
Stochastic Modelling of Red Palm Weevil Using Chemical Injection and Pheromone Traps

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Mathematical Model
- In this model, the total date palm tree population is divided into two classes: susceptible date palm tree denoted by and infected date palm denoted by . In the absence of RPW, the date palm tree grows logistically with an intrinsic growth rate r and carrying capacity k. The natural death rate of the susceptible and infected date palm tree is .
- Assume that the date palm tree is susceptible to infection according to simple mass kinematics with as the RPW transmission coefficient. Experimental studies on palm pests indicated that the functional response pattern of the predator is consistent with the Holling II functional response [15,16]. As a result, we assume that RPW larvae harvest palm trees with Holing type-II functional response. The predation rate of RPW larvae on a date palm tree is c, and constant a is the half-saturation constant. The larvae population decreases by rate due to the transformation from the larva stage to the adult stage. The transition rate of RPW larvae to adults females is given by , whereas a complementing fraction will emerge as males . We assume that larvae decreases at the rate due to the injection of chemical compounds. The natural death of larvae is assumed to be , while the natural death for adult RPW is .
- To indicate the trap’s effect, one can consider the approach proposed by Barclay [17]. We assume the pheromone trap attracted additional females. As a result, the RPW males attracted to pheromone traps according to , where represents the effective rate of pheromone traps on mortality of RPW males. Recently, this approach was used by [18,19,20,21] to investigate the dynamics of the mirid population under mating disruption and trapping.
3. Dynamics of the Stochastic Model
- There is a positive number M such that
- There exists a non-negative function V such that is negative on
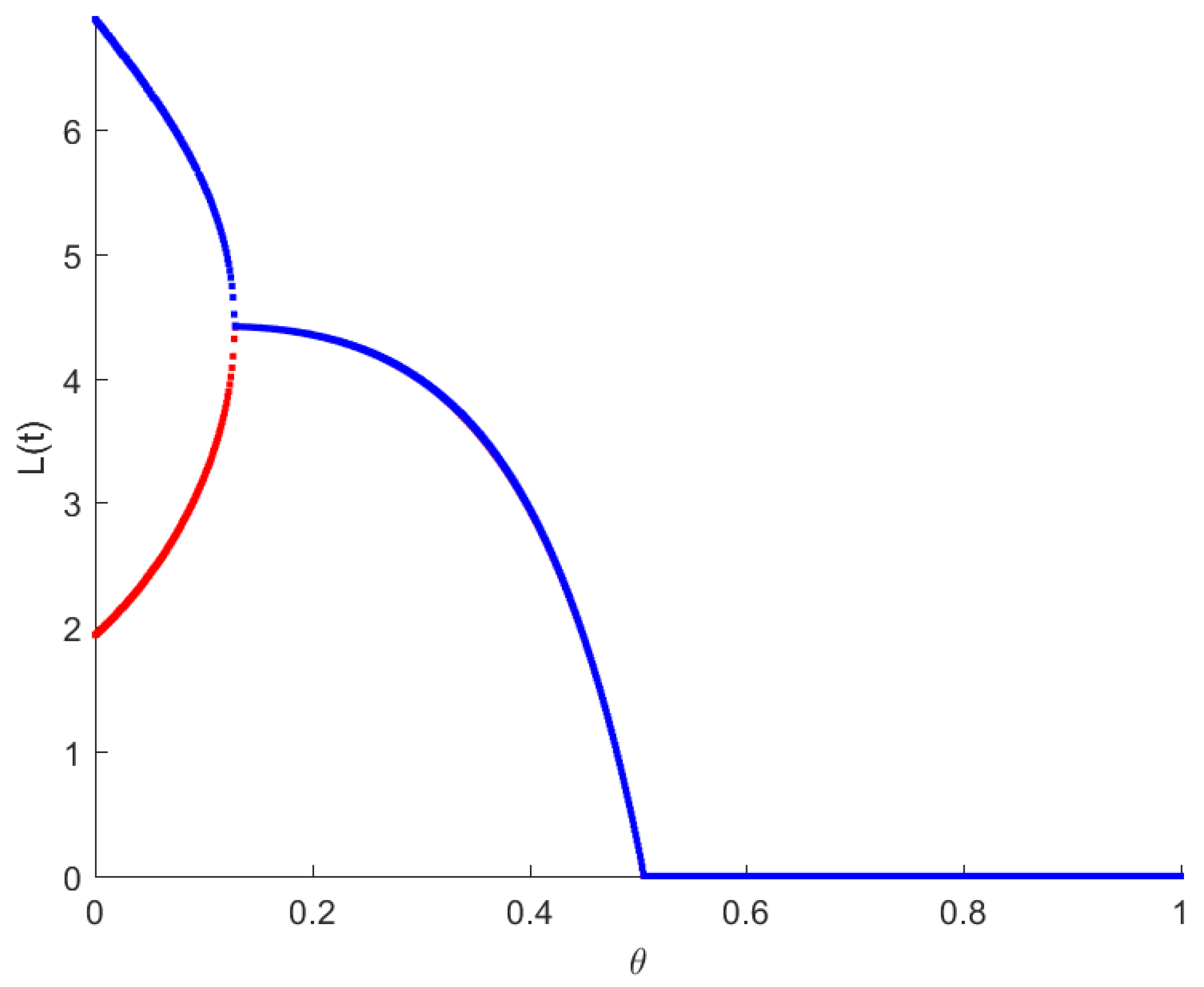
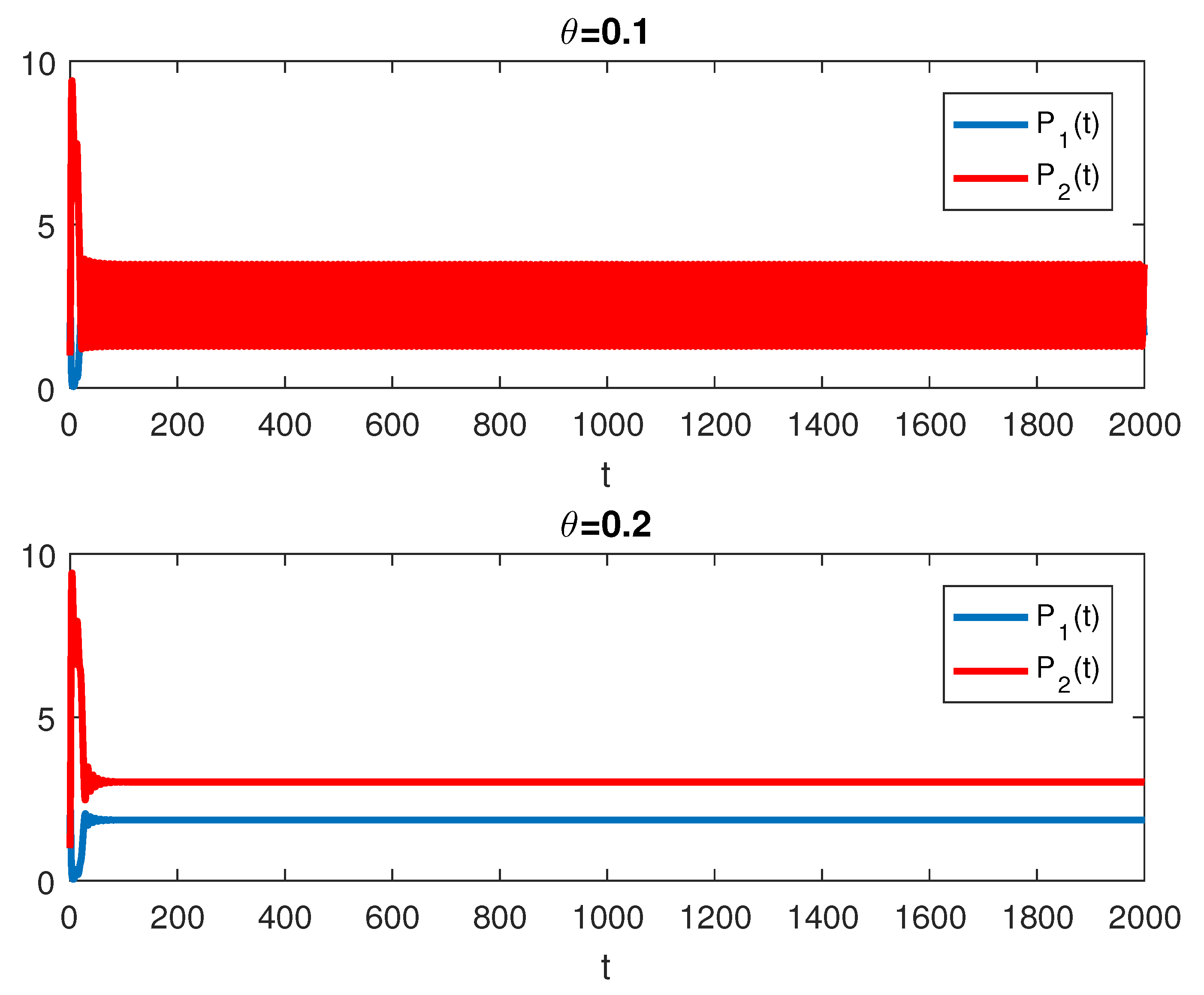
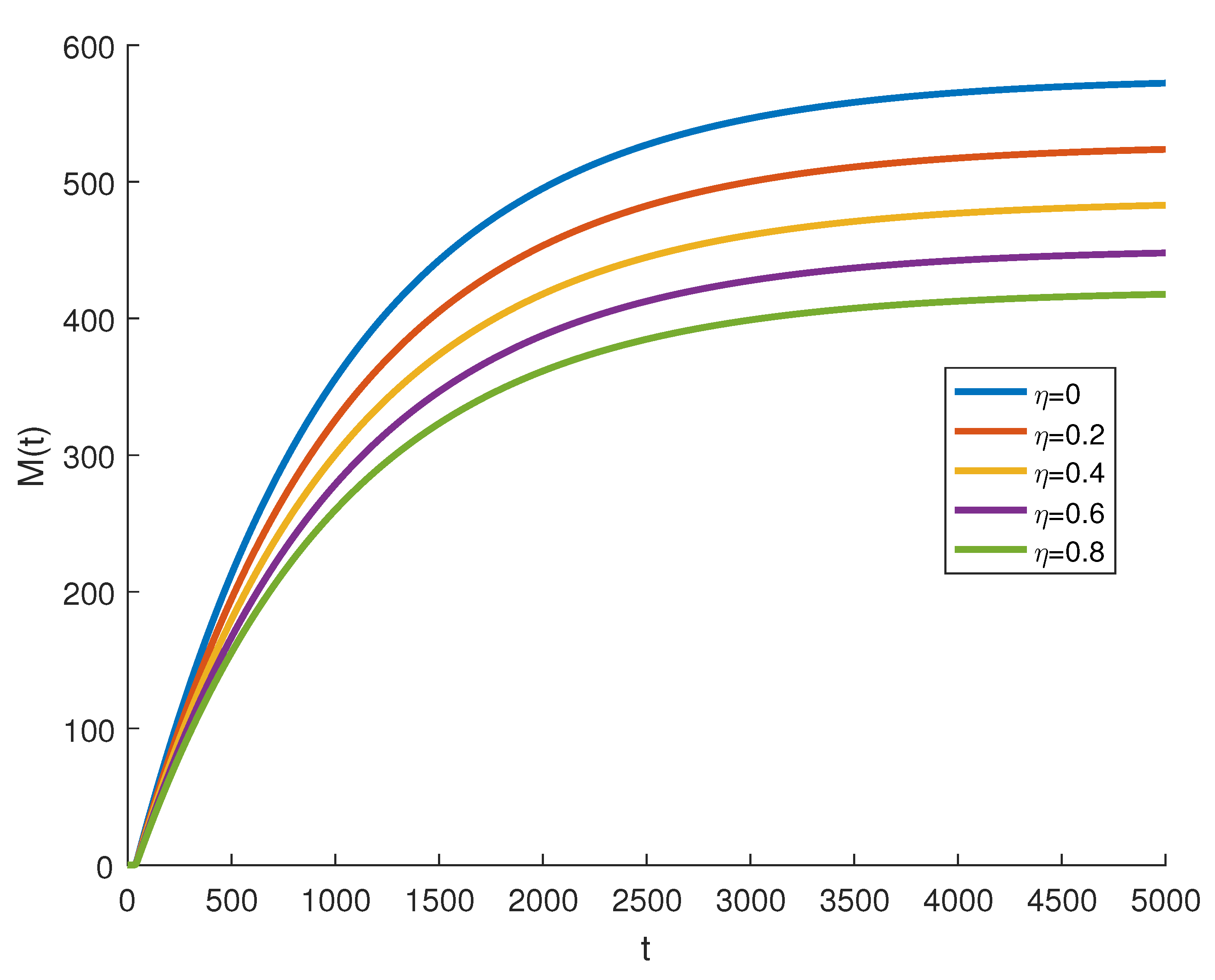
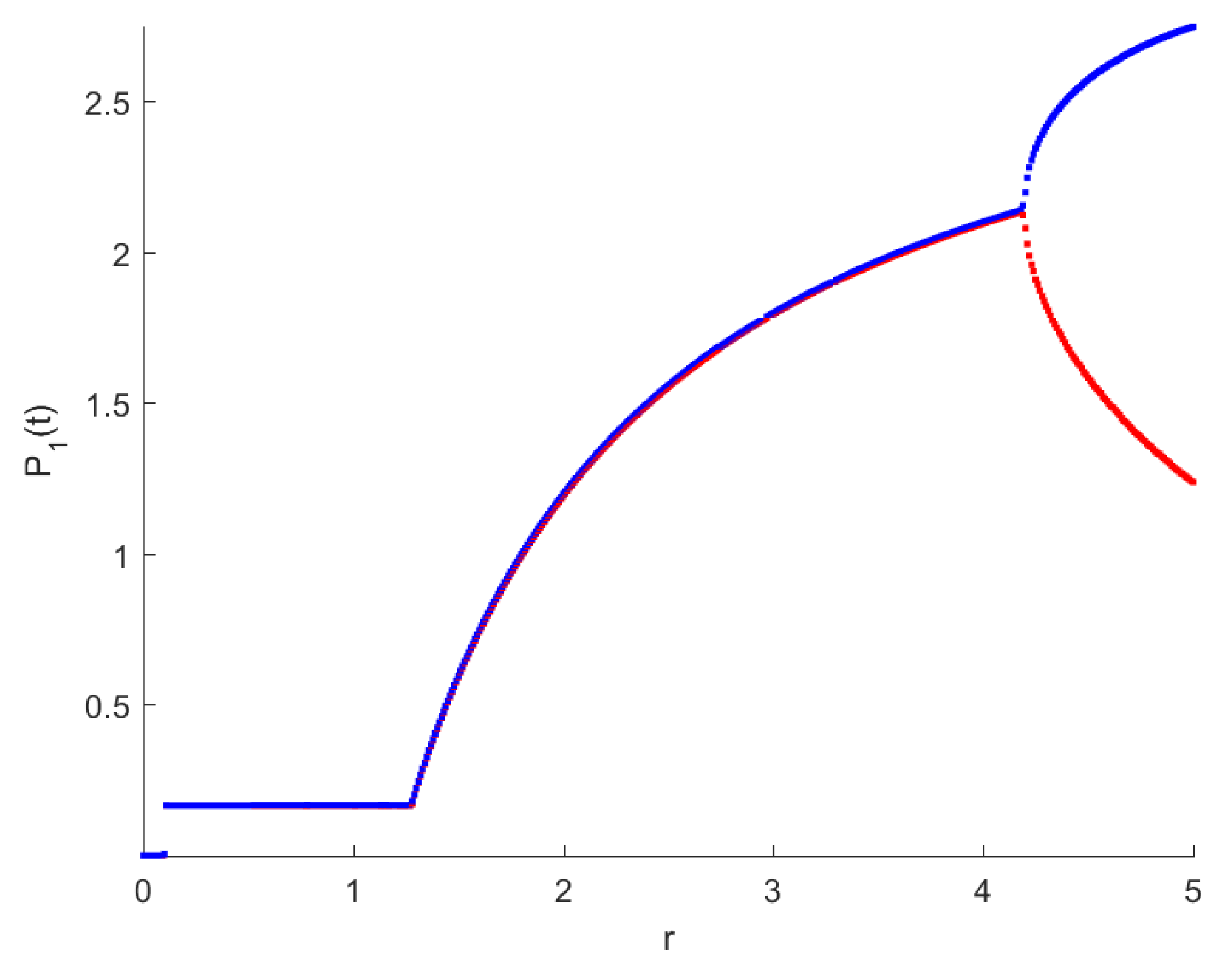
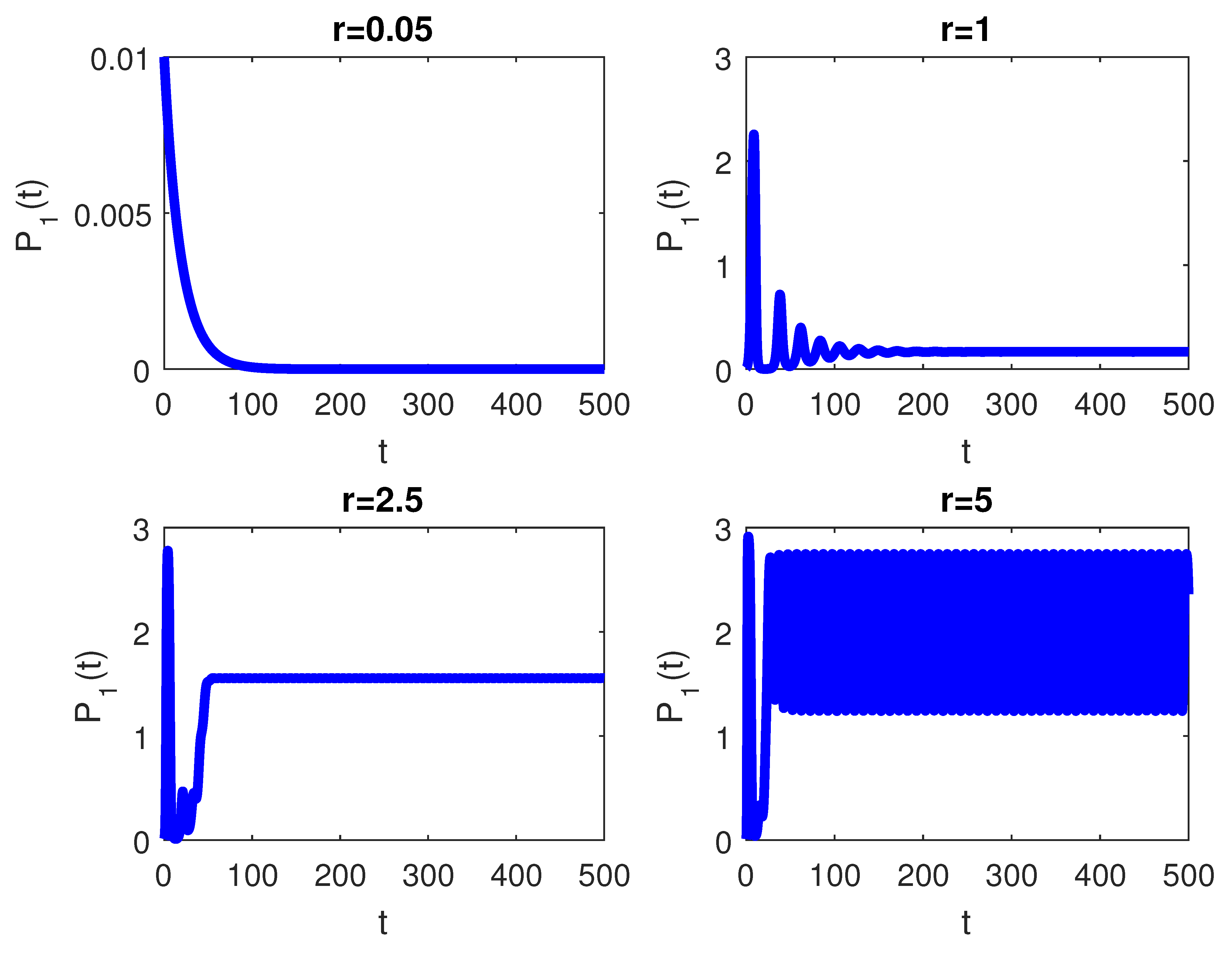
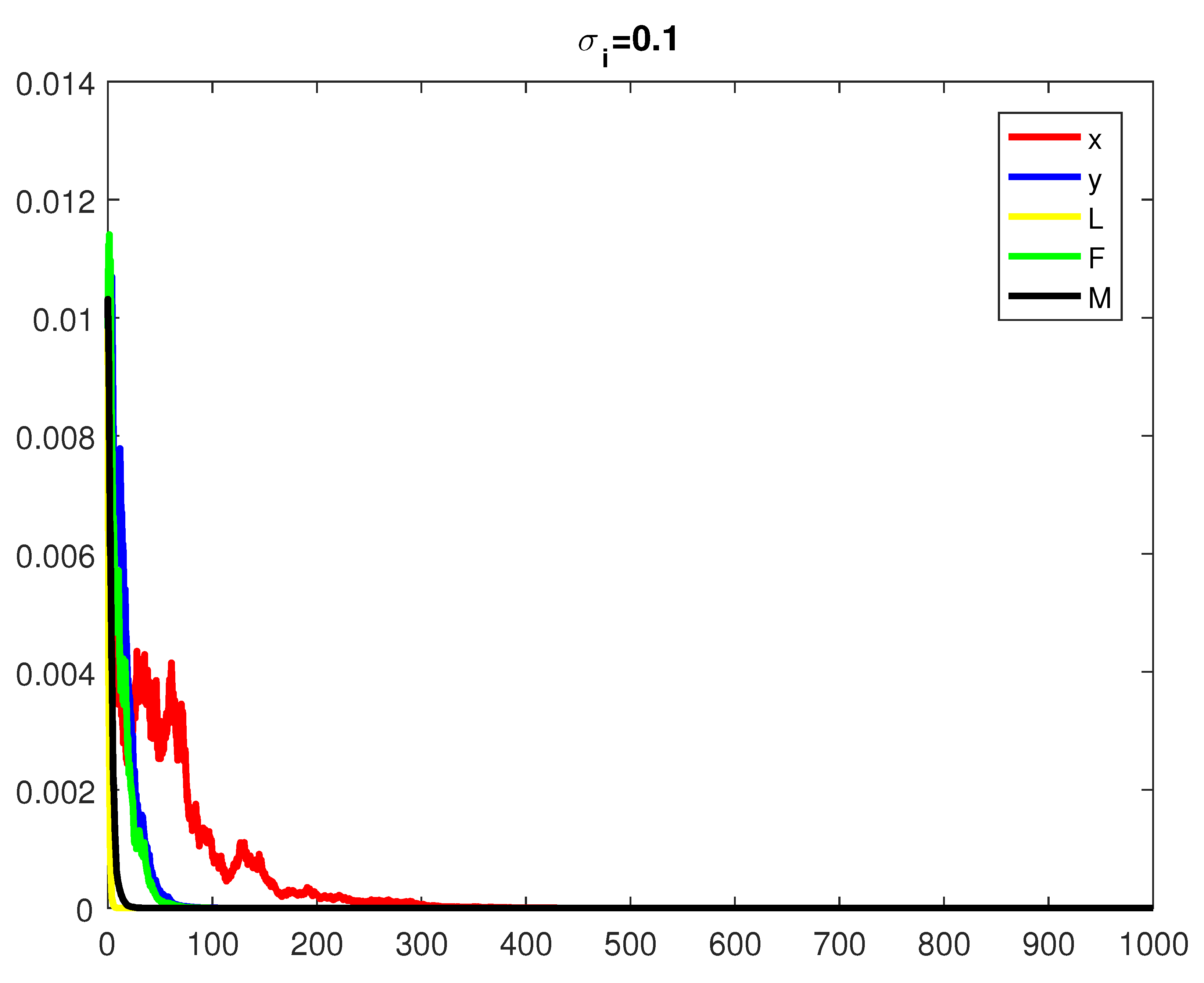
