
Bioactive Natural Compounds with Antiplatelet and Anticoagulant Activity and Their Potential Role in the Treatment of Thrombotic Disorders


Abstract
:1. Introduction
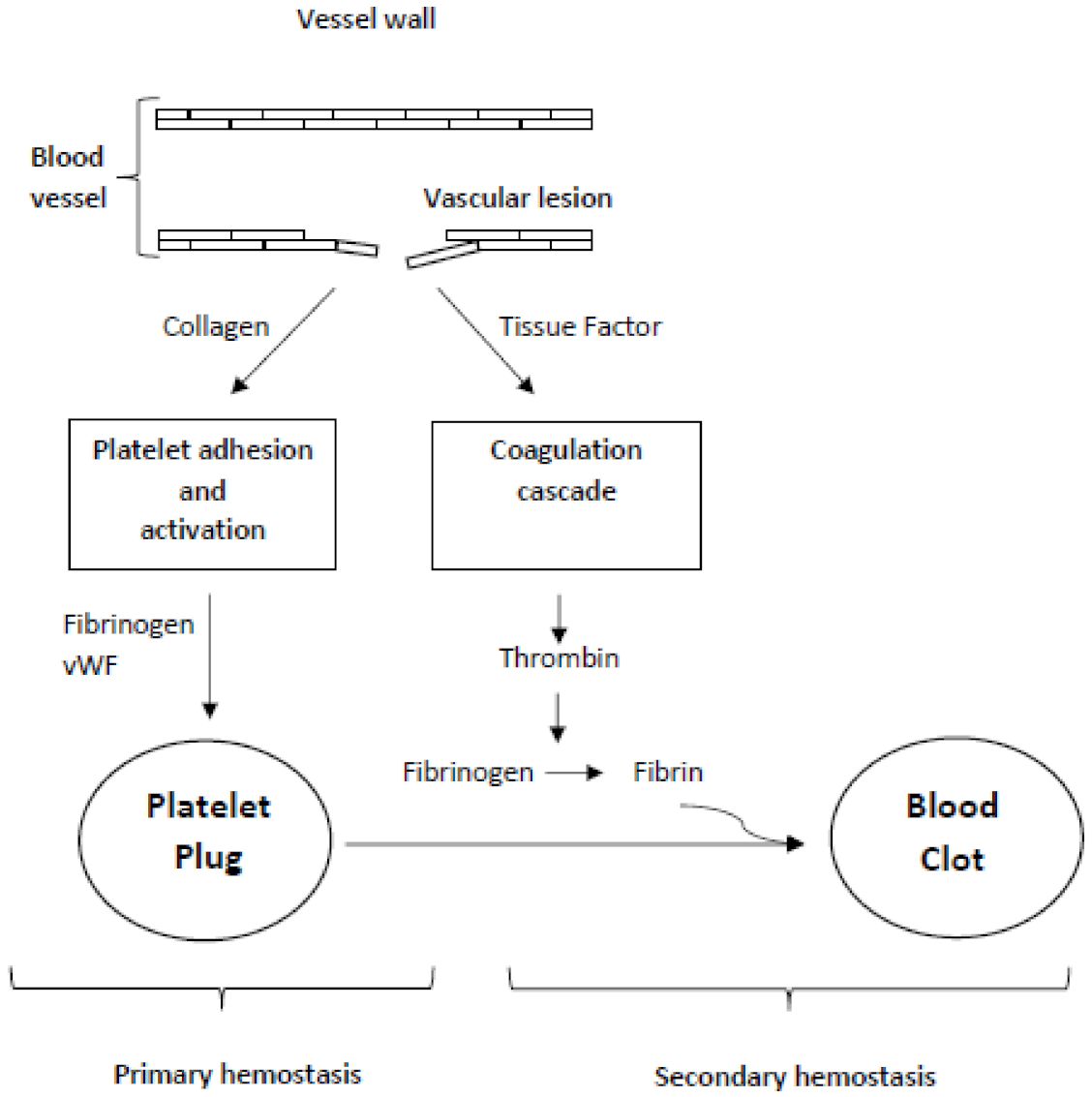
2. Hemostasis
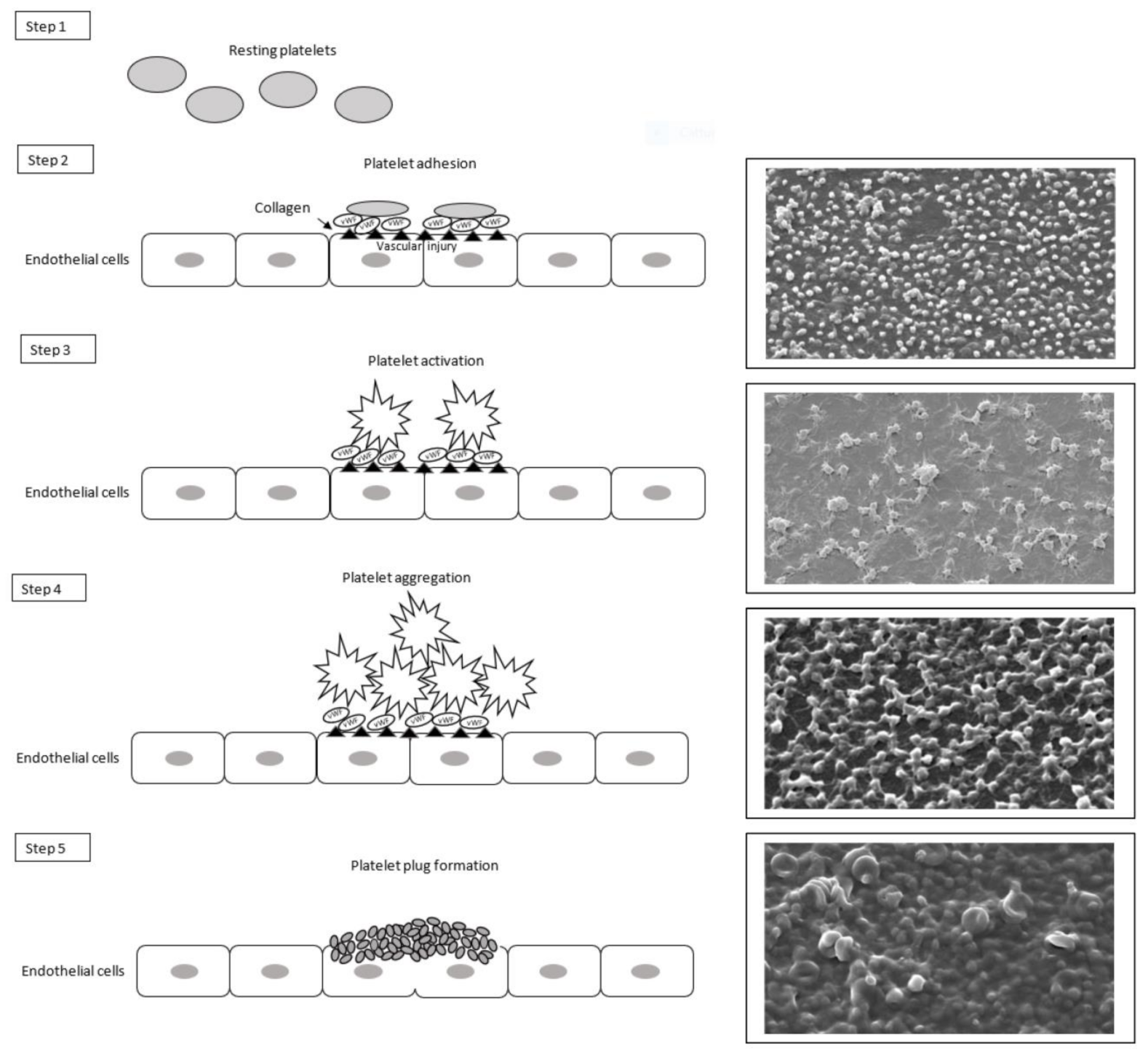
2.1. Primary Hemostasis
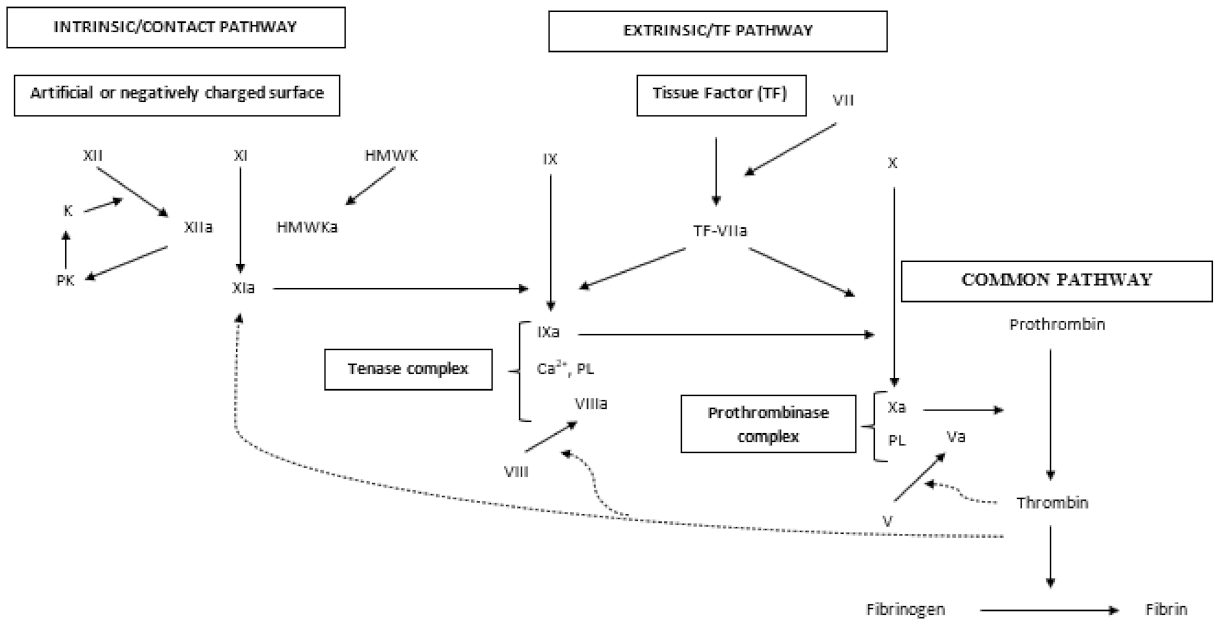
2.2. Secondary Hemostasis
2.2.1. Extrinsic or Tissue Factor Pathway
2.2.2. Intrinsic or Contact Pathway
2.2.3. Tenase Complex
2.2.4. Common Pathway
3. Thrombotic Vascular Diseases
4. Bioactive Compounds from Plants
5. Antiplatelet and Anticoagulant Activity of Plants and Their Bioactive Compounds
5.1. Olea europaea (Oleaceae)
5.2. Chamomilla recutita L. (Asteracee)
5.3. Allium sativum and Allium ursinum (Amaryllidaceae)
5.4. Rosmarinus officinalis (Lamiaceae)
5.5. Fragaria vesca, Echinacea purpurea and Erigeron canadensis L (Rosaceae and Asteraceae)
5.6. Thymus atlanticus and Thymus zygis (Lamiaceae)
5.7. Licania rigida (Chrysobalanaceae)
5.8. Fumaria officinalis (Papaveracee)
5.9. Careya arborea (Lecythidaceae)
5.10. Viola yedoensis (Violaceae)
5.11. Euphorbia resinifera (Euforbiacee)
5.12. Orobanche caryophyllacea, Phelipanche arenaria, and Phelipanche ramosa (Orobanchaceae)
5.13. Genipa americana (Rubiaceae)
5.14. Pseuderanthemum palatiferum (Acanthaceae)
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hrachovinová, I. Diagnostic strategies in disorders of hemostasis. Vnitr. Lek. 2018, 64, 537–544. [Google Scholar] [CrossRef]
- Wolberg, A.S.; Rosendaal, F.R.; Weitz, J.I.; Jaffer, I.H.; Agnelli, G.; Baglin, T.; Mackman, N. Venous thrombosis. Nat. Rev. Dis. Primers 2015, 7, 15006. [Google Scholar] [CrossRef]
- Sadek, M.; Almeida, J.I.; Kabnick, L.S. Endothermal Heat-Induced Thrombosis. In Atlas of Endovascular Venous Surgery, 2nd ed.; Editor Almeida, J.I., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 397–407. [Google Scholar] [CrossRef]
- Saunders, W.B. Venous Thromboembolism. In The Most Common Inpatient Problems in Internal Medicine, 1st ed.; John, C.S., Hylton, V.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 199–221. ISBN 9781416032038. [Google Scholar]
- Vallet, B.; Wiel, E. Endothelial cell dysfunction and coagulation. Crit. Care Med. 2001, 29, 36–41. [Google Scholar] [CrossRef]
- Esmon, C.T. Basic mechanisms and pathogenesis of venous thrombosis. Blood Rev. 2009, 23, 225–229. [Google Scholar] [CrossRef] [Green Version]
- Applegate, J.S.; Gronefeld, D. Factor V Leiden. Radiol. Technol. 2019, 90, 259–273. [Google Scholar]
- Poudel, S.; Zeb, M.; Kondapaneni, V.; Gutlapalli, S.D.; Choudhari, J.; Sodiya, O.T.; Toulassi, I.A.; Cancarevic, I. Association of G20210A Prothrombin Gene Mutation and Cerebral Ischemic Stroke in Young Patients. Cureus 2020, 12, e11984. [Google Scholar] [CrossRef]
- Martinelli, I. Risk factors in venous thromboembolism. Thromb. Haemost. 2001, 86, 395–403. [Google Scholar] [CrossRef]
