
Gomisin L1, a Lignan Isolated from Schisandra Berries, Induces Apoptosis by Regulating NADPH Oxidase in Human Ovarian Cancer Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture and MTT Assay
2.3. PI Staining for Cell Cycle Analysis
2.4. PI and Annexin V Double Staining
2.5. Determination of Intracellular Reactive Oxygen Species (ROS)
2.6. RNA Interference for Gene Knockdown
2.7. Statistical Analysis
3. Results
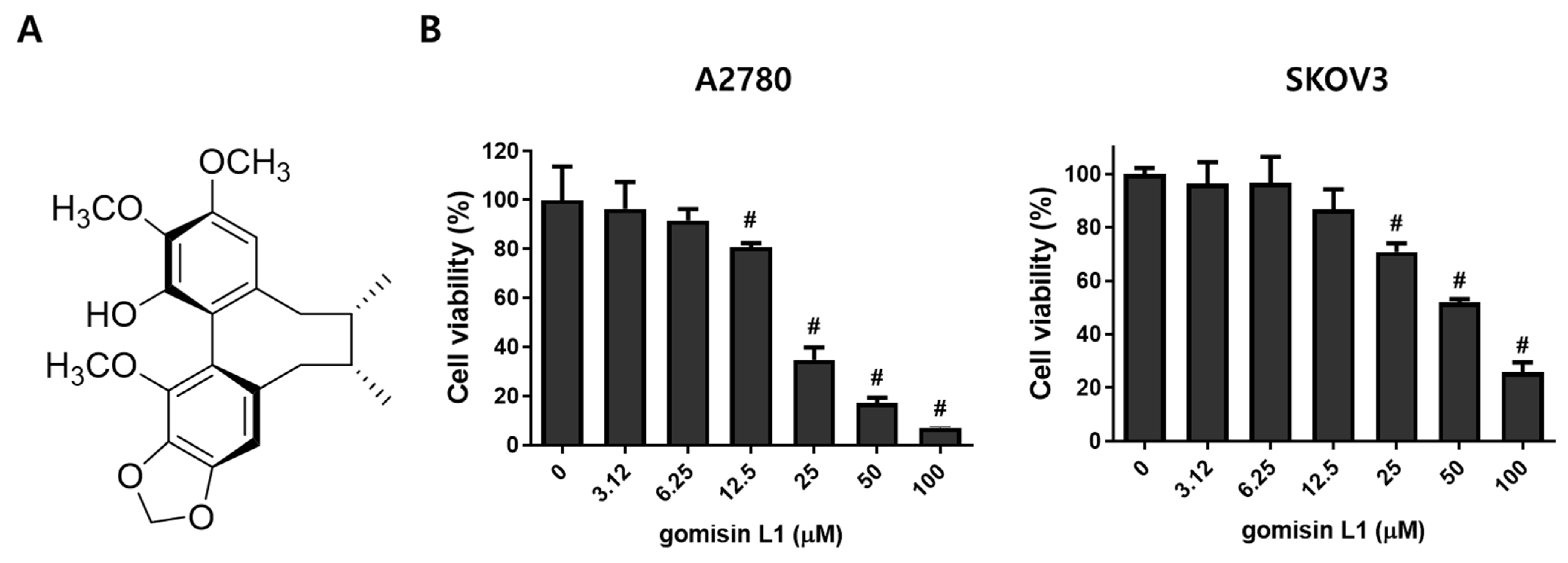
3.1. Gomisin L1 from Schisandra Berries Inhibits the Growth of Human Ovarian Cancer Cells
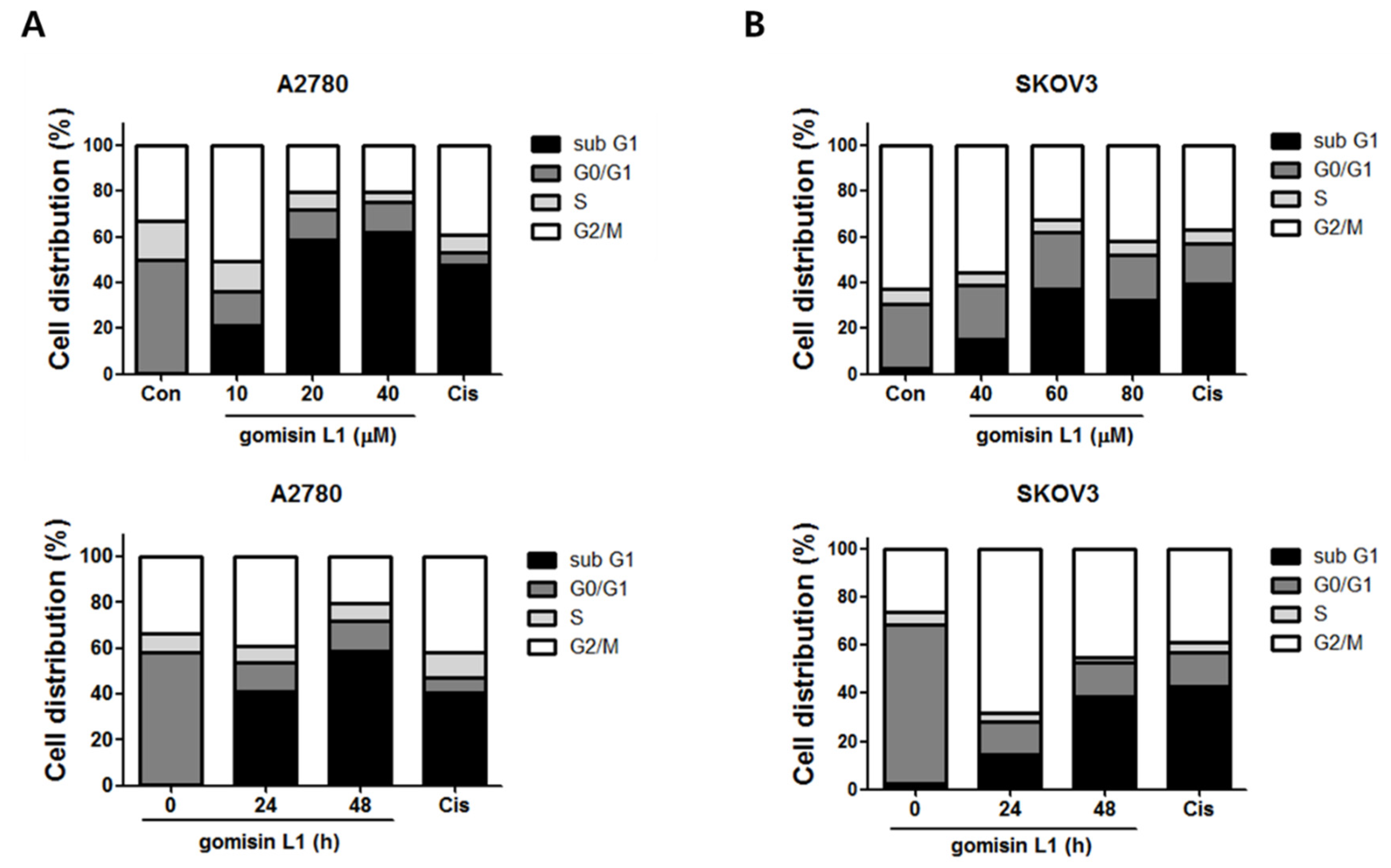
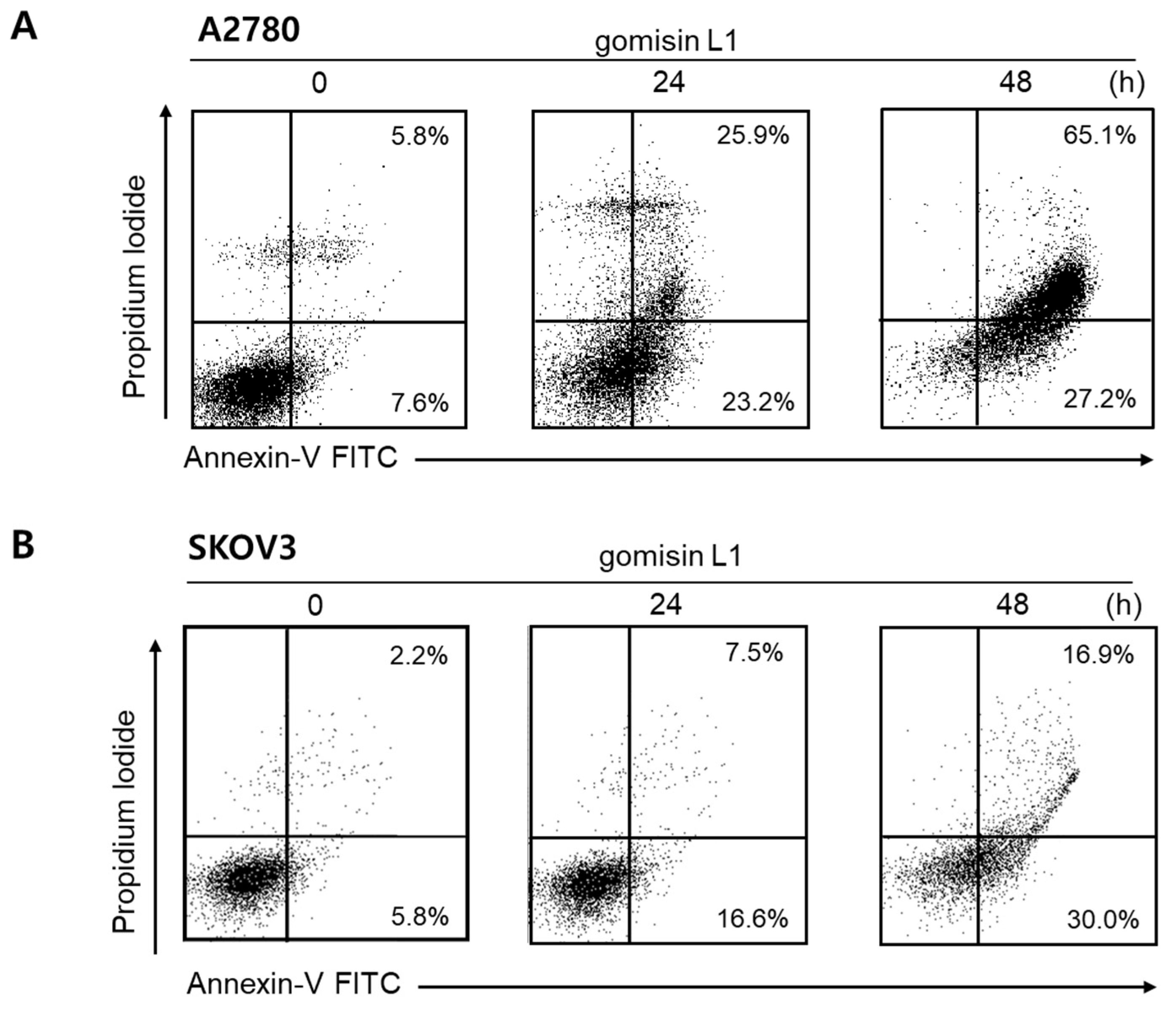
3.2. Gomisin L1 Induces Apoptotic Death of Human Ovarian Cancer Cells
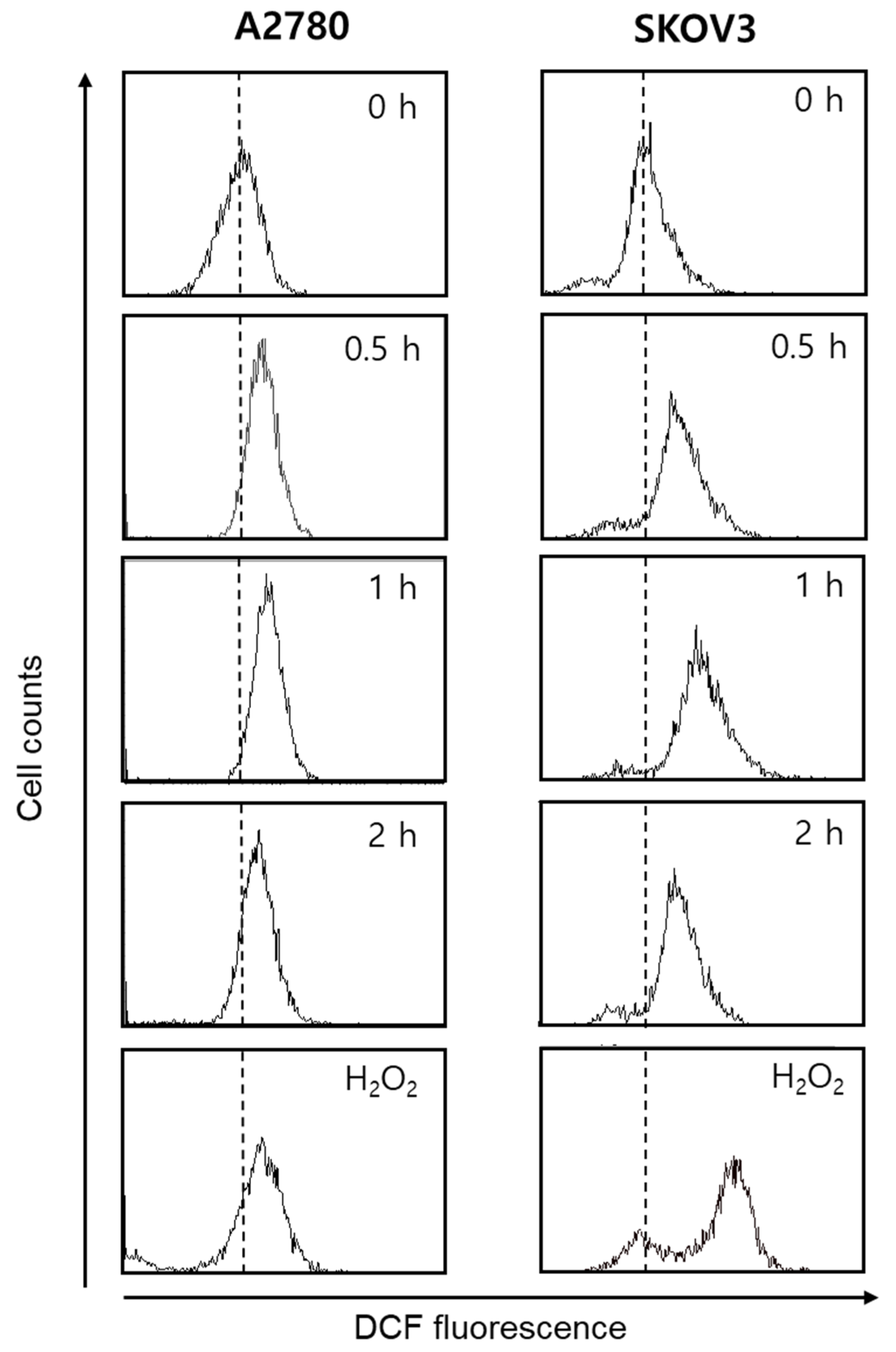
3.3. Gomisin L1-Induced Apoptosis Is Associated with Intracellular Reactive Oxygen Species (ROS) Production
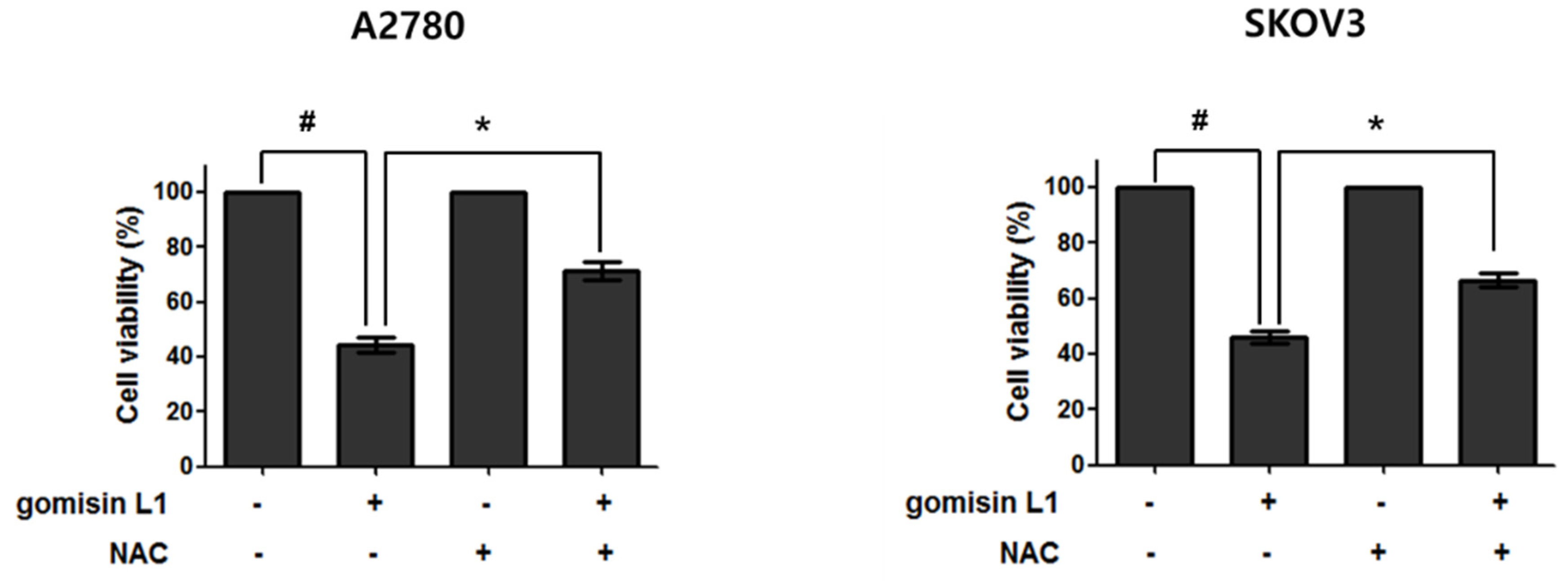
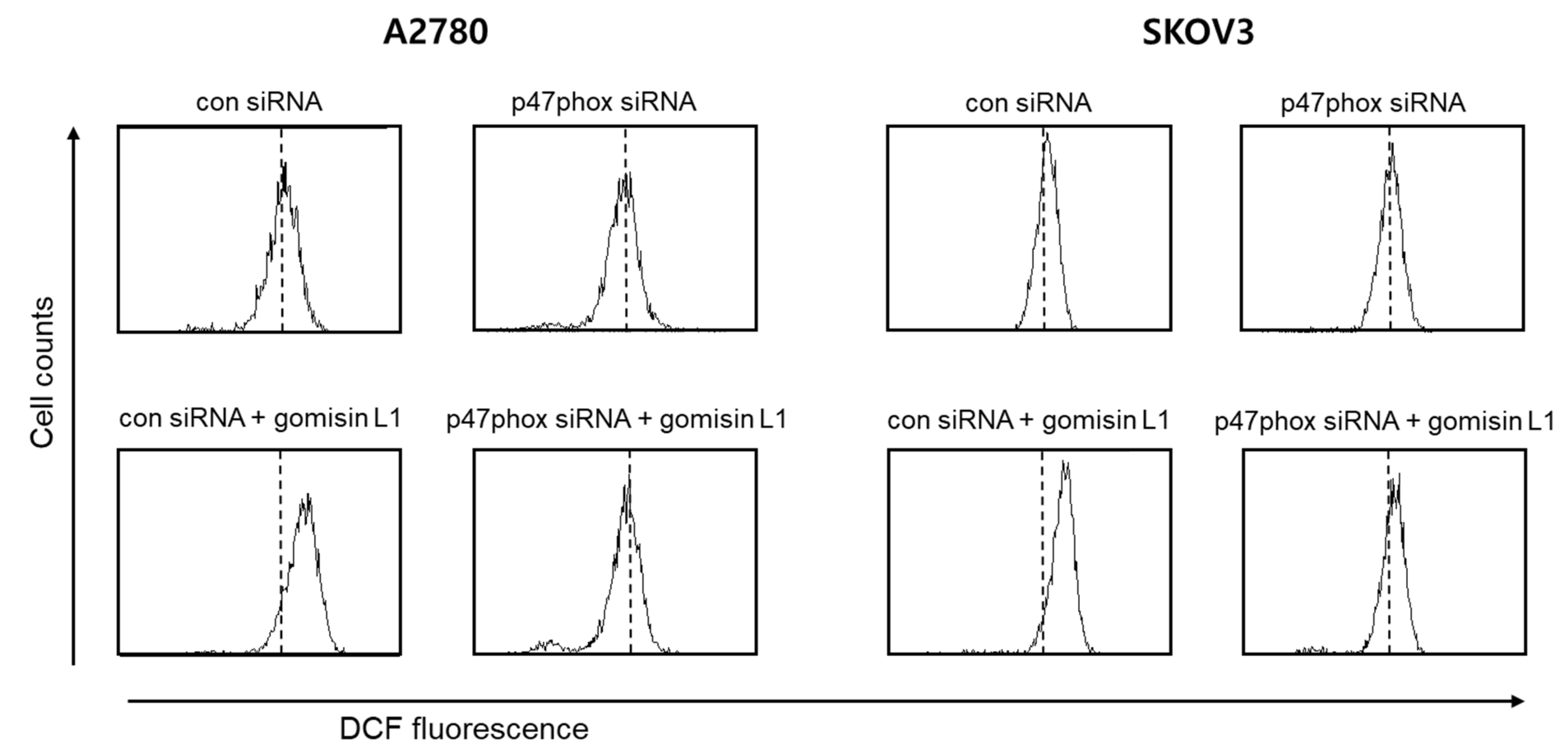
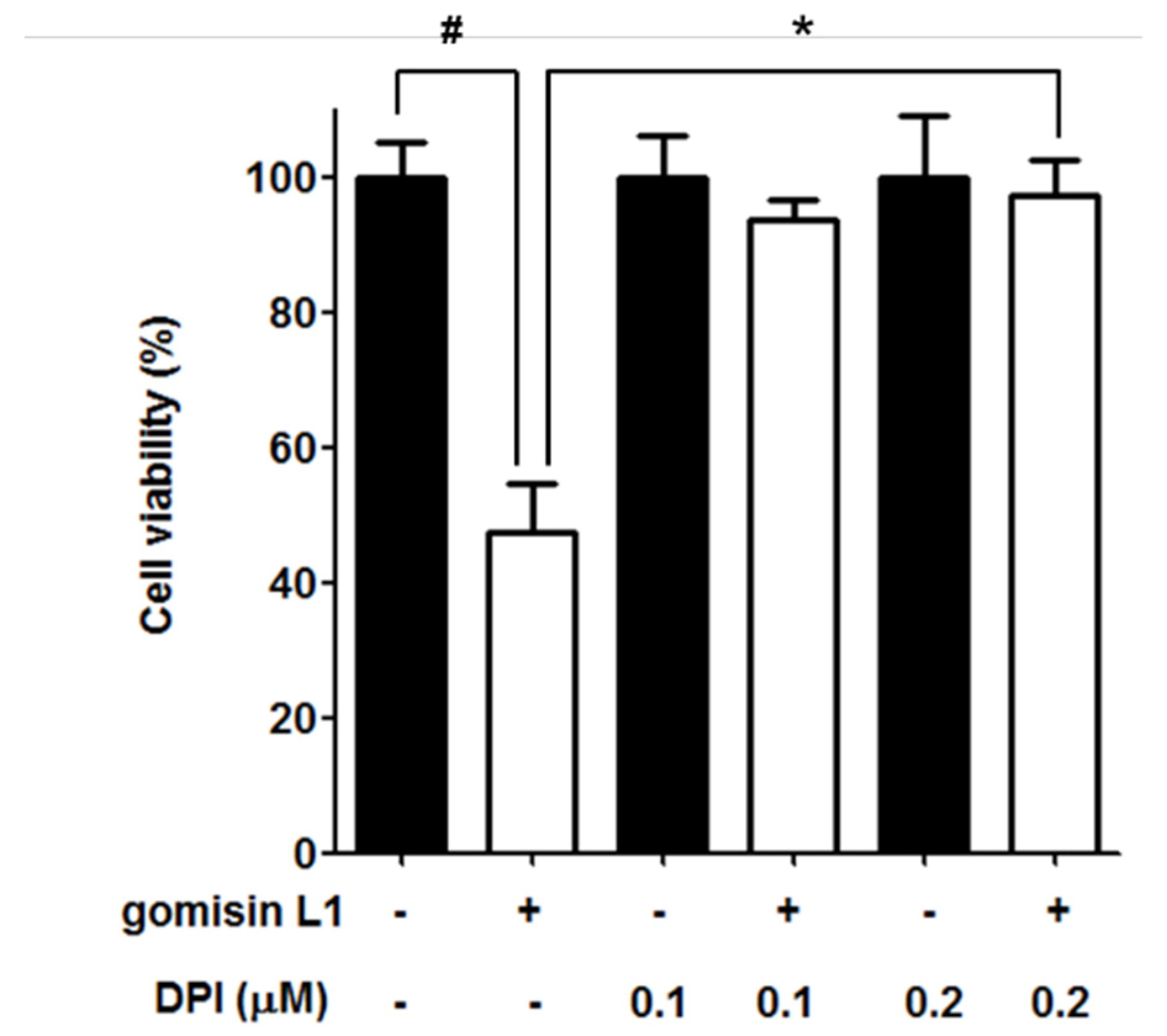
3.4. NADPH Oxidase Is Involved in Gomisin L1-Induced Cell Death
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Hancke, J.; Burgos, R.; Ahumada, F. Schisandra chinensis (Turcz.) Baill. Fitoterapia 1999, 70, 451–471. [Google Scholar] [CrossRef]
- Bensky, D.; Gamble, A. Chinese Herbal Medicine: Materia Medica; Eastland Press: Seattle, WA, USA, 1993. [Google Scholar]
- Lu, Y.; Chen, D.-F. Analysis of Schisandra chinensis and Schisandra sphenanthera. J. Chromatogr. A 2009, 1216, 1980–1990. [Google Scholar] [CrossRef] [PubMed]
- Panossian, A.; Wikman, G. Pharmacology of Schisandra chinensis Bail.: An overview of Russian research and uses in medicine. J. Ethnopharmacol. 2008, 118, 183–212. [Google Scholar] [CrossRef]