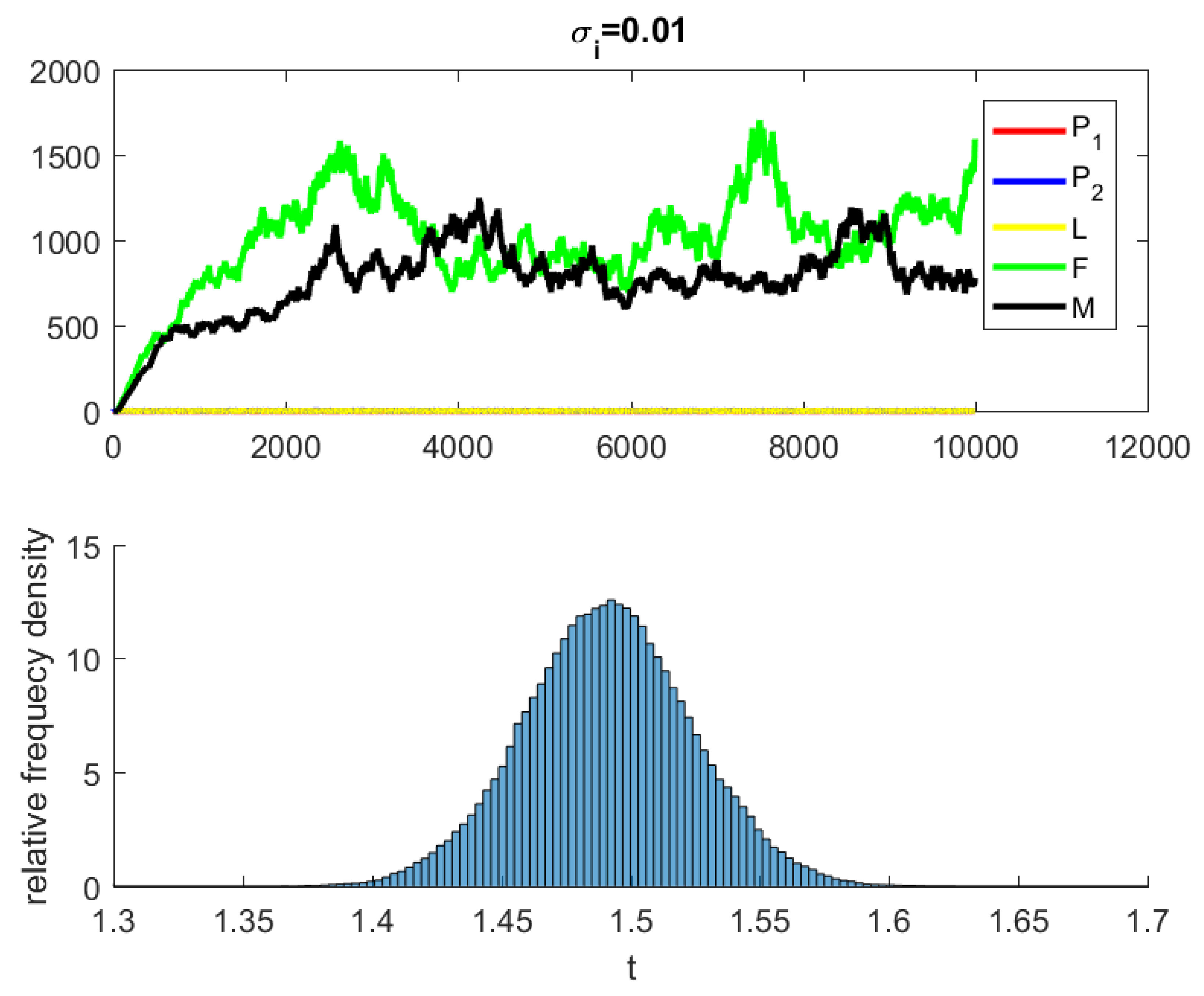
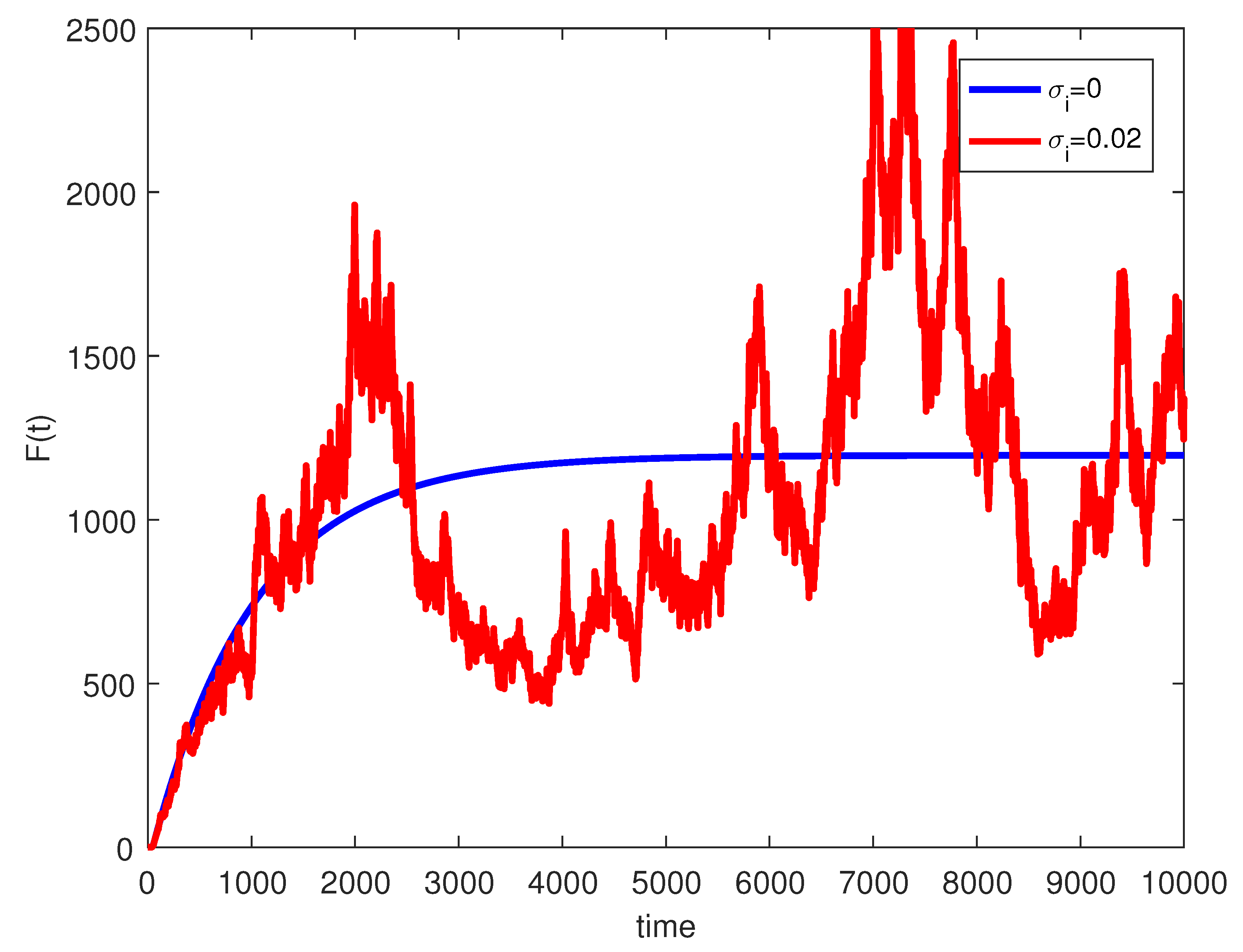
4. Numerical Simulations
5. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abdel-Baky, N.F.; Hamed, K.E.; Al-Otaibi, N.D.; Aldeghairi, M.A. Bioassay of Some Indigenous Entomopathogens for Controlling Rhynchophorus ferrugineus, Olivier in Saudi Arabia. Pak. J. Biol. Sci. 2021, 24, 944–952. [Google Scholar] [CrossRef]
- Wang, G.; Hou, Y.; Zhang, X.; Zhang, J.; Li, J.; Chen, Z. Strong population genetic structure of an invasive species, Rhynchophorus ferrugineus (Olivier), in southern China. Ecol. Evol. 2017, 7, 10770–10781. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Wahab, A.; Abd El-Fattah, A.; El-Shafei, W.; El Helaly, A. Efficacy of aggregation nano gel