- Coppola, A.; Davi, G.; De Stefano, V.; Mancini, F.P.; Cerbone, A.M.; Di Minno, G. Homocysteine, coagulation, platelet function, and thrombosis. Semin. Thromb. Hemost. 2000, 26, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Casini, A.; Neerman-Arbez, M.; Ariëns, R.A.; de Moerloose, P. Dysfibrinogenemia: From molecular anomalies to clinical manifestations and management. J. Thromb. Haemost. 2015, 13, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Rabieian, R.; Boshtam, M.; Zareei, M.; Kouhpayeh, S.; Masoudifar, A.; Mirzaei, H. Plasminogen Activator Inhibitor Type-1 as a Regulator of Fibrosis. J. Cell. Biochem. 2018, 119, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Chapin, J.C.; Hajjar, K.A. Fibrinolysis and the control of blood coagulation. Blood Rev. 2015, 29, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, M.; Manwani, D. Thrombotic disorders. Pediatr. Rev. 2011, 32, 41–43. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Lee, A.I. Thrombotic Risk from Chemotherapy and Other Cancer Therapies. Cancer Treat. Res. 2019, 179, 87–101. [Google Scholar] [CrossRef]
- Jame, S.; Barnes, G. Stroke and thromboembolism prevention in atrial fibrillation. Heart 2020, 106, 10–17. [Google Scholar] [CrossRef]
- Burnett, A.E.; Mahan, C.E.; Vazquez, S.R.; Oertel, L.B.; Garcia, D.A.; Ansell, J. Guidance for the practical management of the direct oral anticoagulants (DOACs) in VTE treatment. J. Thromb. Thrombolysis 2016, 41, 206–232. [Google Scholar] [CrossRef] [Green Version]
- Song, A.B.; Rosovsky, R.P.; Connors, J.M.; Al-Samkari, H. Direct oral anticoagulants for treatment and prevention of venous thromboembolism in cancer patients. Vasc. Health Risk Manag. 2019, 21, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Suwanabol, P.A.; Hoch, J.R. Venous thromboembolic disease. Surg. Clin. N. Am. 2013, 93, 983–995. [Google Scholar] [CrossRef]
- Tritschler, T.; Kraaijpoel, N.; Le Gal, G.; Wells, P.S. Venous Thromboembolism: Advances in Diagnosis and Treatment. JAMA 2018, 320, 1583–1594. [Google Scholar] [CrossRef]
- Cohen, A.T.; Hamilton, M.; Mitchell, S.A.; Phatak, H.; Liu, X.; Bird, A.; Tushabe, D.; Batson, S. Comparison of the Novel Oral Anticoagulants Apixaban, Dabigatran, Edoxaban, and Rivaroxaban in the Initial and Long-Term Treatment and Prevention of Venous Thromboembolism: Systematic Review and Network Meta-Analysis. PLoS ONE 2015, 10, e0144856. [Google Scholar] [CrossRef]
- Hass, B.; Pooley, J.; Harrington, A.E.; Clemens, A.; Feuring, M. Treatment of venous thromboembolism—Effects of different therapeutic strategies on bleeding and recurrence rates and considerations for future anticoagulant management. Thromb. J. 2012, 10, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Institute for Health and Care Excellence. CG92: Venous Thromboembolism—Reducing the Risk. 2010. Available online: http://guidance.nice.org.uk/CG92 (accessed on 15 September 2021).
