Noncoding RNA: An Insight into Chloroplast and Mitochondrial Gene Expressions


Abstract
:1. Introduction
2. Chloroplast: Genome Structure and Regulation
3. Noncoding RNAs in Chloroplasts as a Prospective Regulatory Molecule
4. Plant Mitochondria: Genome Structure and Regulation
5. Noncoding RNAs in Plant Mitochondria as a Prospective Regulatory Factor
6. Most Potential Factors Accountable for Chloroplast and Plant Mitochondrial ncRNAs Biogenesis
7. Methods and Challenges to Identify Noncoding RNAs from Plant Organelles
8. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shin, S.Y.; Jeong, J.S.; Lim, J.Y.; Kim, T.; Park, J.H.; Kim, J.K.; Shin, C. Transcriptomic analyses of rice (Oryza sativa) genes and noncoding RNAs under nitrogen starvation using multiple omics technologies. BMC Genom. 2018, 19, 532. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.I.; Alam, M.; Lightfoot, D.A.; Gurha, P.; Afzal, A.J. Classification and experimental identification of plant long non-coding RNAs. Genomics 2019, 111, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Axtell, M.J. Classification and comparison of small RNAs from plants. Annu. Rev. Plant Biol. 2013, 64, 137–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Q.; Guan, X.; Chen, Z.J. Dynamic roles for small RNAs and DNA methylation during ovule and fiber development in allotetraploid cotton. PLoS Genet. 2015, 11, e1005724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunardon, A.; Johnson, N.R.; Hagerott, E.; Phifer, T.; Polydore, S.; Coruh, C.; Axtell, M.J. Integrated annotations and analyses of small RNA-producing loci from 47 diverse plants. Genome Res. 2020, 30, 497–513. [Google Scholar] [CrossRef]
- Ha, M.; Lu, J.; Tian, L.; Ramachandran, V.; Kasschau, K.D.; Chapman, E.J.; Carrington, J.C.; Chen, X.; Wang, X.J.; Chen, Z.J. Small RNAs serve as a genetic buffer against genomic shock in Arabidopsis interspecific hybrids and allopolyploids. Proc. Natl. Acad. Sci. USA 2009, 106, 17835–17840. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, R.J.; He, Y.; Valdes-Lopez, O.; Khan, S.M.; Joshi, T.; Urich, M.A.; Nery, J.R.; Diers, B.; Xu, D.; Stacey, G.; et al. Epigenome-wide inheritance of cytosine methylation variants in a recombinant inbred population. Genome Res. 2013, 23, 1663–1674. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.S.; Zhang, X.; Zhang, W.; Shen, D.; Wang, H.; Xia, Y.; Qiu, Y.; Song, J.; Wang, C.; Li, X. The association of changes in DNA methylation with temperature-dependent sex determination in cucumber. J. Exp. Bot. 2017, 68, 2899–2912. [Google Scholar] [CrossRef]
- Li, S.; Castillo-Gonzalez, C.; Yu, B.; Zhang, X. The functions of plant small RNAs in development and in stress responses. Plant J. 2017, 90, 654–670. [Google Scholar] [CrossRef] [Green Version]
- Deng, P.; Muhammad, S.; Cao, M.; Wu, L. Biogenesis and regulatory hierarchy of phased small interfering rnas in plants. Plant Biotechnol. J. 2018, 16, 965–975. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Gautam, V.; Singh, S.; Das, S.S.; Verma, S.; Mishra, V.; Mukherjee, S.; Sarkar, A.K. Plant small RNAs: Advancement in the understanding of biogenesis and role in plant development. Planta 2018, 248, 545–558. [Google Scholar] [CrossRef] [PubMed]
- De Felippes, F.F. Gene Regulation Mediated by microRNA-Triggered Secondary Small RNAs in Plants. Plants 2019, 8, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Li, Y.; Cao, X.; Qi, Y. MicroRNAs and Their Regulatory Roles in Plant-Environment Interactions. Annu. Rev. Plant Biol. 2019, 70, 489–525. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-Q.; Allan, A.C.; Yin, X.-R. Small RNAs with a big impact on horticultural traits. Crit. Rev. Plant Sci. 2020, 39, 30–43. [Google Scholar] [CrossRef]
- Velayudha Vimala Kumar, K.; Srikakulam, N.; Padbhanabhan, P.; Pandi, P. Deciphering microRNAs and Their Associated Hairpin Precursors in a Non-Model Plant, Abelmoschus esculentus. Non Coding RNA 2017, 3, 19. [Google Scholar] [CrossRef] [PubMed]
- Hotto, A.M.; Huston, Z.E.; Stern, D.B. Overexpression of a natural chloroplast-encoded antisense RNA in tobacco destabilizes 5S rRNA and retards plant growth. BMC Plant Biol. 2010, 10, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zghidi-Abouzid, O.; Merendino, L.; Buhr, F.; Ghulam, M.M.; Lerbs-Mache, S. Characterization of plastid psbT sense and antisense RNAs. Nucleic Acids Res. 2011, 39, 5379–5387. [Google Scholar] [CrossRef]
- Maier, U.G.; Bozarth, A.; Funk, H.T.; Zauner, S.; Rensing, S.A.; Schmitz-Linneweber, C.; Börner, T.; Tillich, M. Complex chloroplast RNA metabolism: Just debugging the genetic programme? BMC Biol. 2008, 6, 36. [Google Scholar] [CrossRef] [Green Version]