- Xue, Y.; Zhang, Y.-L.; Yang, J.-H.; Du, X.; Pu, J.-X.; Zhao, W.; Li, X.-N.; Xiao, W.-L.; Sun, H.-D. Nortriterpenoids and lignans from the fruit of Schisandra chinensis. Chem. Pharm. Bull. 2010, 58, 1606–1611. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, K.; Taguchi, H.; Ikeya, Y.; Endo, T.; Yosioka, I. The Constituents of Schizandra chinensis Baill. XIII. Quantitative analysis of lignans in the fruits of schizandra chinensis Baill. By high performance liquid chromatography. Yakugaku Zasshi 1983, 103, 743–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, H.-Y.; Park, E.-J.; Hong, J.-Y.; Kang, Y.-J.; Kim, S.-J.; Chung, H.-J.; Woo, E.-R.; Hung, T.M.; Youn, U.J.; Kim, Y.S.; et al. Antiproliferative effects of dibenzocyclooctadiene lignans isolated from Schisandra chinensis in human cancer cells. Bioorg. Med. Chem. Lett. 2008, 18, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.J.; Han, N.R.; Kim, K.Y.; Choi, I.S.; Kim, H.M. Gomisin a decreases the lps-induced expression of inos and cox-2 and activation of rip2/nf-kappab in mouse peritoneal macrophages. Immunopharmacol. Immunotoxicol. 2014, 36, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Teng, H.; Lee, W.Y. Antibacterial and antioxidant activities and chemical compositions of volatile oils extracted from Schisandra chinensis Baill. Seeds using simultaneous distillation extraction method, and comparison with Soxhlet and microwave-assisted extraction. Biosci. Biotechnol. Biochem. 2014, 78, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Waiwut, P.; Shin, M.-S.; Yokoyama, S.; Saiki, I.; Sakurai, H. Gomisin a enhances tumor necrosis factor-α-induced G1 cell cycle arrest via signal transducer and activator of transcription 1-mediated phosphorylation of retinoblastoma protein. Biol. Pharm. Bull. 2012, 35, 1997–2003. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Liu, S.; Wu, M.; Wang, F. Research progress in prognostic factors and biomarkers of ovarian cancer. J. Cancer 2021, 12, 3976–3996. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.; Kim, H.M.; Choi, J.-H.; Jang, D.S. Kudsuphilactone B, a nortriterpenoid isolated from Schisandra chinensis fruit, induces caspase-dependent apoptosis in human ovarian cancer A2780 cells. Arch. Pharmacal Res. 2017, 40, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Ikeya, Y.; Taguchi, H.; Yosioka, I. The constituents of schizandra chinensis Baill. X. The structures of gamma-schizandrin and four new lignans,(-)-gomisins L1 and L2,(±)-gomisin M1 and (+)-gomisin M2. Chem. Pharm. Bull. 1982, 30, 132–139. [Google Scholar] [CrossRef] [Green Version]
- Buttke, T.M.; Sandstrom, P.A. Oxidative stress as a mediator of apoptosis. Immunol. Today 1994, 15, 7–10. [Google Scholar] [CrossRef]
- Davis, W.; Ronai, Z.; Tew, K.D. Cellular thiols and reactive oxygen species in drug-induced apoptosis. J. Pharmacol. Exp. Ther. 2001, 296, 1–6. [Google Scholar] [PubMed]
- Hampton, M.B.; Orrenius, S. Redox regulation of apoptotic cell death. BioFactors 1998, 8, 1–5. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial ROS-induced ROS release: An update and review. Biochim. Biophys. Acta (BBA) Bioenerg. 2006, 1757, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Chan, E.C.; Jiang, F.; Peshavariya, H.M.; Dusting, G.J. Regulation of cell proliferation by NADPH oxidase-mediated signaling: Potential roles in tissue repair, regenerative medicine and tissue engineering. Pharmacol. Ther. 2009, 122, 97–108. [Google Scholar] [CrossRef]
- Jiang, F.; Zhang, Y.; Dusting, G.J. NADPH oxidase-mediated redox signaling: Roles in cellular stress response, stress tolerance, and tissue repair. Pharmacol. Rev. 2011, 63, 218–242. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, T.J.; Klaes, M.; Sendker, J. Lignans in seeds of linum species. Phytochemistry 2012, 82, 89–99. [Google Scholar] [CrossRef]