- Guyatt, G.H.; Akl, E.A.; Crowther, M.; Gutterman, D.D.; Schuunemann, H.J. Antithrombotic Therapy and Prevention of Thrombosis, 9th ed: American College of Chest Physicians Evidence-Based Clinical Practice Guidelines. Chest 2012, 141, 7S–47S. [Google Scholar] [CrossRef]
- Lyman, G.H.; Khorana, A.A.; Kuderer, N.M.; Lee, A.Y.; Arcelus, J.I.; Balaban, E.P.; Clarke, J.M.; Flowers, C.R.; Francis, C.W.; Gates, L.E.; et al. Venous thromboembolism prophylaxis and treatment in patients with cancer: American Society of Clinical Oncology clinical practice guideline update. JCO 2013, 31, 2189–2204. [Google Scholar] [CrossRef] [PubMed]
- Ageno, W.; Gallus, A.S.; Wittkowsky, A.; Crowther, M.; Hylek, E.M.; Palareti, G. Oral anticoagulant therapy: Antithrombotic Therapy and Prevention of Thrombosis, 9th ed: American College of Chest Physicians Evidence-Based Clinical Practice Guidelines. Chest 2012, 141, 44S–88S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolaides, A.; Fareed, J.; Kakkar, A.; Breddin, H.; Goldhaber, S.; Hull, R.; Myers, K.; Samama, M.; Fletcher, J.; Kalodiki, E.; et al. Prevention and treatment of venous thromboembolism. International Consensus Statement (guidelines according to scientific evidence). Int. Angiol. 2006, 25, 101–161. [Google Scholar]
- Ay, C.; Beyer-Westendorf, J.; Pabinger, I. Treatment of cancer-associated venous thromboembolism in the age of direct oral anticoagulants. Ann. Oncol. 2019, 30, 897–907. [Google Scholar] [CrossRef]
- Sikorska, J.; Uprichard, J. Direct Oral Anticoagulants: A Quick Guide. Eur. Cardiol. 2017, 12, 40–45. [Google Scholar] [CrossRef]
- Streiff, M.B.; Agnelli, G.; Connors, J.M.; Crowther, M.; Eichinger, S.; Lopes, R.; McBane, R.D.; Moll, S.; Ansell, J. Guidance for the treatment of deep vein thrombosis and pulmonary embolism. J. Thromb. Thrombolysis 2016, 41, 32–67. [Google Scholar] [CrossRef] [Green Version]
- Vilahur, G.; Badimon, L. Antiplatelet properties of natural products. Vascul. Pharmacol. 2013, 59, 67–75. [Google Scholar] [CrossRef]
- Di Minno, A.; Frigerio, B.; Spadarella, G.; Ravani, A.; Sansaro, D.; Amato, M.; Kitzmiller, J.P.; Pepi, M.; Tremoli, E.; Baldassarre, D. Old and new oral anticoagulants: Food, herbal medicines and drug interactions. Blood Rev. 2017, 31, 193–203. [Google Scholar] [CrossRef]
- Seca, A.M.L.; Pinto, D.C.G.A. Plant Secondary Metabolites as Anticancer Agents: Successes in Clinical Trials and Therapeutic Application. Int. J. Mol. Sci. 2018, 19, 263. [Google Scholar] [CrossRef] [Green Version]
- Kuznetsova, T.A.; Andryukov, B.G.; Makarenkova, I.D.; Zaporozhets, T.S.; Besednova, N.N.; Fedyanina, L.N.; Kryzhanovsky, S.P.; Shchelkanov, M.Y. The Potency of Seaweed Sulfated Polysaccharides for the Correction of Hemostasis Disorders in COVID-19. Molecules 2021, 26, 2618. [Google Scholar] [CrossRef]
- Lippi, G.; Favaloro, E.J.; Franchini, M.; Guidi, G.C. Milestones and perspectives in coagulation and hemostasis. Semin. Thromb. Hemost. 2009, 35, 9–22. [Google Scholar] [CrossRef]
- Sepúlveda, C.; Palomo, I.; Fuentes, E. Primary and secondary haemostasis changes related to aging. Mech. Ageing Dev. 2015, 150, 46–54. [Google Scholar] [CrossRef]
- Berndt, M.C.; Metharom, P.; Andrews, R.K. Primary haemostasis: Newer insights. Haemophilia 2014, 20, 15–22. [Google Scholar] [CrossRef]
- Vilahur, G.; Gutiérrez, M.; Arzanauskaite, M.; Mendieta, G.; Ben-Aicha, S.; Badimon, L. Intracellular platelet signalling as a target for drug development. Vascul. Pharmacol. 2018, 111, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Mahaut-Smith, M.P. The unique contribution of ion channels to platelet and megakaryocyte function. J. Thromb. Haemost. 2012, 10, 1722–1732. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Delaney, M.K.; O’Brien, K.A.; Du, X. Signaling during platelet adhesion and activation. Arter. Thromb. Vasc. Biol. 2010, 30, 2341–2349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.; Carrim, N.; Wang, Y.; Gallant, R.C.; Marshall, A.; Ni, H. Platelets in hemostasis and thrombosis: Novel mechanisms of fibrinogen-independent platelet aggregation and fibronectin-mediated protein wave of hemostasis. J. Biomed. Res. 2015, 29, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Reheman, A.; Spring, C.M.; Kalantari, J.; Marshall, A.H.; Wolberg, A.S.; Gross, P.L.; Weitz, J.I.; Rand, M.L.; Mosher, D.F.; et al. Plasma fibronectin supports hemostasis and regulates thrombosis. J. Clin. Investig. 2014, 124, 4281–4293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkhan, P.; Silver, M.J.; Da Costa, P.B.; Tocantins, L.M. Phosphatidyl serine and blood coagulation. Nature 1958, 182, 1031–1032. [Google Scholar] [CrossRef]
- Jackson, S.P. Arterial thrombosis-insidious, unpredictable and deadly. Nat. Med. 2011, 17, 1423–1436. [Google Scholar] [CrossRef]
- Versteeg, H.H.; Heemskerk, J.W.M.; Levi, M.; Reitsma, P.H. New fundamentals in hemostasis. Physiol. Rev. 2013, 93, 327–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisman, T.; Weeterings, C.; de Groot, P.G. Platelet aggregation: Involvement of thrombin and fibrin(ogen). Front. Biosci. 2005, 10, 2504–2517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, F.P., Jr.; Antunes, O.A.; de Alencastro, R.B.; De Simone, S.G. The Na+ binding channel of human coagulation proteases: Novel insights on the structure and allosteric modulation revealed by molecular surface analysis. Biophys. Chem. 2006, 119, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.R.; Zhang, D.; Oswald, B.E.; Carrim, N.; Wang, X.; Hou, Y.; Zhang, Q.; Lavalle, C.; McKeown, T.; Marshall, A.H.; et al. Platelets are versatile cells: New discoveries in hemostasis, thrombosis, immune responses, tumor metastasis and beyond. Crit. Rev. Clin. Lab. Sci. 2016, 53, 409–430. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ni, H. Fibronectin maintains the balance between hemostasis and thrombosis. Cell. Mol. Life Sci. 2016, 73, 3265–3277. [Google Scholar] [CrossRef]
- Ramström, S.; Rånby, M.; Lindahl, T.L. The role of platelets in blood coagulation—Effects of platelet agonists and GPIIb/IIIa inhibitors studied by free oscillation rheometry. Thromb. Res. 2002, 105, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, M.; Monroe, D.M. A cell-based model of hemostasis. Thromb. Haemost. 2001, 85, 958–965. [Google Scholar] [PubMed]
- Riddel, J.P., Jr.; Aouizerat, B.E.; Miaskowski, C.; Lillicrap, D.P. Theories of blood coagulation. J. Pediatr. Oncol. Nurs. 2007, 24, 123–131. [Google Scholar] [CrossRef]
- Smith, S.A.; Travers, R.J.; Morrissey, J.H. How it all starts: Initiation of the clotting cascade. Crit. Rev. Biochem. Mol. Biol. 2015, 50, 326–336. [Google Scholar] [CrossRef] [Green Version]