- Hammani, K.; Giege, P. RNA metabolism in plant mitochondria. Trends Plant Sci. 2014, 19, 380–389. [Google Scholar] [CrossRef]
- Holec, S.; Lange, H.; Kühn, K.; Alioua, M.; Börner, T.; Gagliardi, D. Relaxed transcription in Arabidopsis mitochondria is counterbalanced by RNA stability control mediated by polyadenylation and polynucleotide phosphorylase. Mol. Cell. Biol. 2006, 26, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- Holec, S.; Lange, H.; Canaday, J.; Gagliardi, D. Coping with cryptic and defective transcripts in plant mitochondria. Biochim. Biophys. Acta 2008, 1779, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Steglich, C.; Futschik, M.E.; Lindell, D.; Voss, B.; Chisholm, S.W.; Hess, W.R. The challenge of regulation in a minimal photoautotroph: Non-coding RNAs in Prochlorococcus. PLoS Genet. 2008, 4, e1000173. [Google Scholar] [CrossRef]
- Ranjan, T.; Kumar, M.; Prasad, B.D.; Kumar, R.R.; Pal, A.K.; Ansar, M.; Singh, B. Chapter 13—Cyanobacterial stress-responsive small RNAs (sRNAs): Players of stress and developmental responses. In Advances in Cyanobacterial Biology; Singh, P.K., Kumar, A., Singh, V.K., Shrivastava, A.K., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 195–204. [Google Scholar]
- Vendramin, R.; Marine, J.-C.; Leucci, E. Non-coding RNAs: The dark side of nuclear–mitochondrial communication. EMBO J. 2017, 36, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.B.; Aronoff, S.; Boardman, N.K.; Bogorad, L.; Boucher, L.J.; Butler, W.L.; Clayton, R.K.; Cohen-Bazire, G.; Dougherty, R.R.; Goedheer, J.C.; et al. The Chlorophylls; Vernon, L.P., Seely, G.R., Eds.; Academic Press: Cambridge, MA, USA, 1966; p. xv. [Google Scholar]
- Staehelin, L.A. Chloroplast structure: From chlorophyll granules to supra-molecular architecture of thylakoid membranes. Photosynth. Res. 2003, 76, 185–196. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. The origins of plastids. Biol. J. Linn. Soc. 1982, 17, 289–306. [Google Scholar] [CrossRef]
- Sakai, A.; Takano, H.; Kuroiwa, H. Organelle nuclei in higher plants: Structure, composition, function, and evolution. Int. Rev. Cytol. 2004, 238, 59–118. [Google Scholar]
- Krupinska, K.; Melonek, J.; Krause, K. New insights into plastid nucleoid structure and functionality. Planta 2013, 237, 653–664. [Google Scholar] [CrossRef] [Green Version]
- Daniell, H.; Lin, C.S.; Yu, M.; Chang, W.J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef] [Green Version]
- Dobrogojski, J.; Adamiec, M.; Lucinski, R. The chloroplast genome: A review. Acta Physiol. Plant 2020, 42, 98. [Google Scholar] [CrossRef]
- Hotto, A.M.; Germain, A.; Stern, D.B. Plastid non-coding RNAs: Emerging candidates for gene regulation. Trends Plant Sci. 2012, 17, 737–744. [Google Scholar] [CrossRef]
- Zoschke, R.; Liere, K.; Borner, T. From seedling to mature plant: Arabidopsis plastidial genome copy number, RNA accumulation and transcription are differentially regulated during leaf development. Plant J. 2007, 50, 710–722. [Google Scholar] [CrossRef] [PubMed]
- Tadini, L.; Jeran, N.; Peracchio, C.; Masiero, S.; Colombo, M.; Pesaresi, P. The plastid transcription machinery and its coordination with the expression of nuclear genome: Plastid-encoded polymerase, nuclear encoded polymerase and the genomes uncoupled 1-mediated retrograde communication. Phil. Trans. R. Soc. B 2020, 375, 20190399. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, E.M. Post-transcriptional control of chloroplast gene expression. Gene Regul. Syst. Biol. 2009, 3, 31–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prikryl, J.; Rojas, M.; Schuster, G.; Barkan, A. Mechanism of RNA stabilization and translational activation by a pentatricopeptide repeat protein. Proc. Natl. Acad. Sci. USA 2011, 108, 415–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhelyazkova, P.; Hammani, K.; Rojas, M.; Voelker, R.; Vargas-Suarez, M.; Borner, T.; Barkan, A. Protein-mediated protection as the predominant mechanism for defining processed mRNA termini in land plant chloroplasts. Nucleic Acids Res. 2012, 40, 3092–3105. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Tan, Z.; Wu, F.; Sheng, P.; Heng, Y.; Wang, X.; Ren, Y.; Wang, J.; Guo, X.; Zhang, X.; et al. A novel chloroplast-localized pentatricopeptide repeat protein involved in splicing affects chloroplast development and abiotic stress response in rice. Mol. Plant 2014, 7, 1329–1349. [Google Scholar] [CrossRef] [Green Version]