- Kim, H.W.; Shin, J.H.; Lee, M.K.; Jang, G.H.; Lee, S.H.; Jang, H.H.; Jeong, S.T.; Kim, J.B.; Kim, H.U.; Lee, M.G.; et al. Qualitative and quantitative analysis of dibenzocyclooctadiene lignans for the fruits of korean “omija” (Schisandra chinensis). Korean J. Med. Crop. Sci. 2015, 23, 385–394. [Google Scholar] [CrossRef]
- Herchi, W.; Arraez-Roman, D.; Trabelsi, H.; Bouali, I.; Boukhchina, S.; Kallel, H.; Carretero, A.S.; Gutierrez, A.F. Phenolic compounds in flaxseed: A review of their properties and analytical methods. An overview of the last decade. J. Oleo Sci. 2014, 63, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.S.; Lee, J.H.; Park, H.S.; Lee, G.S.; Kim, H.W.; Ha, K.T.; Kim, B.J. Schizandra chinensis extracts induce apoptosis in human gastric cancer cells via jnk/p38 mapk activation and the ros-mediated/mitochondria-dependent pathway. Pharm. Biol. 2015, 53, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.; Shin, S.Y.; Lee, Y.; Hyun, J.; Yong, Y.; Park, J.C.; Lee, Y.H.; Lim, Y. A compound isolated from Schisandra chinensis induces apoptosis. Bioorg. Med. Chem. Lett. 2011, 21, 6054–6057. [Google Scholar] [CrossRef]
- Kim, S.-J.; Min, H.-Y.; Lee, E.J.; Kim, Y.S.; Bae, K.; Kang, S.S.; Lee, S.K. Growth inhibition and cell cycle arrest in the G0/G1 by schizandrin, a dibenzocyclooctadiene lignan isolated from Schisandra chinensis, on T47D human breast cancer cells. Phytother. Res. 2009, 24, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, H.; Inoue, H.; Waiwut, P.; Saiki, I.; Shimada, Y. Gomisin N enhances TRAIL-induced apoptosis via reactive oxygen species-mediated up-regulation of death receptors 4 and 5. Int. J. Oncol. 2011, 40, 1058–1065. [Google Scholar] [CrossRef]