pheromone traps on the catchability of Rhynchophorus ferrugineus (Olivier) in Egypt. Braz. J. Biol. 2020, 81, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Faleiro, J.; Abdallah, A.B.; El-Bellaj, M.; Al-Ajlan, A.; Oihabi, A. Threat of the red palm weevil, Rhynchophorus ferrugineus (Olivier) to date palm plantations in North Africa. Arab. J. Plant Prot. 2012, 30, 274–280. [Google Scholar]
- Nangai, V.L.; Martin, B. Interpreting the acoustic characteristics of RPW towards its detection-A review. In Proceedings of the IOP Conference Series: Materials Science and Engineering, Narsimha Reddy Engineering College, Hyderabad, India, 3–4 July 2017; Volume 225, p. 012178. [Google Scholar]
- Al-Dosary, N.M.; Al-Dobai, S.; Faleiro, J.R. Review on the management of red palm weevil Rhynchophorus ferrugineus Olivier in date palm Phoenix dactylifera L. Emir. J. Food Agric. 2016, 28, 34–44. [Google Scholar] [CrossRef]
- Archer, L.; Crane, J.H.; Albrecht, U. Trunk Injection as a Tool to Deliver Plant Protection Materials—An Overview of Basic Principles and Practical Considerations. Horticulturae 2022, 8, 552. [Google Scholar] [CrossRef]
- El-Shafie, H.A.F.; Faleiro, J.R. Red palm weevil Rhynchophorus ferrugineus (Coleoptera: Curculionidae): Global invasion, current management options, challenges and future prospects. In Invasive Species-Introduction Pathways, Economic Impact, and Possible Management Options; 2020; Available online: https://doi.org/10.5772/intechopen.93391 (accessed on 6 July 2022).