- Yerramsetty, P.; Stata, M.; Siford, R.; Sage, T.L.; Sage, R.F.; Wong, G.K.-S.; Albert, V.A.; Berry, J.O. Evolution of RLSB, a nuclear-encoded S1 domain RNA binding protein associated with post-transcriptional regulation of plastid-encoded rbcL mRNA in vascular plants. BMC Evol. Biol. 2016, 16, 141. [Google Scholar] [CrossRef] [Green Version]
- Gutteridge, S.; Manavski, N.; Schmid, L.-M.; Meurer, J. RNA-stabilization factors in chloroplasts of vascular plants. Essays Biochem. 2018, 62, 51–64. [Google Scholar] [CrossRef] [Green Version]
- Pfannschmidt, T.; Yang, C. The hidden function of photosynthesis: A sensing system for environmental conditions that regulates plant acclimation responses. Protoplasma 2012, 249 (Suppl. S2), S125–S136. [Google Scholar] [CrossRef]
- Bradbeer, J.W.; Atkinson, Y.E.; Borner, T.; Hagemann, R. Cytoplasmic synthesis of plastid polypeptides may be controlled by plastid-synthesized RNA. Nature 1979, 279, 816–817. [Google Scholar] [CrossRef]
- Feng, P.; Guo, H.; Chi, W.; Chai, X.; Sun, X.; Xu, X.; Ma, J.; Rochaix, J.D.; Leister, D.; Wang, H.; et al. Chloroplast retrograde signal regulates flowering. Proc. Natl. Acad. Sci. USA 2016, 113, 10708–10713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobik, K.; Burch-Smith, T.M. Chloroplast signaling within, between and beyond cells. Front. Plant. Sci. 2015, 6, 781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, W.; Feng, P.; Ma, J.; Zhang, L. Metabolites and chloroplast retrograde signaling. Curr. Opin. Plant Biol. 2015, 25, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Brautigam, K.; Dietzel, L.; Pfannschmidt, T. Plastid-nucleus communication: Anterograde and retrograde signaling in the development and function of plastids. In Cell and Molecular Biology of Plastids; Springer: Berlin/Heidelberg, Germany, 2007; pp. 409–455. [Google Scholar]
- Yoo, C.Y.; Han, S.; Chen, M. Nucleus-to-Plastid Phytochrome Signaling in Controlling Chloroplast Biogenesis. Annu. Plant Rev. 2020, 3, 251–280. [Google Scholar]
- Fei, X.T.; Shi, Q.Q.; Liu, Y.L.; Yang, T.X.; Wei, A.Z. RNA sequencing and functional analyses reveal regulation of novel drought-responsive, long-non-coding RNA in Zanthoxylum bungeanum Maxim. Plant Growth Regul. 2020, 90, 425–440. [Google Scholar] [CrossRef]
- Shi, C.; Wang, S.; Xia, E.H.; Jiang, J.-J.; Zeng, F.-C.; Gao, L.-Z. Full transcription of the chloroplast genome in photosynthetic eukaryotes. Sci. Rep. 2016, 6, 30135. [Google Scholar] [CrossRef]
- Zoschke, R.; Bock, R. Chloroplast Translation: Structural and Functional Organization, Operational Control, and Regulation. Plant Cell 2018, 30, 745–770. [Google Scholar] [CrossRef] [Green Version]
- Robles, P.; Quesada, V. Transcriptional and Post-transcriptional Regulation of Organellar Gene Expression (OGE) and Its Roles in Plant Salt Tolerance. Int. J. Mol. Sci. 2019, 20, 1056. [Google Scholar] [CrossRef] [Green Version]
- Stern, D.B.; Gruissem, W. Control of plastid gene expression: 3′ inverted repeats act as mRNA processing and stabilizing elements, but do not terminate transcription. Cell 1987, 51, 1145–1157. [Google Scholar] [CrossRef]
- Mullet, J.E.; Klein, R.R. Transcription and RNA stability are important determinants of higher plant chloroplast RNA levels. EMBO J. 1987, 6, 1571–1579. [Google Scholar] [CrossRef]
- Mercer, T.R.; Neph, S.; Dinger, M.E.; Crawford, J.; Smith, M.A.; Shearwood, A.M.; Haugen, E.; Bracken, C.P.; Rackham, O.; Stamatoyannopoulos, J.A.; et al. The human mitochondrial transcriptome. Cell 2011, 146, 645–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larriba, E.; Rial, E.; del Mazo, J. The landscape of mitochondrial small non-coding RNAs in the PGCs of male mice, spermatogonia, gametes and in zygotes. BMC Genom. 2018, 19, 634. [Google Scholar] [CrossRef] [PubMed]
- Thairu, M.W.; Hansen, A.K. It’s a small, small world: Unravelling the role and evolution of small RNAs in organelle and endosymbiont genomes. FEMS Microbiol. Lett. 2019, 366, fnz049. [Google Scholar] [CrossRef] [PubMed]
- Demarsy, E.; Buhr, F.; Lambert, E.; Lerbs-Mache, S. Characterization of the plastid-specific germination and seedling establishment transcriptional programme. J. Exp. Bot. 2012, 63, 925–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Kang, Y.; Shin, S.C.; Park, H.; Lee, H. Combined Analysis of the Chloroplast Genome and Transcriptome of the Antarctic Vascular Plant Deschampsia antarctica Desv. PLoS ONE 2014, 9, e92501. [Google Scholar] [CrossRef] [Green Version]
- Loizeau, K.; Qu, Y.; Depp, S.; Fiechter, V.; Ruwe, H.; Lefebvre-Legendre, L.; Schmitz-Linneweber, C.; Goldschmidt-Clermont, M. Small RNAs reveal two target sites of the RNA-maturation factor Mbb1 in the chloroplast of Chlamydomonas. Nucleic Acids Res. 2014, 42, 3286–3297. [Google Scholar] [CrossRef] [Green Version]