- Huang, M.; Jin, J.; Sun, H.; Liu, G.T. Reversal of P-glycoprotein-mediated multidrug resistance of cancer cells by five schizandrins isolated from the Chinese herb Fructus Schizandrae. Cancer Chemother. Pharmacol. 2008, 62, 1015–1026. [Google Scholar] [CrossRef] [PubMed]
- Fong, W.-F.; Wan, C.-K.; Zhu, G.-Y.; Chattopadhyay, A.; Dey, S.; Zhao, Z.; Shen, X.-L. Schisandrol A from Schisandra chinensis reverses P-glycoprotein-mediated multidrug resistance by affecting pgp-substrate complexes. Planta Med. 2007, 73, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Suh, W.S.; Park, S.Y.; Min, B.S.; Kim, S.H.; Song, J.H.; Shim, S.H. The antiproliferative effects of compounds isolated from schisandra chinensis. Korean J. Food Sci. Technol. 2014, 46, 665–670. [Google Scholar] [CrossRef] [Green Version]
- Ashkenazi, A.; Dixit, V.M. Death receptors: Signaling and modulation. Science 1998, 281, 1305–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamata, H.; Hirata, H. Redox regulation of cellular signalling. Cell. Signal. 1999, 11, 1–14. [Google Scholar] [CrossRef]
- Inoue, M.; Sakaguchi, N.; Isuzugawa, K.; Tani, H.; Ogihara, Y. Role of reactive oxygen species in gallic acid-induced apoptosis. Biol. Pharm. Bull. 2000, 23, 1153–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simamura, E.; Hirai, K.-I.; Shimada, H.; Koyama, J.; Niwa, Y.; Shimizu, S. Furanonaphthoquinones cause apoptosis of cancer cells by inducing the production of reactive oxygen species by the mitochondrial voltage-dependent anion channel. Cancer Biol. Ther. 2006, 5, 1523–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-L.; Fang, L.-W.; Lu, S.-P.; Chou, C.-K.; Luh, T.-Y.; Lai, M.-Z. DNA-damaging reagents induce apoptosis through reactive oxygen species-dependent Fas aggregation. Oncogene 2003, 22, 8168–8177. [Google Scholar] [CrossRef] [Green Version]