- Ahmad, I. Integrated Pest Management of Rhynchophorus ferrugineus Olivier: An Efficient Approach to Reduce Infestation in Date Palm Trees. Pak. J. Zool. 2021, 54, 927. [Google Scholar] [CrossRef]
- Chihaoui-Meridja, S.; Harbi, A.; Abbes, K.; Chaabane, H.; La Pergola, A.; Chermiti, B.; Suma, P. Systematicity, persistence and efficacy of selected insecticides used in endotherapy to control the red palm weevil Rhynchophorus ferrugineus (Olivier, 1790) on Phoenix canariensis. Phytoparasitica 2020, 48, 75–85. [Google Scholar] [CrossRef]
- Faleiro, J. A review of the issues and management of the red palm weevil Rhynchophorus ferrugineus (Coleoptera: Rhynchophoridae) in coconut and date palm during the last one hundred years. Int. J. Trop. Insect Sci. 2006, 26, 135–154. [Google Scholar]
- Nurashikin-Khairuddin, W.; Abdul-Hamid, S.N.A.; Mansor, M.S.; Bharudin, I.; Othman, Z.; Jalinas, J. A Review of Entomopathogenic Nematodes as a Biological Control Agent for Red Palm Weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae). Insects 2022, 13, 245. [Google Scholar] [CrossRef]
- Vacas, S.; Melita, O.; Michaelakis, A.; Milonas, P.; Minuz, R.; Riolo, P.; Abbass, M.K.; Lo Bue, P.; Colazza, S.; Peri, E.; et al. Lures for red palm weevil trapping systems: Aggregation pheromone and synthetic kairomone. Pest Manag. Sci. 2017, 73, 223–231. [Google Scholar] [CrossRef]
- Soomro, M.H.; Mari, J.M.; Nizamani, I.A.; Gilal, A.A. Impact of trapping density on the performance of aggregation pheromone against Rhynchophorus ferrugineus (Coleoptera: Dryophthoridae). Int. J. Entomol Res. 2020, 5, 129–132. [Google Scholar]
- Shehab, B.; Ammr, M. The Influence of Prey and Predator Density on THE Predacious Behavior of Chrysoperla Mutata (Maclachlan) Larvae Feeding on Dubas Nymphs Ommatissus lybicus Deberg. Iraqi J. Sci. 2008, 49, 40–49. [Google Scholar]
- Al-Rawy, M.A.; Hamad, B.S.; Abdullatif, A.M. Factors affecting the effectiveness of Chrysoperla mutata (McL.) larvae feeding on dubas nymphs Ommatissus lybicus DeBerg. J. Educ. Sci. Stud. 2013, 1, 387–398. [Google Scholar]
- Barclay, H.J.; Jorge, H. Models for assessing the male annihilation of Bactrocera spp. with methyl eugenol baits. Ann. Entomol. Soc. Am. 2014, 107, 81–96. [Google Scholar] [CrossRef] [Green Version]
- Anguelov, R.; Dufourd, C.; Dumont, Y. Mathematical model for pest–insect control using mating disruption and trapping. Appl. Math. Model. 2017, 52, 437–457. [Google Scholar] [CrossRef] [Green Version]
- Ntahomvukiye, J.P.; Temgoua, A.; Bowong, S. Study of the population dynamics of Busseola fusca, maize pest. Acta Biotheor. 2018, 66, 379–397. [Google Scholar] [CrossRef]
- Xiang, S.; Pei, Y.; Liang, X. Analysis and optimization based on a sex pheromone and pesticide pest model with gestation delay. Int. J. Biomath. 2019, 12, 1950054. [Google Scholar] [CrossRef]
- Tapi, M.D.; Bagny-Beilhe, L.; Dumont, Y. Miridae control using sex-pheromone traps Modeling, analysis and simulations. Nonlinear Anal. Real World Appl. 2020, 54, 103082. [Google Scholar] [CrossRef]
- Alnafisah, Y.; El-Shahed, M. Deterministic and Stochastic Prey–Predator Model for Three Predators and a Single Prey. Axioms 2022, 11, 156. [Google Scholar] [CrossRef]
- Xu, C.; Ren, G.; Yu, Y. Extinction analysis of stochastic predator–prey system with stage structure and crowley–martin functional response. Entropy 2019, 21, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, G. Dynamics of a stochastic population model with predation effects in polluted environments. Adv. Differ. Equ. 2021, 2021, 189. [Google Scholar] [CrossRef]
- Mu, Y.; Lo, W.C. Stochastic dynamics of populations with refuge in polluted turbidostat. Chaos Solitons Fractals 2021, 147, 110963. [Google Scholar] [CrossRef]
- Wang, Z.; Deng, M.; Liu, M. Stationary distribution of a stochastic ratio-dependent predator-prey system with regime-switching. Chaos Solitons Fractals 2021, 142, 110462. [Google Scholar] [CrossRef]
- Salman, S.; Yousef, A.; Elsadany, A. Dynamic behavior and bifurcation analysis of a deterministic and stochastic coupled logistic map system. Int. J. Dyn. Control 2022, 10, 69–85. [Google Scholar] [CrossRef]