- Norris, L.A. Blood coagulation. Best Pract. Res. Clin. Obstet. Gynaecol. 2003, 17, 369–383. [Google Scholar] [CrossRef]
- Morrissey, J.H.; Broze, G.J., Jr. Tissue factor and the initiation and regulation (TFPI) of coagulation. In Hemostasis and Thrombosis: Basic Principles and Clinical Practice, 6th ed.; Marder, V.J., Aird, W.C., Bennett, J.S., Schulman, S., White, G.C., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 163–178. [Google Scholar]
- Morrisey, J.H. Tissue factor: An enzyme cofactor and a true receptor. Thromb. Haemost. 2001, 86, 66–74. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, C.S.; Miraglia, C.C.; Rickles, F.R.; Shuman, M.A. Cleavage of blood coagulation factor XIII and fibrinogen by thrombin during in vitro clotting. J. Clin. Investig. 1985, 75, 1463–1470. [Google Scholar] [CrossRef]
- Silverberg, M.; Dunn, J.T.; Garen, L.; Kaplan, A.P. Autoactivation of human Hageman factor. Demonstration utilizing a synthetic substrate. J. Biol. Chem. 1980, 255, 7281–7286. [Google Scholar] [CrossRef]
- Tankersley, D.L.; Finlayson, J.S. Kinetics of activation and autoactivation of human factor XII. Biochemistry 1984, 23, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Boon, G.D. An overview of hemostasis. Toxicol. Pathol. 1993, 21, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Müller, F.; Gailani, D.; Renné, T. Factor XI and XII as antithrombotic targets. Curr. Opin. Hematol. 2011, 18, 349–355. [Google Scholar] [CrossRef] [Green Version]
- Scott, C.F.; Silver, L.D.; Purdon, A.D.; Colman, R.W. Cleavage of human high molecular weight kininogen by factor XIa in vitro. Effect on structure and function. J. Biol. Chem. 1985, 260, 10856–10863. [Google Scholar] [CrossRef]
- Renné, T.; Schmaier, A.H.; Nickel, K.F.; Blombäck, M.; Maas, C. In vivo roles of factor XII. Blood 2012, 120, 4296–4303. [Google Scholar] [CrossRef] [Green Version]
- Kane, W.H. Factor V. In Haemostasis and Thrombosis, Basic Principles and Clinical Practice, 4th ed.; Colman, R.W., Hirsh, J., Marder, V.J., Clowes, A.W., George, J.N., Edwin, W., Salzman, J.B., Eds.; Lippincott/Williams and Wilkins: New York, NY, USA, 2001; pp. 157–169. [Google Scholar]
- Schreuder, M.; Reitsma, P.H.; Bos, M.H.A. Blood coagulation factor Va’s key interactive residues and regions for prothrombinase assembly and prothrombin binding. J. Thromb. Haemost. 2019, 17, 1229–1239. [Google Scholar] [CrossRef] [Green Version]
- Brummel, K.E.; Paradis, S.G.; Butenas, S.; Mann, K.G. Thrombin functions during tissue factor-induced blood coagulation. Blood 2002, 100, 148–152. [Google Scholar] [CrossRef] [Green Version]
- Kalafatis, M.; Egan, J.O.; van’t Veer, C.; Cawthern, K.M.; Mann, K.G. The regulation of clotting factors. Crit. Rev. Eukaryot Gene Expr. 1997, 7, 241–280. [Google Scholar] [CrossRef]
- Preston, F.E.; Greaves, M. Thrombotic vascular disease. Br. J. Hosp. Med. 1985, 34, 46–50. [Google Scholar]
- Koupenova, M.; Kehrel, B.E.; Corkrey, H.A.; Freedman, J.E. Thrombosis and platelets: An update. Eur. Heart J. 2017, 38, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Heit, J.A.; Spencer, F.A.; White, R.H. The epidemiology of venous thromboembolism. J. Thromb. Thrombolysis 2016, 41, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, A.; Fullam, L.; Brownstein, A.P.; Maynard, G.A.; Ansell, J.; Varga, E.A.; Friedman, R.J.; Rickles, F.R. Deep vein thrombosis (DVT) and pulmonary embolism (PE): Awareness and prophylaxis practices reported by patients with cancer. Cancer Investig. 2015, 33, 405–410. [Google Scholar] [CrossRef]
- Goldhaber, S.Z.; Bounameaux, H. Pulmonary embolism and deep vein thrombosis. Lancet 2012, 379, 1835–1846. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.T.; Ikram, A.; Saeed, O.; Afridi, T.; Sila, C.A.; Smith, M.S.; Irshad, K.; Shuaib, A. Deep Vein Thrombosis in Acute Stroke—A Systemic Review of the Literature. Cureus 2017, 9, e1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, S.R. The post-thrombotic syndrome. Hematol. Am. Soc. Hematol. Educ. Program 2016, 1, 413–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberhardt, R.T.; Raffetto, J.D. Chronic venous insufficiency. Circulation 2014, 130, 333–346. [Google Scholar] [CrossRef]
- Di Nisio, M.; van Es, N.; Büller, H.R. Deep vein thrombosis and pulmonary embolism. Lancet 2016, 388, 3060–3073. [Google Scholar] [CrossRef]
- Malaponte, G.; Signorelli, S.S.; Bevelacqua, V.; Polesel, J.; Taborelli, M.; Guarneri, C.; Fenga, C.; Umezawa, K.; Libra, M. Increased Levels of NF-kB-Dependent Markers in Cancer-Associated Deep Venous Thrombosis. PLoS ONE 2015, 10, e0132496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sallah, S.; Wan, J.Y.; Nguyen, N.P. Venous thrombosis in patients with solid tumors: Determination of frequency and characteristics. Thromb. Haemost. 2002, 7, 575–579. [Google Scholar]
- Wun, T.; White, R.H. Venous thromboembolism (VTE) in patients with cancer: Epidemiology and risk factors. Cancer Investig. 2009, 27, 63–74. [Google Scholar] [CrossRef]
- Rickles, F.R.; Falanga, A. Molecular basis for the relationship between thrombosis and cancer. Thromb. Res. 2001, 102, 215–224. [Google Scholar] [CrossRef]
- Rickles, F.R.; Edward, R.L. Activation of blood coagulation in cancer: Trousseau’s syndrome revisited. Blood 1983, 6, 14–17. [Google Scholar] [CrossRef] [Green Version]
- Sorensen, H.T.; Mellemkjaer, L.; Steffensen, F.H.; Olsen, J.H.; Nielsen, G.L. The risk of a diagnosis of cancer after primary deep venous thrombosis or pulmonary embolism. N. Engl. J. Med. 1998, 338, 1169–1173. [Google Scholar] [CrossRef]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, B.B.; Gehlot, P. Inflammation and cancer: How friendly is the relationship for cancer patients? Curr. Opin. Pharmacol. 2009, 9, 351–369. [Google Scholar] [CrossRef] [Green Version]
- Esmon, C.T. Does inflammation contribute to thrombotic events? Haemostasis 2000, 30, S34–S40. [Google Scholar] [CrossRef]
- Szaba, F.M.; Smiley, S.T. Roles for thrombin and fibrinogen in cytokine/chemokine production and macrophage adhesion in vivo. Blood 2002, 99, 1053–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamponi, S. Potential use of plants and their extracts in the treatment of coagulation disorders in COVID-19 disease: A narrative review. LCM 2021, 4, 26. [Google Scholar] [CrossRef]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qiu, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: A descriptive study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Connors, J.M.; Levy, J.H. COVID-19 and its implications for thrombosis and anticoagulation. Blood J. Am. Soc. Hematol. 2020, 135, 2033–2040. [Google Scholar] [CrossRef]