- Gotoh, Y.; Cooper, J.A. Reactive oxygen species- and dimerization-induced activation of apoptosis signal-regulating kinase 1 in tumor necrosis factor-α signal transduction. J. Biol. Chem. 1998, 273, 17477–17482. [Google Scholar] [CrossRef] [Green Version]
- Casares, C.; Ramírez-Camacho, R.; Trinidad, A.; Roldán, A.; Jorge, E.; García-Berrocal, J.R. Reactive oxygen species in apoptosis induced by cisplatin: Review of physiopathological mechanisms in animal models. Eur. Arch. Oto-Rhino-Laryngol. 2012, 269, 2455–2459. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, K.; Zhang, K.; Zhu, L.; Zhou, F. Ziyuglycoside II induces cell cycle arrest and apoptosis through activation of ROS/JNK pathway in human breast cancer cells. Toxicol. Lett. 2014, 227, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Gong, M.-W.; Peng, Z.-F.; Zhou, T.; Ying, M.-G.; Zheng, Q.-H.; Liu, Q.-Y.; Zhang, Q.-Q. The marine fungal metabolite, dicitrinone B, induces A375 cell apoptosis through the ROS-related caspase pathway. Mar. Drugs 2014, 12, 1939–1958. [Google Scholar] [CrossRef] [Green Version]
- Yan, T.; Shang, L.; Wang, M.; Zhang, C.; Zhao, X.; Bi, K.; Jia, Y. Lignans from Schisandra chinensis ameliorate cognition deficits and attenuate brain oxidative damage induced by D-galactose in rats. Metab. Brain Dis. 2016, 31, 653–661. [Google Scholar] [CrossRef]
- Hu, D.; Yang, Z.; Yao, X.; Wang, H.; Han, N.; Liu, Z.; Wang, Y.; Yang, J.; Yin, J. Dibenzocyclooctadiene lignans from Schisandra chinensis and their inhibitory activity on NO production in lipopolysaccharide-activated microglia cells. Phytochemistry 2014, 104, 72–78. [Google Scholar] [CrossRef]