- Tajima, N.; Sato, S.; Maruyama, F.; Kurokawa, K.; Ohta, H.; Tabata, S.; Sekine, K.; Moriyama, T.; Sato, N. Analysis of the complete plastid genome of the unicellular red alga Porphyridium purpureum. J. Plant Res. 2014, 127, 389–397. [Google Scholar] [CrossRef]
- Dorrell, R.G.; Hinksman, G.A.; Howe, C.J. Diversity of transcripts and transcript processing forms in plastids of the dinoflagellate alga Karenia mikimotoi. Plant Mol. Biol. 2016, 90, 233–247. [Google Scholar] [CrossRef] [Green Version]
- Facella, P.; Carbone, F.; Placido, A.; Perrotta, G. Cryptochrome 2 extensively regulates transcription of chloroplast genome in tomato. FEBS Open Bio 2017, 7, 456–471. [Google Scholar] [CrossRef]
- Dou, Y.; Li, S.; Yang, W.; Liu, K.; Du, Q.; Ren, G.; Yu, B.; Zhang, C. Genome-wide Discovery of Circular RNAs in the Leaf and Seedling Tissues of Arabidopsis Thaliana. Curr. Genom. 2017, 18, 360–365. [Google Scholar] [CrossRef] [Green Version]
- Amiryousefi, A.; Hyvonen, J.; Poczai, P. The chloroplast genome sequence of bittersweet (Solanum dulcamara): Plastid genome structure evolution in Solanaceae. PLoS ONE 2018, 13, e0196069. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Jiang, Z.; Gao, L.; You, C.; Ma, X.; Wang, X.; Xu, X.; Mo, B.; Chen, X.; Liu, L. Genome-Wide Transcript and Small RNA Profiling Reveals Transcriptomic Responses to Heat Stress. Plant Physiol. 2019, 181, 609–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Wang, Q.; Li, X.; Wang, G.; Wan, Y. Detecting of chloroplast circular RNAs in Arabidopsis thaliana. Plant Signal. Behav. 2019, 14, 1621088. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shi, D.; Bai, Y.; Liu, Y. High-throughput sequencing uncover Ficus tikoua Bur. chloroplast genome. J. Plant Biochem. Biotechnol. 2020, 29, 171–182. [Google Scholar] [CrossRef]
- Lung, B.; Zemann, A.; Madej, M.J.; Schuelke, M.; Techritz, S.; Ruf, S.; Bock, R.; Huttenhofer, A. Identification of small non-coding RNAs from mitochondria and chloroplasts. Nucleic Acids Res. 2006, 34, 3842–3852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohorianu, I.; Schwach, F.; Jing, R.; Lopez-Gomollon, S.; Moxon, S.; Szittya, G.; Sorefan, K.; Moulton, V.; Dalmay, T. Profiling of short RNAs during fleshy fruit development reveals stage-specific sRNAome expression patterns. Plant J. 2011, 67, 232–246. [Google Scholar] [CrossRef]
- Gonzalez-Ibeas, D.; Blanca, J.; Donaire, L.; Saladie, M.; Mascarell-Creus, A.; Cano-Delgado, A.; Garcia-Mas, J.; Llave, C.; Aranda, M.A. Analysis of the melon (Cucumis melo) small RNAome by high-throughput pyrosequencing. BMC Genom. 2011, 12, 393. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yu, X.; Wang, H.; Lu, Y.-Z.; de Ruiter, M.; Prins, M.; He, Y.-K. A novel class of heat-responsive small RNAs derived from the chloroplast genome of Chinese cabbage (Brassica rapa). BMC Genom. 2011, 12, 289. [Google Scholar] [CrossRef] [Green Version]
- Zhelyazkova, P.; Sharma, C.M.; Forstner, K.U.; Liere, K.; Vogel, J.; Borner, T. The primary transcriptome of Barley chloroplasts:Numerous noncoding RNAs and the dominating role of the plastid-encoded RNA polymerase. Plant Cell 2012, 24, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Hackenberg, M.; Shi, B.-J.; Gustafson, P.; Langridge, P. A transgenic transcription factor (TaDREB3) in barley affects the expression of microRNAs and other small non-coding RNAs. PLoS ONE 2012, 7, e42030. [Google Scholar] [CrossRef] [Green Version]
- Sablok, G.; Yang, K.; Chen, R.; Wen, X. tRNA derived smallRNAs: smallRNAs repertoire has yet to be decoded in plants. Front. Plant Sci. 2017, 8, 1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Xu, Z.; Sheng, J. tRNA-derived small RNA: A novel regulatory small non-coding RNA. Genes 2018, 9, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, F.; Guo, Z. Emerging role of a novel small non-coding regulatory RNA: tRNA-derived small RNA. ExRNA 2019, 1, 39. [Google Scholar] [CrossRef] [Green Version]
- Cognat, V.; Morelle, G.; Megel, C.; Lalande, S.; Molinier, J.; Vincent, T.; Small, I.; Duchene, A.M.; Marechal-Drouard, L. The nuclear and organellar tRNA-derived RNA fragment population in Arabidopsis thaliana is highly dynamic. Nucleic Acids Res. 2017, 45, 3460–3472. [Google Scholar] [CrossRef] [PubMed]
- Cavaiuolo, M.; Kuras, R.; Wollman, F.-A.; Choquet, Y.; Vallon, O. Small RNA profiling in Chlamydomonas: Insights into chloroplast RNA metabolism. Nucleic Acids Res. 2017, 45, 10783–10799. [Google Scholar] [CrossRef] [Green Version]
- Dorrell, R.G.; Nisbet, R.E.R.; Barbrook, A.C.; Rowden, S.J.L.; Howe, C.J. Integrated Genomic and Transcriptomic Analysis of the Peridinin Dinoflagellate Amphidinium carterae Plastid. Protist 2019, 170, 358–373. [Google Scholar] [CrossRef]