- Lee, S.G.; Fralick, M.; Sholzberg, M. Coagulopathy associated with COVID-19. CMAJ 2020, 192, E583. [Google Scholar] [CrossRef]
- Levi, M.; Thachil, J.; Iba, T.; Levy, J.H. Coagulation abnormalities and thrombosis in patients with COVID-19. Lancet Haematol. 2020, 7, e438–e440. [Google Scholar] [CrossRef]
- Henry, B.M.; de Oliveira, M.H.S.; Benoit, S.; Plebani, M.; Lippi, G. Hematologic, biochemical and immune biomarker abnormalities associated with severe illness and mortality in coronavirus disease 2019 (COVID-19): A meta-analysis. Clin. Chem. Lab. Med. 2020, 58, 1021–1028. [Google Scholar] [CrossRef] [Green Version]
- Debuc, B.; Smadja, D.M. Is COVID-19 a New hematologic disease? Stem. Cell Rev. Rep. 2021, 17, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Ichiyama, M.; Ohga, S.; Ochiai, M.; Tanaka, K.; Matsunaga, Y.; Kusuda, T.; Inoue, H.; Ishimura, M.; Takimoto, T.; Koga, Y.; et al. Age-specific onset and distribution of the natural anticoagulant deficiency in pediatric thromboembolism. Pediatr. Res. 2016, 79, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Franchini, M.; Marano, G.; Cruciani, M.; Mengoli, C.; Pati, I.; Masiello, F.; Veropalumbo, E.; Pupella, S.; Vaglio, S.; Liumbruno, G.M. COVID-19-associated coagulopathy. Diagnosis 2020, 7, 357–363. [Google Scholar] [CrossRef]
- Liu, Z.; Xiao, X.; Wei, X.; Li, J.; Yang, J.; Tan, H.; Zhu, J.; Zhang, Q.; Wu, J.; Liu, L. Composition and divergence of coronavirus spike proteins and host ACE2 receptors predict potential intermediate hosts of SARS-CoV-2. J. Med. Virol. 2020, 92, 595–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leisman, D.E.; Deutschman, C.S.; Legrand, M. Facing COVID-19 in the ICU: Vascular dysfunction, thrombosis, and dysregulated inflammation. Intensive Care Med. 2020, 46, 1105–1108. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Noubouossie, D.F.; Whelihan, M.F.; Yu, Y.B.; Sparkenbaugh, E.; Pawlinski, R.; Monroe, D.M.; Key, N.S. In vitro activation of coagulation by human neutrophil DNA and histone proteins but not neutrophil extracellular traps. Blood 2017, 129, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, H.; Qu, M.; Nan, K.; Cao, H.; Cata, J.P.; Chen, W.; Miao, C. Review: The Emerging Role of Neutrophil Extracellular Traps in Sepsis and Sepsis-Associated Thrombosis. Front. Cell Infect. Microbiol. 2021, 11, 653228. [Google Scholar] [CrossRef]
- Becker, R.C. COVID-19 update: Covid-19-associated coagulopathy. J. Thromb. Thrombolysis 2020, 50, 54–67. [Google Scholar] [CrossRef]
- Tomo, S.; Kumar, K.P.; Roy, D.; Sankanagoudar, S.; Purohit, P.; Yadav, D.; Banerjee, M.; Sharma, P.; Misra, S. Complement activation and coagulopathy—An ominous duo in COVID19. Expert Rev. Hematol. 2021, 14, 155–173. [Google Scholar] [CrossRef]
- Oikonomopoulou, K.; Ricklin, D.; Ward, P.A.; Lambris, J.D. Interactions between coagulation and complement—Their role in inflammation. Semin. Immunopathol. 2011, 34, 151–165. [Google Scholar] [CrossRef] [Green Version]
- McLean, K.C.; James, A.H. Diagnosis and Management of VTE in Pregnancy. Clin. Obstet. Gynecol. 2018, 61, 206–218. [Google Scholar] [CrossRef]
- Nyakudya, T.T.; Tshabalala, T.; Dangarembizi, R.; Erlwanger, K.H.; Ndhlala, A.R. The Potential Therapeutic Value of Medicinal Plants in the Management of Metabolic Disorders. Molecules 2020, 25, 2669. [Google Scholar] [CrossRef]
- Gholipour, S.; Sewell, R.D.E.; Lorigooini, Z.; Rafieian-Kopaei, M. Medicinal Plants and Atherosclerosis: A Review on Molecular Aspects. Curr. Pharm. Des. 2018, 24, 3123–3131. [Google Scholar] [CrossRef]
- Cavalcanti, R.N.; Forster-Carneiro, T.; Gomes, M.T.M.S.; Rostagno, M.A.; Prado, J.M.; Meireles, M.A.A. Uses and Applications of Extracts from Natural Sources. In Natural Product Extraction: Principles and Applications; Green Chemistry Series; Rostagno, M.A., Prado, J.M., Eds.; The Royal Society of Chemistry: London, UK, 2013; pp. 1–57. [Google Scholar]
- Segneanu, A.E.; Velciov, S.M.; Olariu, S.; Cziple, S.; Damian, D.; Grozescu, I. Bioactive Molecules Profile from Natural Compounds. In Amino Acid—New Insights and Roles in Plant and Animal; Asao, T., Ed.; IntechOpen: London, UK, 2017; pp. 209–228. [Google Scholar]
- Bar-Ya’akov, I.; Tian, L.; Amir, R.; Holland, D. Primary Metabolites, Anthocyanins, and Hydrolyzable Tannins in the Pomegranate Fruit. Front. Plant Sci. 2019, 10, 620. [Google Scholar] [CrossRef] [Green Version]
- Erb, M.; Kliebenstein, D.J. Plant Secondary Metabolites as Defenses, Regulators, and Primary Metabolites: The Blurred Functional Trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- Thirumurugan, D.; Cholarajan, A.; Suresh SSVijayakumar, R.; Vijayakumar, R. An Introductory Chapter: Secondary Metabolites. In Secondary Metabolites—Sources and Applications; Asao, T., Ed.; IntechOpen: London, UK, 2018; pp. 1–20. [Google Scholar] [CrossRef] [Green Version]
- Gorlenko, C.L.; Kiselev, H.Y.; Budanova, E.V.; Zamyatnin, A.A., Jr.; Ikryannikova, L.N. Plant Secondary Metabolites in the Battle of Drugs and Drug-Resistant Bacteria: New Heroes or Worse Clones of Antibiotics? Antibiotics 2020, 9, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaynab, M.; Fatima, M.; Abbas, S.; Sharif, Y.; Umair, M.; Zafar, M.H.; Bahadar, K. Role of secondary metabolites in plant defense against pathogens. Microb. Pathog. 2018, 124, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Anand, U.; Jacobo-Herrera, N.; Altemimi, A.; Lakhssassi, N. A comprehensive review on medicinal plants as antimicrobial therapeutics: Potential avenues of biocompatible drug discovery. Metabolites 2019, 9, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, J.J.; El Haouari, M.; Jardin, I.; Alonso, N.; Regodon, S.; Diez-Bello, R.; Redondo, P.C.; Rosado, J.A. Flavonoids and Platelet-Derived Thrombotic Disorders. Curr. Med. Chem. 2019, 26, 7035–7047. [Google Scholar] [CrossRef] [PubMed]
- Marimoutou, M.; Le Sage, F.; Smadja, J.; Lefebvre d’Hellencourt, C.; Gonthier, M.P.; Robert-Da Silva, C. Antioxidant polyphenol-rich extracts from the medicinal plants Antirhea borbonica, Doratoxylon apetalum and Gouania mauritiana protect 3T3-L1 preadipocytes against H2O2, TNFα and LPS inflammatory mediators by regulating the expression of superoxide dismutase and NF-κB genes. J. Inflamm. 2015, 12, 10. [Google Scholar] [CrossRef] [Green Version]
- Bijak, M.; Saluk, J.; Szelenberger, R.; Nowak, P. Popular naturally occurring antioxidants as potential anticoagulant drugs. Chem. Biol. Interact 2016, 257, 35–45. [Google Scholar] [CrossRef]
