|---|
| FAD1 | FAD1A.for | 5′-ATCGGCGGAATTAACAACTCA-3′ |
| FAD1A.rev | 5′-TTGCCAAATGCATGAATGATTT-3′ | |
| FAD1B.for | 5′-GCCTAGCGGC GTGATAGTTAA-3′ | |
| FAD1B.rev | 5′-TGCTGGCTTAGTAACGGAATTG-3′ | |
| FAD1C.for | 5′-CATTTGGCAAGGACGCAGAA-3′ | |
| SDH1 | SDH1.for | 5′-GCCAATTCCTTGTTGGATCTTG-3′ |
| SDH1.rev | 5′-TGGCAACCCAGGCTGTAAAG-3′ | |
| ACT1 | ACT1.for | 5′-TTCCATCCAAGCCGTTTTGT-3′ |
| ACT1.rev | 5′-GGCGTGAGGTAGAGAGAAACCA-3′ | |
| Oligo(dT)-AP | 5′-GACCACGCGTATCGATGTCGAC(T)16V-3′ | |
| AP.rev | 5′-GACCACGCGTATCGATGTCGAC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruni, F.; Giancaspero, T.A.; Oreb, M.; Tolomeo, M.; Leone, P.; Boles, E.; Roberti, M.; Caselle, M.; Barile, M. Subcellular Localization of Fad1p in Saccharomyces cerevisiae: A Choice at Post-Transcriptional Level? Life 2021, 11, 967. https://doi.org/10.3390/life11090967
Bruni F, Giancaspero TA, Oreb M, Tolomeo M, Leone P, Boles E, Roberti M, Caselle M, Barile M. Subcellular Localization of Fad1p in Saccharomyces cerevisiae: A Choice at Post-Transcriptional Level? Life. 2021; 11(9):967. https://doi.org/10.3390/life11090967
Chicago/Turabian StyleBruni, Francesco, Teresa Anna Giancaspero, Mislav Oreb, Maria Tolomeo, Piero Leone, Eckhard Boles, Marina Roberti, Michele Caselle, and Maria Barile. 2021. "Subcellular Localization of Fad1p in Saccharomyces cerevisiae: A Choice at Post-Transcriptional Level?" Life 11, no. 9: 967. https://doi.org/10.3390/life11090967