- Hotto, A.M.; Schmitz, R.J.; Fei, Z.; Ecker, J.R.; Stern, D.B. Unexpected diversity of chloroplast noncoding RNAs as revealed by deep sequencing of the Arabidopsis transcriptome. G3 2011, 1, 559–570. [Google Scholar] [CrossRef] [Green Version]
- Ruwe, H.; Schmitz-Linneweber, C. Short non-coding RNA fragments accumulating in chloroplasts: Footprints of RNA binding proteins? Nucleic Acids Res. 2012, 40, 3106–3116. [Google Scholar] [CrossRef]
- Ruwe, H.; Gutmann, B.; Schmitz-Linneweber, C.; Small, I.; Kindgren, P. The E domain of CRR2 participates in sequence-specific recognition of RNA in plastids. New Phytol. 2019, 222, 218–229. [Google Scholar] [CrossRef]
- Castandet, B.; Germain, A.; Hotto, A.M.; Stern, D.B. Systematic sequencing of chloroplast transcript termini from Arabidopsis thaliana reveals >200 transcription initiation sites and the extensive imprints of RNA-binding proteins and secondary structures. Nucleic Acids Res 2019, 47, 11889–11905. [Google Scholar] [CrossRef] [Green Version]
- Georg, J.; Honsel, A.; Voß, B.; Rennenberg, H.; Hess, W.R. A long antisense RNA in plant chloroplasts. New Phytol. 2010, 186, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Sharwood, R.E.; Hotto, A.M.; Bollenbach, T.J.; Stern, D.B. Overaccumulation of the chloroplast antisense RNA AS5 is correlated with decreased abundance of 5S rRNA in vivo and inefficient 5S rRNA maturation in vitro. RNA 2011, 17, 230–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Zhang, J.; Yuan, G.; Liu, C. Complex interplay among DNA modification, noncoding RNA expression and protein-coding RNA expression in Salvia miltiorrhiza chloroplast genome. PLoS ONE 2014, 9, e99314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, G.; Sharma, S.; Upadhyay, S.K.; Singh, K. Long non-coding RNAs coordinate developmental transitions and other key biological processes in grapevine. Sci. Rep. 2019, 9, 3552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, M.W.; Burger, G.; Lang, B.F. Mitochondrial evolution. Science 1999, 283, 1476–1481. [Google Scholar] [CrossRef] [Green Version]
- Andersson, S.G.; Karlberg, O.; Canback, B.; Kurland, C.G. On the origin of mitochondria: A genomics perspective. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2003, 358, 165–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesaresi, P.; Schneider, A.; Kleine, T.; Leister, D. Interorganellar communication. Curr. Opin. Plant Biol. 2007, 10, 600–606. [Google Scholar] [CrossRef]
- Dong, Y.; Yoshitomi, T.; Hu, J.F.; Cui, J. Long noncoding RNAs coordinate functions between mitochondria and the nucleus. Epigenet. Chromatin 2017, 10, 41. [Google Scholar] [CrossRef] [Green Version]
- Gualberto, J.M.; Newton, K.J. Plant Mitochondrial Genomes: Dynamics and Mechanisms of Mutation. Annu. Rev. Plant Biol. 2017, 68, 225–252. [Google Scholar] [CrossRef]
- Christensen, A.C. Mitochondrial DNA repair and genome evolution. Ann. Plant Rev. 2018, 50, 11–32. [Google Scholar]
- Gualberto, J.M.; Kuhn, K. DNA-binding proteins in plant mitochondria: Implications for transcription. Mitochondrion 2014, 19, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Kozik, A.; Rowan, B.A.; Lavelle, D.; Berke, L.; Schranz, M.E.; Michelmore, R.W.; Christensen, A.C. The alternative reality of plant mitochondrial DNA: One ring does not rule them all. PLoS Genet. 2019, 15, 1008373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unseld, M.; Marienfeld, J.R.; Brandt, P.; Brennicke, A. The mitochondrial genome of Arabidopsis thaliana contains 57 genes in 366,924 nucleotides. Nat. Genet. 1997, 15, 57–61. [Google Scholar] [CrossRef]
- Alverson, A.J.; Rice, D.W.; Dickinson, S.; Barry, K.; Palmer, J.D. Origins and recombination of the bacterial-sized multichromosomal mitochondrial genome of cucumber. Plant Cell 2011, 23, 2499–2513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, D.W.; Alverson, A.J.; Richardson, A.O.; Young, G.J.; Sanchez-Puerta, M.V.; Munzinger, J.; Barry, K.; Boore, J.L.; Zhang, Y.; dePamphilis, C.W.; et al. Horizontal transfer of entire genomes via mitochondrial fusion in the angiosperm Amborella. Science 2013, 342, 1468–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Stone, J.D.; Storchova, H.; Sloan, D.B. High transcript abundance, RNA editing, and small RNAs in intergenic regions within the massive mitochondrial genome of the angiosperm Silene noctiflora. BMC Genom. 2015, 16, 938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ro, S.; Ma, H.; Park, C.; Ortogero, N.; Song, R.; Hennig, G.W.; Zheng, H.; Lin, Y.M.; Moro, L.; Hsieh, J.T.; et al. The mitochondrial genome encodes abundant small noncoding RNAs. Cell Res. 2013, 23, 759–774. [Google Scholar] [CrossRef] [Green Version]