- Abbas, M.; Saeed, F.; Anjum, F.M.; Afzaal, M.; Tufail, T.; Bashir, M.S.; Ishtiaq, A.; Hussain, S.; Suleria, H.A.R. Natural polyphenols: An overview. Int. J. Food Prop. 2017, 20, 1689–1699. [Google Scholar] [CrossRef] [Green Version]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- El Haouari, M.; Rosado, J.A. Medicinal Plants with Antiplatelet Activity. Phytother. Res. 2016, 30, 1059–1071. [Google Scholar] [CrossRef]
- Ghanbari, R.; Anwar, F.; Alkharfy, K.M.; Gilani, A.H.; Saari, N. Valuable nutrients and functional bioactives in different parts of olive (Olea europaea L.)—A review. Int. J. Mol. Sci. 2012, 13, 3291–3340. [Google Scholar] [CrossRef]
- El, S.N.; Karakaya, S. Olive tree (Olea europaea) leaves: Potential beneficial effects on human health. Nutr. Rev. 2009, 67, 632–638. [Google Scholar] [CrossRef]
- Martínez, L.; Ros, G.; Nieto, G. Hydroxytyrosol: Health Benefits and Use as Functional Ingredient in Meat. Medicines 2018, 5, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa, J.A.; Lopez-Villodres, J.A.; Asensi, R.; Espartero, J.L.; Rodriguez-Gutierez, G.; De La Cruz, J.P. Virgin olive oil polyphenol hydroxytyrosol acetate inhibits in vitro platelet aggregation in human whole blood: Comparison with hydroxytyrosol and acetylsalicylic acid. Br. J. Nutr. 2009, 101, 1157–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio-Senent, F.; de Roos, B.; Duthie, G.; Fernández-Bolaños, J.; Rodríguez-Gutiérrez, G. Inhibitory and synergistic effects of natural olive phenols on human platelet aggregation and lipid peroxidation of microsomes from vitamin E-deficient rats. Eur. J. Nutr. 2015, 54, 1287–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuck, K.L.; Hayball, P.J. Major phenolic compounds in olive oil: Metabolism and health effects. J. Nutr. Biochem. 2002, 13, 636–644. [Google Scholar] [CrossRef]
- Zbidi, H.; Salido, S.; Altarejos, J.; Perez-Bonilla, M.; Bartegi, A.; Rosado, J.A.; Salido, G.M. Olive tree wood phenolic compounds with human platelet antiaggregant properties. Blood Cells Mol. Dis. 2009, 42, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.; Mok, M.; Christensen, A.M.; Turner, A.H.; Hawley, J.A. The effects of polyphenols in olive leaves on platelet function. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Benavente-Garcia, O.; Castillo, J.; Lorente, J.; Ortuño, A.; Del Rio, J.A. Antioxidant activity of phenolics extracted from Olea europaea L. leaves. Food Chem. 2000, 68, 457–462. [Google Scholar] [CrossRef]
- McKay, D.L.; Blumberg, J.B. A review of the bioactivity and potential health benefits of chamomile tea (Matricaria recutita L.). Phytother. Res. 2006, 20, 519–530. [Google Scholar] [CrossRef]
- Pierre, S.; Crosbie, L.; Duttaroy, A.K. Inhibitory effect of aqueous extracts of some herbs on human platelet aggregation in vitro. Platelets 2005, 16, 469–473. [Google Scholar] [CrossRef]
- Bijak, M.; Saluk, J.; Tsirigotis-Maniecka, M.; Komorowska, H.; Wachowicz, B.; Zaczyńska, E.; Czarny, A.; Czechowski, F.; Nowak, P.; Pawlaczyk, I. The influence of conjugates isolated from Matricaria chamomilla L. on platelets activity and cytotoxicity. Int. J. Biol. Macromol. 2013, 61, 218–229. [Google Scholar] [CrossRef]
- Liu, J.; Bai, R.; Liu, Y.; Zhang, X.; Kan, J.; Jin, C. Isolation, structural characterization and bioactivities of naturally occurring polysaccharide-polyphenolic conjugates from medicinal plants-A review. Int. J. Biol. Macromol. 2018, 107, 2242–2250. [Google Scholar] [CrossRef]
- Rana, S.V.; Pal, R.; Vaiphei, K.; Sharma, S.K.; Ola, R.P. Garlic in health and disease. Nutr. Res. Rev. 2011, 24, 60–71. [Google Scholar] [CrossRef] [Green Version]
- Jagroop, I.A. Plant extracts inhibit ADP-induced platelet activation in humans: Their potential therapeutic role as ADP antagonists. Purinergic Signal 2014, 10, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, G.; Trueman, L.; Crowther, T.; Thomas, B.; Smith, B. Onions—A global benefit to health. Phytother. Res. 2002, 16, 603–615. [Google Scholar] [CrossRef]
- Makheja, A.N.; Bailey, J.M. Antiplatelet constituents of garlic and onion. Agents Actions 1990, 29, 360–363. [Google Scholar] [CrossRef] [PubMed]
- Rahman, K.; Lowe, G.M. Garlic and cardiovascular disease: A critical review. J. Nutr. 2006, 136, 736–740. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.S.; Yamato, O.; Yamasaki, M.; Maede, Y. Modulatory influence of sodium 2-propenyl thiosulfate from garlic on cyclooxygenase activity in canine platelets: Possible mechanism for the anti-aggregatory effect. Prostaglandins Leukot. Essent. Fat. Acids 2005, 72, 351–355. [Google Scholar] [CrossRef]
- MacDonald, J.A.; Marchand, M.E.; Langler, R.F. Improving upon the in vitro biological activity of antithrombotic disulfides. Blood Coagul. Fibrinolysis 2004, 15, 447–450. [Google Scholar] [CrossRef]
- Hiyasat, B.; Sabha, D.; Grotzinger, K.; Kempfert, J.; Rauwald, J.W.; Mohr, F.W.; Dhein, S. Antiplatelet activity of Allium ursinum and Allium sativum. Pharmacology 2009, 83, 197–204. [Google Scholar] [CrossRef]
- Briggs, W.H.; Xiao, H.; Parkin, K.L.; Shen, C.; Goldman, I.L. Differential inhibition of human platelet aggregation by selected Allium thiosulfinates. J. Agric. Food Chem. 2000, 48, 5731–5735. [Google Scholar] [CrossRef]
- Beretta, H.V.; Bannoud, F.; Insani, M.; Berli, F.; Hirschegger, P.; Galmarini, C.R.; Cavagnaro, P.F. Relationships Between Bioactive Compound Content and the Antiplatelet and Antioxidant Activities of Six Allium Vegetable Species. Food Technol. Biotechnol. 2017, 55, 266–275. [Google Scholar] [CrossRef]
- Borges, R.S.; Ortiz, B.L.S.; Pereira, A.C.M.; Keita, H.; Tavares Carvalho, J.C. Rosmarinus officinalis essential oil: A review of its phytochemistry, anti-inflammatory activity, and mechanisms of action involved. J. Ethnopharmacol. 2019, 229, 29–45. [Google Scholar] [CrossRef]
- Lamponi, S.; Baratto, M.C.; Miraldi, E.; Baini, G.; Biagi, M. Chemical Profile, Antioxidant, Anti-Proliferative, Anticoagulant and Mutagenic Effects of a Hydroalcoholic Extract of Tuscan Rosmarinus officinalis. Plants 2021, 10, 97. [Google Scholar] [CrossRef]
- Khouya, T.; Ramchoun, M.; Hmidani, A.; Amrani, S.; Harnafi, H.; Benlyas, M.; Zegzouti, Y.F.; Alem, C. Anti-inflammatory, anticoagulant and antioxidant effects of aqueous extracts from Moroccan thyme varieties. Asian Pac. J. Trop. Biomed. 2015, 5, 636–644. [Google Scholar] [CrossRef] [Green Version]
- Altinier, G.; Sosa, S.; Aquino, R.P.; Mencherini, T.; Mencherini, T.; Della Loggia, R.; Tubaro, A. Characterization of topical antiinflammatory compounds in Rosmarinus officinalis L. J. Agric. Food Chem. 2007, 55, 1718–1723. [Google Scholar] [CrossRef] [PubMed]
- Benincá, P.J.; Bastos Dalmarco, J.; Pizzolatti, M.G.; Fröde, T.S. Analysis of the anti-inflammatory properties of Rosmarinus officinalis L. in mice. Food Chem. 2011, 124, 468–475. [Google Scholar] [CrossRef]