- Yu, H.-Y.; Chen, Z.-Y.; Sun, B.; Liu, J.; Meng, F.-Y.; Liu, Y.; Tian, T.; Jin, A.; Ruan, H.-L. Lignans from the fruit of Schisandra glaucescens with antioxidant and neuroprotective properties. J. Nat. Prod. 2014, 77, 1311–1320. [Google Scholar] [CrossRef]
- Xie, Y.; Hao, H.; Wang, H.; Guo, C.; Kang, A.; Wang, G. Reversing effects of lignans on CCl4-induced hepatic CYP450 down regulation by attenuating oxidative stress. J. Ethnopharmacol. 2014, 155, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Raffoul, J.J. Potential anticancer properties of grape antioxidants. J. Oncol. 2012, 2012, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Khamphio, M.; Barusrux, S.; Weerapreeyakul, N. Sesamol induces mitochondrial apoptosis pathway in HCT116 human colon cancer cells via pro-oxidant effect. Life Sci. 2016, 158, 46–56. [Google Scholar] [CrossRef]
- Nasser, M.I.; Han, T.; Adlat, S.; Tian, Y.; Jiang, N. Inhibitory effects of schisandrin b on human prostate cancer cells. Oncol. Rep. 2018, 41, 677–685. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Yang, W. Gomisin J inhibits the glioma progression by inducing apoptosis and reducing HKII-regulated glycolysis. Biochem. Biophys. Res. Commun. 2020, 529, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Meitzler, J.L.; Antony, S.; Wu, Y.; Juhasz, A.; Liu, H.; Jiang, G.; Lu, J.; Roy, K.; Doroshow, J.H. NADPH Oxidases: A perspective on reactive oxygen species production in tumor biology. Antioxid. Redox Signal. 2014, 20, 2873–2889. [Google Scholar] [CrossRef] [Green Version]
- Hou, D.; Liu, Z.; Xu, X.; Liu, Q.; Zhang, X.; Kong, B.; Wei, J.-J.; Gong, Y.; Shao, C. Increased oxidative stress mediates the antitumor effect of PARP inhibition in ovarian cancer. Redox Biol. 2018, 17, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, T.; Lambeth, J.D. Molecular evolution of Phox-related regulatory subunits for NADPH oxidase enzymes. BMC Evol. Biol. 2007, 7, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juhasz, A.; Ge, Y.; Markel, S.; Chiu, A.; Matsumoto, L.; Van Balgooy, J.; Roy, K.; Doroshow, J.H. Expression of NADPH oxidase homologues and accessory genes in human cancer cell lines, tumours and adjacent normal tissues. Free Radic. Res. 2009, 43, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Carmona-Cuenca, I.; Roncero, C.; Sancho, P.; Caja, L.; Fausto, N.; Fernández, M.; Fabregat, I. Upregulation of the NADPH oxidase NOX4 by TGF-beta in hepatocytes is required for its pro-apoptotic activity. J. Hepatol. 2008, 49, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Terazawa, R.; Kojima, K.; Nakane, K.; Deguchi, T.; Ando, M.; Tsukamasa, Y.; Ito, M.; Nozawa, Y. Cisplatin induces production of reactive oxygen species via NADPH oxidase activation in human prostate cancer cells. Free Radic. Res. 2011, 45, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
- Maraldi, T. Natural Compounds as modulators of NADPH oxidases. Oxidative Med. Cell. Longev. 2013, 2013, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ko, Y.H.; Jeong, M.; Jang, D.S.; Choi, J.-H. Gomisin L1, a Lignan Isolated from Schisandra Berries, Induces Apoptosis by Regulating NADPH Oxidase in Human Ovarian Cancer Cells. Life 2021, 11, 858. https://doi.org/10.3390/life11080858
Ko YH, Jeong M, Jang DS, Choi J-H. Gomisin L1, a Lignan Isolated from Schisandra Berries, Induces Apoptosis by Regulating NADPH Oxidase in Human Ovarian Cancer Cells. Life. 2021; 11(8):858. https://doi.org/10.3390/life11080858
Chicago/Turabian StyleKo, Young Hyun, Miran Jeong, Dae Sik Jang, and Jung-Hye Choi. 2021. "Gomisin L1, a Lignan Isolated from Schisandra Berries, Induces Apoptosis by Regulating NADPH Oxidase in Human Ovarian Cancer Cells" Life 11, no. 8: 858. https://doi.org/10.3390/life11080858