- Small, I.D.; Rackham, O.; Filipovska, A. Organelle transcriptomes: Products of a deconstructed genome. Curr. Opin. Microbiol. 2013, 16, 652–658. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, A.; Wallet, C.; Iqbal, R.K.; Gualberto, J.M.; Lotfi, F. Organellar non-coding RNAs: Emerging regulation mechanisms. Biochimie. 2015, 117, 48–62. [Google Scholar] [CrossRef]
- Allen, J.F.; Raven, J.A. Stefan Binder and Axel Brennicke.Gene expression in plant mitochondria: Transcriptional and post–transcriptional control. Phil. Trans. R. Soc. Lond. B 2003, 358, 181–189. [Google Scholar]
- Hedtke, B.; Borner, T.; Weihe, A. One RNA polymerase serving two genomes. EMBO Rep. 2000, 1, 435–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhn, K.; Weihe, A.; Borner, T. Multiple promoters are a common feature of mitochondrial genes in Arabidopsis. Nucleic Acids Res. 2005, 33, 337–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocak, S.; Linder, P. DEAD-box proteins: The driving forces behind RNA metabolism. Nat. Rev. Mol. Cell Biol. 2004, 5, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Kohler, D.; Schmidt-Gattung, S.; Binder, S. The DEAD-box protein PMH2 is required for efficient group II intron splicing in mitochondria of Arabidopsis thaliana. Plant Mol. Biol. 2010, 72, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Kang, H. Emerging roles of RNA-binding proteins in plant growth, development, and stress responses. Mol. Cells 2016, 39, 179–185. [Google Scholar]
- Burzio, V.A.; Villota, C.; Villegas, J.; Landerer, E.; Boccardo, E.; Villa, L.L.; Martinez, R.; Lopez, C.; Gaete, F.; Toro, V.; et al. Expression of a family of noncoding mitochondrial RNAs distinguishes normal from cancer cells. Proc. Natl. Acad. Sci. USA 2009, 106, 9430–9434. [Google Scholar] [CrossRef] [Green Version]
- Leucci, E.; Vendramin, R.; Spinazzi, M.; Laurette, P.; Fiers, M.; Wouters, J.; Radaelli, E.; Eyckerman, S.; Leonelli, C.; Vanderheyden, K.; et al. Melanoma addiction to the long non-coding RNA SAMMSON. Nature 2016, 531, 518–522. [Google Scholar] [CrossRef]
- Ruwe, H.; Wang, G.; Gusewski, S.; Schmitz-Linneweber, C. Systematic analysis of plant mitochondrial and chloroplast small RNAs suggests organelle-specific mRNA stabilization mechanisms. Nucleic Acids Res. 2016, 44, 7406–7417. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Kaul, S.; Rounsley, S.; Shea, T.P.; Benito, M.I.; Town, C.D.; Fujii, C.Y.; Mason, T.; Bowman, C.L.; Barnstead, M.; et al. Sequence and analysis of chromosome 2 of the plant Arabidopsis thaliana. Nature 1999, 402, 761–768. [Google Scholar] [CrossRef]
- Stone, J.D.; Kolouskova, P.; Sloan, D.B.; Storchova, H. Non-coding RNA may be associated with cytoplasmic male sterility in Silene vulgaris. J. Exp. Bot. 2017, 68, 1599–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Liu, X.; Xu, B.; Zhao, N.; Yang, X.; Zhang, M. Identification of miRNAs and their targets using high-throughput sequencing and degradome analysis in cytoplasmic male-sterile and its maintainer fertile lines of Brassica juncea. BMC Genom. 2013, 14, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Zhang, X.; Yao, Q.; Yuan, Y.; Li, X.; Wei, F.; Zhao, Y.; Zhang, Q.; Wang, Z.; Jiang, W.; et al. The miRNAs and their regulatory networks responsible for pollen abortion in Ogura-CMS Chinese cabbage revealed by high-throughput sequencing of miRNAs, degradomes, and transcriptomes. Front. Plant Sci. 2015, 6, 894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, X.; Li, J.; Zhang, H.; He, T.; Han, S.; Li, Y.; Yang, S.; Gai, J. Identification of miRNAs and their targets by high-throughput sequencing and degradome analysis in cytoplasmic male-sterile line NJCMS1A and its maintainer NJCMS1B of soybean. BMC Genom. 2016, 17, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karanthamalai, J.; Chodon, A.; Chauhan, S.; Pandi, G. DNA N6-methyladenine modification in plant genomes-A glimpse into emerging epigenetic code. Plants 2020, 9, 247. [Google Scholar] [CrossRef] [Green Version]
- Murik, O.; Chandran, S.A.; Nevo-Dinur, K.; Sultan, L.D.; Best, C.; Stein, Y.; Hazan, C. Ostersetzer-Biran, O. Topologies of N6-adenosine methylation (m6A) in land plant mitochondria and their putative effects on organellar gene expression. Plant J. 2020, 101, 1269–1286. [Google Scholar] [CrossRef]
- Meng, X.; Li, X.; Zhang, P.; Wang, J.; Zhou, Y.; Chen, M. Circular RNA: An emerging key player in RNA world. Brief. Bioinform. 2017, 18, 547–557. [Google Scholar] [CrossRef]