- Olas, B.; Żuchowski, J.; Lis, B.; Skalski, B.; Kontek, B.; Grabarczyk, Ł.; Stochmal, A. Comparative chemical composition, antioxidant and anticoagulant properties of phenolic fraction (a rich in non-acylated and acylated flavonoids and non-polar compounds) and non-polar fraction from Elaeagnus rhamnoides (L.) A. Nelson fruits. Food Chem. 2018, 247, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Osunsanmi, F.O.; Zaharare, G.E.; Oyinloye, B.E.; Mosa, R.A.; Ikhile, M.I.; Shode, S.O.; Ogunyinka, I.B.; Opoku, A.R. Antithrombotic, anticoagulant and antiplatelet activity of betulinic acid and 3β-acetoxybetulinic acid from Melaleuca bracteata ‘Revolution Gold’ (Myrtaceae) Muell leaf. Trop. J. Pharm. Res. 2018, 17, 1983–1989. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-J.; Jin, Y.-R.; Lee, J.-H.; Yu, J.-Y.; Han, X.-H.; Oh, K.-W.; Hong, J.T.; Kim, T.-J.; Yun, Y.-P.l. Antiplatelet Activity of Carnosic Acid, a Phenolic Diterpene from Rosmarinus officinalis. Planta Med. 2007, 73, 121–127. [Google Scholar] [CrossRef]
- Saluk-Juszczak, J.; Pawlaczyk, I.; Olas, B.; Kołodziejczyk, J.; Ponczek, M.; Nowak, P.; Tsirigotis-Wołoszczak, M.; Wachowicz, B.; Gancarz, R. The effect of polyphenolic-polysaccharide conjugates from selected medicinal plants of Asteraceae family on the peroxynitrite-induced changes in blood platelet proteins. Int. J. Biol. Macromol. 2010, 47, 700–705. [Google Scholar] [CrossRef]
- Pawlaczyk, I.; Czerchawski, L.; Pilecki, W.; Lamer-Zarawskac, E.; Gancarza, R. Polyphenolic-polysaccharide compounds from selected medicinal plants of Asteraceae and Rosaceae families: Chemical characterization and blood anticoagulant activity. Carbohydr. Polym. 2009, 77, 568e575. [Google Scholar] [CrossRef]
- Pawlaczyk, I.; Czerchawski, L.; Kuliczkowski, W.; Karolko, B.; Pilecki, W.; Witkiewicz, W.; Gancarz, R. Anticoagulant and anti-platelet activity of polyphenolic-polysaccharide preparation isolated from the medicinal plant Erigeron canadensis L. Thromb. Res. 2011, 127, 328–340. [Google Scholar] [CrossRef]
- Nabavi, S.M.; Marchese, A.; Izadi, M.; Curti, V.; Daglia, M.; Nabavi, S.F. Plants belonging to the genus Thymus as antibacterial agents: From farm to pharmacy. Food Chem. 2015, 15, 339–347. [Google Scholar] [CrossRef]
- Bellakhdar, J.; Claisse, R.; Fleurentin, J.; Younos, C. Repertory of standard herbal drugs in the Moroccan pharmacopoea. J. Ethnopharmacol. 1991, 35, 123–143. [Google Scholar] [CrossRef]
- Hmidani, A.; Tariq Bouhlali, E.D.; Khouya, T.; Ramchounbd, M.; Filali-Zegzoutia, Y.; Benlyasa, M.; Alemb, C. Effect of extraction methods on antioxidant and anticoagulant activities of Thymus atlanticus aerial part. Sci. Afr. 2019, 5, e00143. [Google Scholar] [CrossRef]
- Hmidani, A.; Tariq Bouhlali, E.D.; Khouya, T.; Ramchounbd, M.; Filali-Zegzoutia, Y.; Alemb, C.; Benlyasa, M. Antioxidant, anti-inflammatory and anticoagulant activities of three Thymus species grown in southeastern Morocco. FJPS 2019, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Khouya, T.; Ramchoun, M.; Hmidani, A.; Tariq Bouhlali, E.D.; Amrani, S.; Alem, C. Phytochemical analysis and bioactivity evaluation of Moroccan Thymus atlanticus (Ball) fractions. Sci. Afr. 2021, 11, e00716. [Google Scholar] [CrossRef]
- Nam, S.; Park, H.-J.; Nam, K.-S. The antithrombotic effect of caffeic acid is associated with a cAMP-dependent pathway and clot retraction in human platelets. Thromb. Res. 2020, 195, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J 2017, 25, 149–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.H.; Kim, K.J.; Kim, S. Comparative Effect of Quercetin and Quercetin-3-O-β-d-Glucoside on Fibrin Polymers, Blood Clots, and in Rodent Models. J. Biochem. Mol. Toxicol. 2016, 30, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Bijak, M.; Ponczek, M.B.; Nowak, P. Polyphenol compounds belonging to flavonoids inhibit activity of coagulation factor X. Int. J. Biol. Macromol. 2014, 65, 129–135. [Google Scholar] [CrossRef]
- Duarte da Luz, J.R.; Silva do Nascimento, T.E.; Araujo-Silva, G.; de Rezende, A.A.; BrandãoNeto, J.; Ururahy, M.A.G.; Luchessia, A.D.; Lópeze, J.A.; Rocha, H.A.O.; Almeida, M.d.G. Licania rigida Benth leaf extracts: Assessment of toxicity and potential anticoagulant effect. S. Afr. J. Bot. 2021, 139, 217–225. [Google Scholar] [CrossRef]
- Parra Pessoa, I.; Lopes Neto, J.J.; Silva de Almeida, T.; Felipe Farias, D.; Vieira, L.R.; Lima de Medeiros, J.; Augusti Boligon, A.; Peijnenburg, A.; Castelar, I.; Fontenele Urano Carvalho, A. Polyphenol Composition, Antioxidant Activity and Cytotoxicity of Seeds from Two Underexploited Wild Licania Species: L. rigida and L. tomentosa. Molecules 2016, 21, 1755. [Google Scholar] [CrossRef] [Green Version]
- Close, R.V.; Giglio, A.M.; Patti, A.F.G.; Lippi, G.; Rizzo, M.; Toth, P.P.; Banach, M. Polyphenols: Potential use in the prevention and treatment of cardiovascular diseases. Curr. Pharm. Des. 2018, 24, 239–258. [Google Scholar] [CrossRef]
- Edziri, H.; Guerrab, M.; Anthonissen, R.; Maha, M.; Verschaeve, L. Phytochemical screening, antioxidant, anticoagulant and in vitro toxic and genotoxic properties of aerial parts extracts of Fumaria officinalis L. growing in Tunisia. S. Afr. J. Bot. 2020, 130, 268–273. [Google Scholar] [CrossRef]
- Satish, K.; Vrushabendra, S.; Kamal, K.G.; Mohan, B. Review on Careya arborea Roxb. Int. J. Res. Ayurveda Pharm. 2010, 1, 306–315. [Google Scholar]
- Gupta, P.C.; Sharma, N.; Rao, C.V. Pharmacognostic studies of the leaves and stem of Careya arborea Roxb. Asian Pac. J. Trop. Biomed. 2012, 2, 404–408. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, M.; Genda, T.; Suehira, M.; Satoh, H.; Miki, A.; Hori, S.; Sawada, Y. Increased anticoagulant activity of warfarin used in combination with doxifluridine. Cancer Chemother. Pharmacol. 2010, 66, 969–972. [Google Scholar] [CrossRef]
- Wadje Shailaja, D.; Wankhede Balaji, G.; Kalambkar Mahesh, R. Identification of bioactive compounds and cytotoxic activity of Careya arborea Roxb. Leaves. J. Pharmacogn. Phytochem. 2019, 2, 362–365. [Google Scholar]
- Ariyaratna, R.A.Y.K.; Amarasinghe, N.R.; Gunawardena, D. Antioxidant phenolic constituents from the fruits of Careya arborea. In Peradeniya University Research Sessions Purse 2007, Part I Agricultural, Biological and Medical Sciences; Editorial Board; University of Peradeniya: Peradeniya, Shri Lanka, 2007; Volume 128, p. 103. [Google Scholar]
- Wang, K.L.C.; Colgrave, M.L.; Gustafson, K.R.; Ireland, D.C.; Goransson, U.; Craik, D.J. Anti-HIV cyclotides from the Chinese medicinal herb Viola yedoensis. J. Nat. Prod. 2008, 71, 47–52. [Google Scholar] [CrossRef]