- Liu, Q.; Jiang, D.; Hayat, T.; Alsaedi, A.; Ahmad, B. A stochastic SIRS epidemic model with logistic growth and general nonlinear incidence rate. Phys. Stat. Mech. Its Appl. 2020, 551, 124152. [Google Scholar] [CrossRef]
- Li, Q.; Cong, F.; Liu, T.; Zhou, Y. Stationary distribution of a stochastic HIV model with two infective stages. Phys. Stat. Mech. Its Appl. 2020, 554, 124686. [Google Scholar] [CrossRef]
- Khan, A.; Hussain, G.; Yusuf, A.; Usman, A.H.; Humphries, U.W. A hepatitis stochastic epidemic model with acute and chronic stages. Adv. Differ. Equ. 2021, 2021, 181. [Google Scholar] [CrossRef]
- Wang, X.; Wang, C.; Wang, K. Extinction and persistence of a stochastic SICA epidemic model with standard incidence rate for HIV transmission. Adv. Differ. Equ. 2021, 2021, 1–17. [Google Scholar] [CrossRef]
- Wang, X.; Tan, Y.; Cai, Y.; Wang, K.; Wang, W. Dynamics of a stochastic HBV infection model with cell-to-cell transmission and immune response. Math. Biosci. Eng. 2021, 18, 616–642. [Google Scholar] [CrossRef]
- Lan, G.; Yuan, S.; Song, B. The impact of hospital resources and environmental perturbations to the dynamics of SIRS model. J. Frankl. Inst. 2021, 358, 2405–2433. [Google Scholar] [CrossRef]
- Ikram, R.; Khan, A.; Zahri, M.; Saeed, A.; Yavuz, M.; Kumam, P. Extinction and stationary distribution of a stochastic COVID-19 epidemic model with time-delay. Comput. Biol. Med. 2022, 141, 105115. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Liu, J.; Zhang, S. Analysis of a stochastic eco-epidemiological model with modified Leslie–Gower functional response. Adv. Differ. Equ. 2018, 2018, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Mao, X. Stochastic Differential Equations and Applications; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Li, L.; Zhao, W. Deterministic and stochastic dynamics of a modified Leslie-Gower prey-predator system with simplified Holling-type IV scheme. Math. Biosci. Eng. 2021, 18, 2813–2831. [Google Scholar] [CrossRef]
- Li, J.; Shan, M.; Banerjee, M.; Wang, W. Stochastic dynamics of feline immunodeficiency virus within cat populations. J. Frankl. Inst. 2016, 353, 4191–4212. [Google Scholar] [CrossRef] [Green Version]
- Arnold, L. Stochastic Differential Equations; Wiley-Interscience: New York, NY, USA, 1974; p. 243. [Google Scholar]
- Khasminskii, R. Stochastic Stability of Differential Equations; Springer: Berlin/Heidelberg, Germany, 2011; Volume 66. [Google Scholar]
- Caraballo, T.; Kloeden, P.E. The persistence of synchronization under environmental noise. Proc. R. Soc. A Math. Phys. Eng. Sci. 2005, 461, 2257–2267. [Google Scholar] [CrossRef]
- Ji, C.; Jiang, D.; Liu, H.; Yang, Q. Existence, uniqueness and ergodicity of positive solution of mutualism system with stochastic perturbation. Math. Probl. Eng. 2010, 2010, 684926. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhao, W. Dynamical analysis of a stochastic SIRS epidemic model with saturating contact rate. Math. Biosci. Eng. 2020, 17, 5925–5943. [Google Scholar] [CrossRef]
- Huang, Y.; Shi, W.; Wei, C.; Zhang, S. A stochastic predator–prey model with Holling II increasing function in the predator. J. Biol. Dyn. 2021, 15, 1–18. [Google Scholar] [CrossRef]
- Higham, D.J. An algorithmic introduction to numerical simulation of stochastic differential equations. SIAM Rev. 2001, 43, 525–546. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Shahed, M.; Al-Nujiban, A.; Abdel-Baky, N.F. Stochastic Modelling of Red Palm Weevil Using Chemical Injection and Pheromone Traps. Axioms 2022, 11, 334. https://doi.org/10.3390/axioms11070334
El-Shahed M, Al-Nujiban A, Abdel-Baky NF. Stochastic Modelling of Red Palm Weevil Using Chemical Injection and Pheromone Traps. Axioms. 2022; 11(7):334. https://doi.org/10.3390/axioms11070334
Chicago/Turabian StyleEl-Shahed, Moustafa, Asma Al-Nujiban, and Nagdy F. Abdel-Baky. 2022. "Stochastic Modelling of Red Palm Weevil Using Chemical Injection and Pheromone Traps" Axioms 11, no. 7: 334. https://doi.org/10.3390/axioms11070334