- Wilusz, J.E. A 360° view of circular RNAs: From biogenesis to functions. Wiley Interdiscip. Rev. RNA 2018, 9, e1478. [Google Scholar] [CrossRef] [Green Version]
- Guria, A.; Kumar, K.V.V.; Srikakulam, N.; Krishnamma, A.; Chanda, S.; Sharma, S.; Fan, X.; Pandi, G. Circular RNA profiling by Illumina sequencing via template-dependent multiple displacement amplification. BioMed Res. Int. 2019, 2019, 2756516. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Li, S.; Chen, M. Characterization and function of circular RNAs in plants. Front. Mol. Biosci. 2020, 7, 91. [Google Scholar] [CrossRef]
- Darbani, B.; Noeparvar, S.; Borg, S. Identification of circular RNAs from the parental genes involved in multiple aspects of cellular metabolism in barley. Front. Plant Sci. 2016, 7, 776. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Filipenko, S.J.; Darracq, A.; Adams, K.L. Expression of a transferred nuclear gene in a mitochondrial genome. Curr. Plant Biol. 2014, 1, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Glover, K.E.; Spencer, D.F.; Gray, M.W. Identification and structural characterization of nucleus-encoded transfer RNAs imported into wheat mitochondria. J. Biol. Chem. 2001, 276, 639–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salinas, T.; Duchêne, A.-M.; Delage, L.; Nilsson, S.; Glaser, E.; Zaepfel, M.; Maréchal-Drouard, L. The voltage-dependent anion channel, a major component of the tRNA import machinery in plant mitochondria. Proc. Natl. Acad. Sci. USA 2006, 103, 18362–18367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanita Lima, M.; Smith, D.R. Pervasive Genome-Wide Transcription in the organelle genomes of diverse plastid-bearing protists. G3 2017, 7, 3789–3796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharwood, R.E.; Halpert, M.; Luro, S.; Schuster, G.; Stern, D.B. Chloroplast RNase J compensates for inefficient transcription termination by removal of antisense RNA. RNA 2011, 17, 2165–2176. [Google Scholar] [CrossRef] [Green Version]
- Germain, A.; Herlich, S.; Larom, S.; Kim, S.H.; Schuster, G.; Stern, D.B. Mutational analysis of Arabidopsis chloroplast polynucleotide phosphorylase reveals roles for both RNase PH core domains in polyadenylation, RNA 3′-end maturation and intron degradation. Plant J. 2011, 67, 381–394. [Google Scholar] [CrossRef]
- Barkan, A.; Small, I. Pentatricopeptide repeat proteins in plants. Ann. Rev. Plant Biol. 2014, 65, 415–442. [Google Scholar] [CrossRef]
- Gomez, G.; Pallas, V. Studies on subcellular compartmentalization of plant pathogenic noncoding RNAs give new insights into the intracellular RNA-traffic mechanisms. Plant Physiol. 2012, 159, 558–564. [Google Scholar] [CrossRef] [Green Version]
- Barrey, E.; Saint-Auret, G.; Bonnamy, B.; Damas, D.; Boyer, O.; Gidrol, X. Pre-microRNA and mature microRNA in human mitochondria. PLoS ONE 2011, 6, e20220. [Google Scholar] [CrossRef]
- Sripada, L.; Tomar, D.; Singh, R. Mitochondria: One of the destinations of miRNAs. Mitochondrion 2012, 12, 593–599. [Google Scholar] [CrossRef]
- Sripada, L.; Tomar, D.; Prajapati, P.; Singh, R.; Singh, A.K.; Singh, R. Systematic Analysis of Small RNAs Associated with Human Mitochondria by Deep Sequencing: Detailed Analysis of Mitochondrial Associated miRNA. PLoS ONE 2012, 7, e44873. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-Y.; Li, Q.-Z.; Feng, Z.-X. Non-coding RNA identification based on topology secondary structure and reading frame in organelle genome level. Genomics 2016, 107, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Castandet, B.; Hotto, A.M.; Strickler, S.R.; Stern, D.B. ChloroSeq, an optimized chloroplast RNA-Seq bioinformatic pipeline, reveals remodeling of the organellar transcriptome under heat stress. G3 2016, 6, 2817–2827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigematsu, M.; Honda, S.; Loher, P.; Telonis, A.G.; Rigoutsos, I.; Kirino, Y. YAMAT-seq: An efficient method for high-throughput sequencing of mature transfer RNAs. Nucleic Acids Res. 2017, 45, e70. [Google Scholar] [CrossRef] [PubMed]
- Pawar, K.; Shigematsu, M.; Loher, P.; Honda, S.; Rigoutsos, I.; Kirino, Y. Exploration of CCA-added RNAs revealed the expression of mitochondrial non-coding RNAs regulated by CCA-adding enzyme. RNA Biol. 2019, 16, 1817–1825. [Google Scholar] [CrossRef]
- Small, I.; Marechal-Drouard, L.; Masson, J.; Pelletier, G.; Cosset, A.; Weil, J.H.; Dietrich, A. In vivo import of a normal or mutagenized heterologous transfer RNA into the mitochondria of transgenic plants: Towards novel ways of influencing mitochondrial gene expression? EMBO J. 1992, 11, 1291–1296. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, D.; Dreher, T.W. The tRNA-like structure of Turnip yellow mosaic virus RNA is a 3′-translational enhancer. Virology 2004, 321, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Val, R.; Wyszko, E.; Valentin, C.; Szymanski, M.; Cosset, A.; Alioua, M.; Dreher, T.W.; Barciszewski, J.; Dietrich, A. Organelle trafficking of chimeric ribozymes and genetic manipulation of mitochondria. Nucleic Acids Res. 2011, 39, 9262–9274. [Google Scholar] [CrossRef]