- Zhou, H.Y.; Hong, J.L.; Shu, P.; Ni, Y.J.; Qin, M.J. A new dicoumarin and anticoagulant activity from Viola yedoensis Makino. Fitoterapia 2009, 80, 283–285. [Google Scholar] [CrossRef]
- Shivaprasad, H.V.; Riyaz, M.; Kumar, R.V.; Dharmappa, K.K.; Tarannum, S.; Siddesha, J.M.; Rajesh, R.; Vishwanath, B.S. Cysteine proteases from the Asclepiadaceae plants latex exhibited thrombin and plasmin like activities. J. Thromb. Thrombolysis 2009, 28, 304–308. [Google Scholar] [CrossRef]
- Siritapetawee, J.; Khunkaewla, P.; Thumanu, K. Roles of a protease from Euphorbia resinifera latex in human anticoagulant and antithrombotic activities. Chem. Biol. Interact. 2020, 329, 109223. [Google Scholar] [CrossRef]
- Skalski, B.; Pawelec, P.; Jedrejek, D.; Rolnik, A.; Pietukhov, R.; Piwowarczyk, R.; Stochmal, A.; Olas, B. Antioxidant and anticoagulant effects of phenylpropanoid glycosides isolated from broomrapes (Orobanche caryophyllacea, Phelipanche arenaria, and P. ramosa). Biomed. Pharmacother. 2021, 139, 111618. [Google Scholar] [CrossRef]
- Dickson, L.; Tenon, M.; Svilar, L.; Fança-Berthon, P.; Martin, J.C.; Rogez, H.; Vaillant, F. Genipap (Genipa americana L.) juice intake biomarkers after medium-term consumption. Food Res. Int. 2020, 137, 109375. [Google Scholar] [CrossRef] [PubMed]
- Madeira, J.C.; da Silva, G.V.L.; Batista, J.J.; Saraiva, G.D.; Santos, G.R.C.; Assreuy, A.M.S.; Mourão, P.A.S.; Pereira, M.G. An arabinogalactan-glycoconjugate from Genipa americana leaves present anticoagulant, antiplatelet and antithrombotic effects. Carbohydr. Polym. 2018, 202, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Madeira, J.C.; Farias, L.A.S.; Luz, C.P.; Assreuy, A.M.S.; Pereira, M.G. Per oral rat treatment with glyconjugate fractions of Genipa americana leaves protects thrombus formation. Blood Coagul. Fibrinolysis 2020, 31, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Padee, P.; Nualkaew, S.; Talubmook, C.; Sakuljaitrong, S. Hypoglycemic effect of a leaf extract of Pseuderanthemum palatiferum (Nees) Radlk. in normal and streptozotocin-induced diabetic rats. J. Ethnopharmacol. 2010, 132, 491–496. [Google Scholar] [CrossRef]
- Ho, T.C.; Kiddane, A.T.; Sivagnanam, S.P.; Park, J.S.; Cho, Y.J.; Getachew, A.T.; Nguyen, T.T.; Kim, G.D.; Chun, B.S. Green extraction of polyphenolic-polysaccharide conjugates from Pseuderanthemum palatiferum (Nees) Radlk.: Chemical profile and anticoagulant activity. Int. J. Biol. Macromol. 2020, 157, 484–493. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Z.; Su, F.; Li, J.; Boadi, E.O.; Chang, Y.X.; Wang, H. Study Structure Activity Relationship of Natural Flavonoids against Thrombin by Molecular Docking Virtual Screening Combined with Activity Evaluation In Vitro. Molecules 2020, 25, 422. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Family | Plants | Classes of Metabolites in the Extracts | Antiplatelet/Anticoagulant Activity of the Extracts |
|---|---|---|---|
| Oleaceae | Olea europaea | oleuropein, (+)-cycloolivil, hydroxytyrosol, hydroxytyrosol acetate, | in vitro inhibition of platelet aggregation and activation [120,121,122,123,124,125,126,127,128,129] |
| Asteracee | Chamomilla recutita L. | flavonoids apigenin, quercetin, patuletin, luteolin and their glucosides, terpenoids, alphabisabolol and its oxides, azulene, including chamazulene | in vitro inhibition of platelet aggregation [130,131,132,133] |
| Amaryllidaceae | Allium sativum, Allium ursinum | sulfur compounds (alliin, allicin, ajoene, allyl propyl disulfide, diallyl trisulfide (DATS), S-allylcysteine (SAC), vinyldithiins, S-allylmercaptocysteine), enzymes (al-liinase, peroxidases, tyrosinase), amino acids (arginine and others) and their glycosides, minerals | in vitro inhibition of platelet aggregation [134,135,136,137,138,139,140,141,142,143] |
| Lamiaceae | Rosmarinus officinalis, Thymus atlanticus, Thymus zygis | tritepenes, ursolic acid, oleanolic acid, betulinic acid, carnosol, micromeric acid, caffeic acid, rosmarinic acid, quercetin, rutin, hyperoside, luteolin-7-O-glucoside | in vitro prolongation of TT, in vivo inhibition of platelet aggregation [144,145,146,147,148,149,150,151], in vitro prolongation of APTT and PT [155,156,157,158,159,160,161,162,163] |
| Rosaceae and Asteraceae | Fragaria vesca, Echinacea purpurea, Erigeron canadensis L. | hexuronic acids and phenolic glycoconjugates | in vitro prolongation of APTT and PT [152,153,154] |
| Chrysobalanaceae | Licania rigida | gallic acid, catechin, chlorogenic acid, caffeic acid, epicatechin, ellagic acid, rutin, quercitrin, quercetin, kaempferol and kaempferol glycoside | in vitro prolongation of APTT and PT; anti-Xa and anti-IIa activity [164,165,166] |
| Papaveracee | Fumaria officinalis | phenolics and flavonoids | in vitro prolongation of APTT and PT [165,166] |
| Lecythidaceae | Careya arborea | 3,4-dihydroxybenzoic acid, quercetin 3-O-glucopyranoside, kaempferol 3-Oglucopyranoside, quercetin 3-O-(6-O-glucopyranosyl)-gluco pyranoside, gallic acid, | in vitro prolongation of APTT, PT and TT [168,169,170,171,172] |
| Violaceae | Viola yedoensis | dimeresculetin | in vitro prolongation of APTT, PT and TT [173,174] |
| Euforbiacee | Euphorbia resinifera | serine protease EuRP-61 | in vitro prolongation of APTT and PT and inhibition of platelet aggregation [175,176] |
| Orobanchaceae | Orobanche caryophyllacea, Phelipanche arenaria, Phelipanche ramosa | phenylpropanoid glycosides: tubuloside A, poliumoside, 3-O-methylpoliumoside | in vitro prolongation of APTT, PT and TT [177] |
| Rubiaceae | Genipa americana | glycoconjugates composed mainly by arabinose, galactose and uronic acid | in vitro prolongation of APTT and inhibition platelets aggregation; in vivo inhibition of venous thrombus formation and increasing of bleeding time [178,179] |
| Acanthaceae | Pseuderanthemum palatiferum | olyphenolic–polysaccharide conjugates | in vitro prolongation of APTT and PT [181,182] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamponi, S. Bioactive Natural Compounds with Antiplatelet and Anticoagulant Activity and Their Potential Role in the Treatment of Thrombotic Disorders. Life 2021, 11, 1095. https://doi.org/10.3390/life11101095
Lamponi S. Bioactive Natural Compounds with Antiplatelet and Anticoagulant Activity and Their Potential Role in the Treatment of Thrombotic Disorders. Life. 2021; 11(10):1095. https://doi.org/10.3390/life11101095
Chicago/Turabian StyleLamponi, Stefania. 2021. "Bioactive Natural Compounds with Antiplatelet and Anticoagulant Activity and Their Potential Role in the Treatment of Thrombotic Disorders" Life 11, no. 10: 1095. https://doi.org/10.3390/life11101095