- Niazi, A.K.; Delannoy, E.; Iqbal, R.K.; Mileshina, D.; Val, R.; Gabryelska, M.; Wyszko, E.; Soubigou-Taconnat, L.; Szymanski, M.; Barciszewski, J.; et al. Mitochondrial transcriptome control and intercompartment cross-talk during plant development. Cells 2019, 8, 583. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Wang, R.; Lei, C.; Zhou, N. CRISPR-Cas System for RNA Detection and Imaging. Chem. Res. Chin. Univ. 2020, 36, 157–163. [Google Scholar] [CrossRef]
- Wang, H.; Nakamura, M.; Abbott, T.R.; Zhao, D.; Luo, K.; Yu, C.; Nguyen, C.M.; Lo, A.; Daley, T.P.; La Russa, M.; et al. CRISPR-mediated live imaging of genome editing and transcription. Science 2019, 365, 1301–1305. [Google Scholar] [CrossRef] [PubMed]



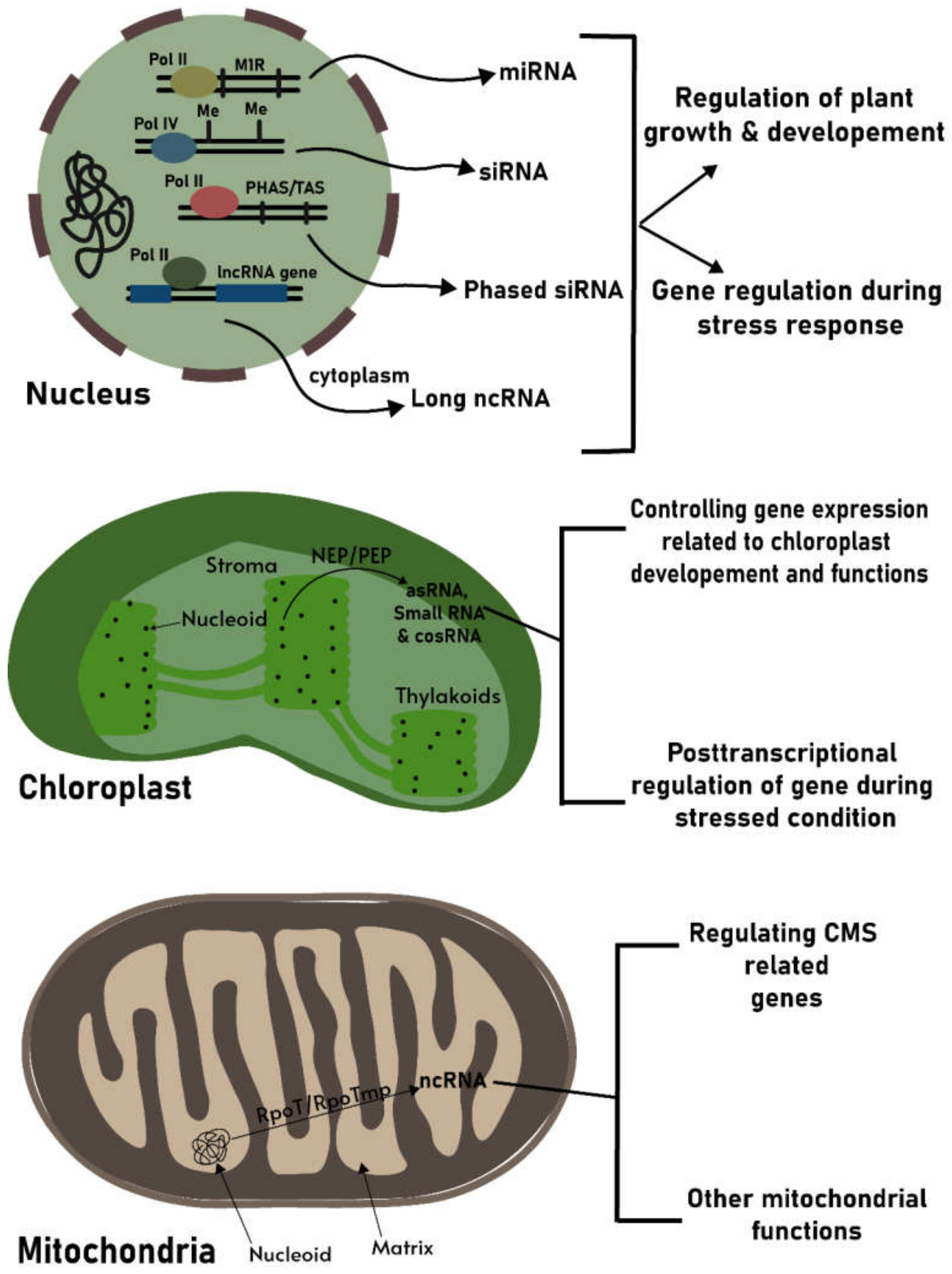{kind=link}
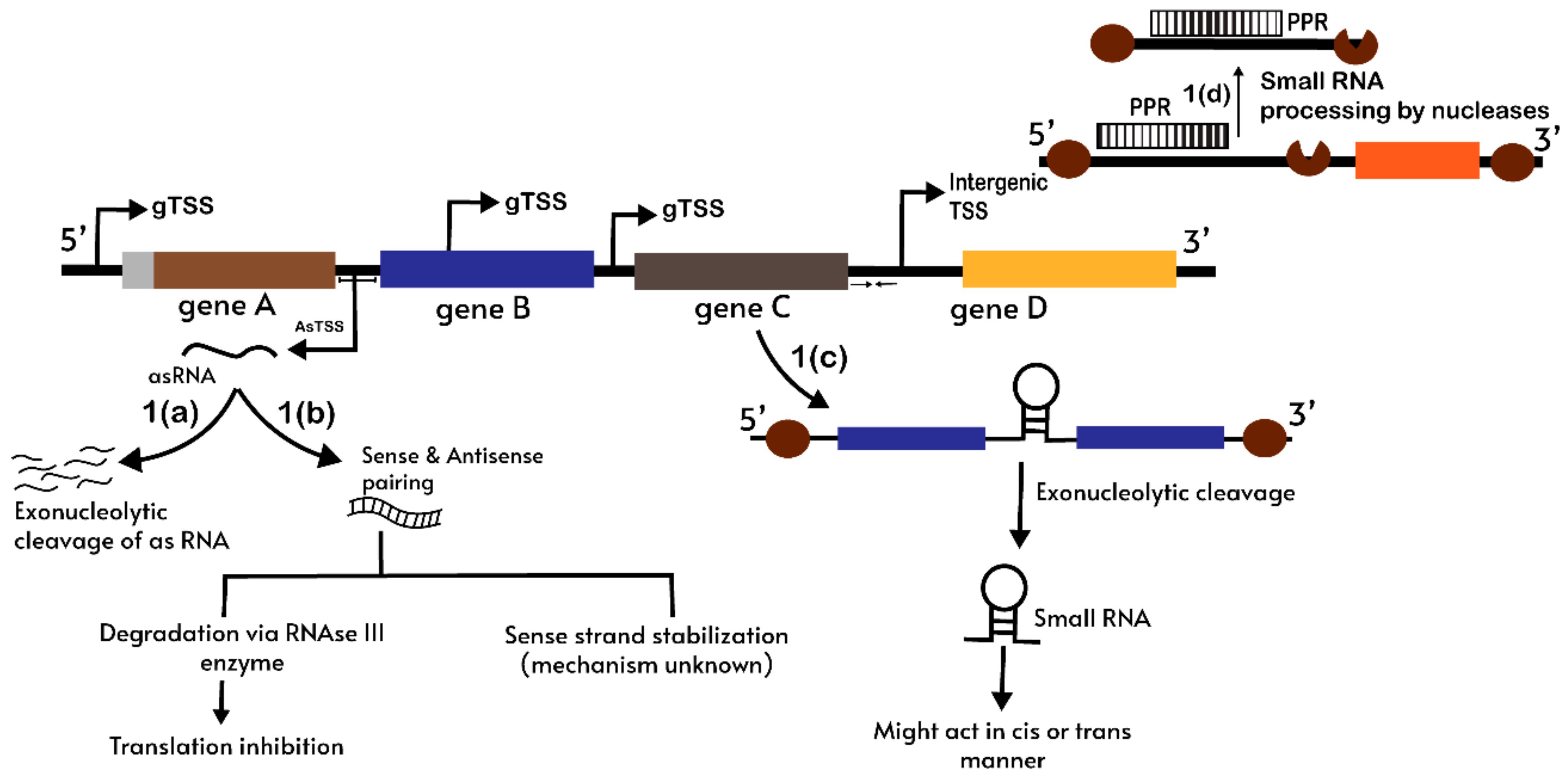{kind=link}
| Noncoding RNA. (Name, Numbers) | Length (nt) | Source (Plants/Algae) | Corresponding Sequence | ncRNA Detection Methodology | Reference |
|---|---|---|---|---|---|
| Antisense RNA | - | Arabidopsis thaliana | Opposite strand of rbcL, psbA and rpl33 | Macro array analysis | [57] |
| Small ncRNAs, 27 | 20–24 | Deschampsia antarctica Desv. | Located near 5′/3′ ends of genes, i.e., psbH-petB, atpH 5′ end, ndhB 5′ end, and petD_rpoA | RNA sequencing data analysis | [58] |
| Short RNA | 17–24 | Chlamydomonas reinhardtii | 5′ end of psbB and psbH | High-throughput RNA sequencing | [59] |
| rnpB | - | P. purpureum | - | Plastid genome sequencing | [60] |
| Antisense RNAs | - | Karenia mikimotoi (dinoflagellate) | psbA, psbD, ycf4, rps13 and rps11 | cDNA libreary sequencing, RT-PCR, cloning of 5′ end obtained from RACE | [61] |
| miRNA, 79 | 22 | Solanum lycopersicum (tomato) | Intergenic and intronic regions; Tpr and PPR, lucine rich repeat gene | miRBASE Search tool against tomato leaf trancriptome data | [62] |
| circRNAs | - | Arabidopsis thaliana | Exonic | Based on ncRNA sequencing dataset | [63] |
| ncRNA in IGS (intergenic spacer) region | - | Solanum dulcamara | Intergenic regions | RNA sequencing library from NCBI short read archive for Solanum dulcamara (SRR2056039) | [64] |
| sRNA | 19 | Zea mays ssp. mays | 3′ UTR of psbH gene | sRNA sequencing, Northern blot | [65] |
| circRNAs | - | Arabidopsis thaliana | Intronic, exonic, intergenic | circRNA sequencing | [66] |
| ncRNA | - | Ficus tikoua | - | CP genome sequencing and assembly | [67] |
| Potential Methods to Detect Organelle ncRNA | Description | Reference |
|---|---|---|
| Differential RNA sequencing (dRNA-seq) | dRNA-seq was used to discriminate primary transcript from processed 5′ ends by sequencing differential cDNA library | [72] |
| Terminome-seq | Library synthesis was conducted by using the Illumina TruSeq Small RNA library preparation kit, where the RNA populations containing a 5ʹ phosphate and 3ʹ hydroxyl group were captured | [83] |
| Algorithm-based ncRNA prediction | Secondary structure; three reading frames with triplates of nucleotide and motif information from ncRNA primary sequences were used as parameters to recognize ncRNAs via ID-SVM algorithm | [135] |
| ChloroSeq | A bioinformatics approach that is used to study chloroplast RNAs from strand-specific RNA-seq datasets | [136] |
| sRNA-miner | Publically available RNA-seq datasets were used to detect organelle ncRNAs | [111] |
| YAMAT-seq | ncRNA species with post-transcriptionally added signature secondary structure, i.e., 5′-NCCA-3′ or 5′-CCA-3′ protruding at 3′end can be detected | [137] |
| “PKTLS” (a chimeric transfer RNA-like structure with catalytic activity) | Knockdown of important organellar transcript | [141] |
| Fluorophore-labeled CRISPR-Cas-based probes | RNA imaging in live cells | [144] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anand, A.; Pandi, G. Noncoding RNA: An Insight into Chloroplast and Mitochondrial Gene Expressions. Life 2021, 11, 49. https://doi.org/10.3390/life11010049
Anand A, Pandi G. Noncoding RNA: An Insight into Chloroplast and Mitochondrial Gene Expressions. Life. 2021; 11(1):49. https://doi.org/10.3390/life11010049
Chicago/Turabian StyleAnand, Asha, and Gopal Pandi. 2021. "Noncoding RNA: An Insight into Chloroplast and Mitochondrial Gene Expressions" Life 11, no. 1: 49. https://doi.org/10.3390/life11010049