

Bioleaching of Chalcopyrite by a New Strain Leptospirillum ferrodiazotrophum Ksh-L Isolated from a Dump-Bioleaching System of Kashen Copper-Molybdenum Mine
 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Conditions and Isolation
2.2. Morphology Studies
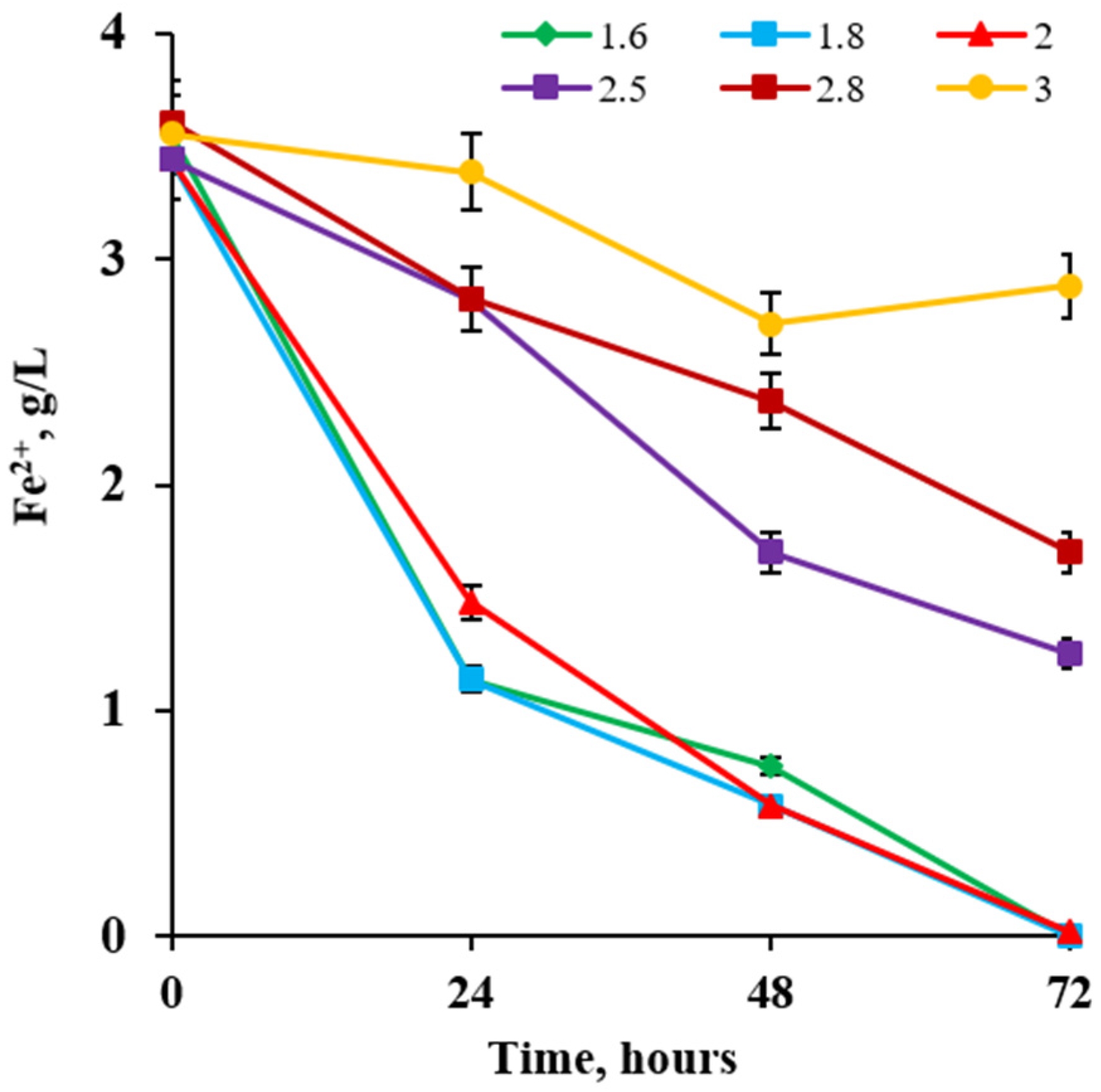
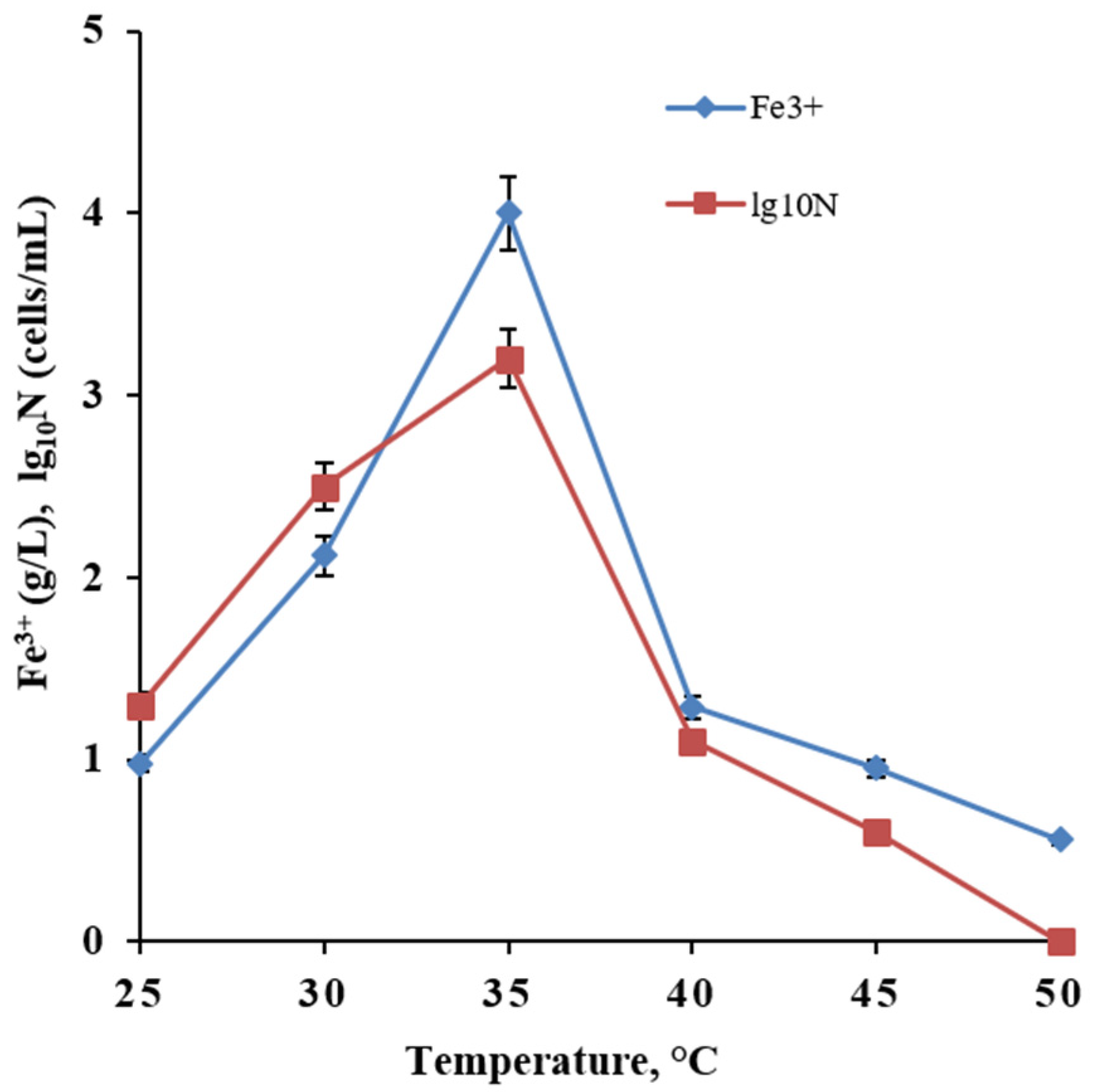
2.3. Optimal pH and Temperature for Growth
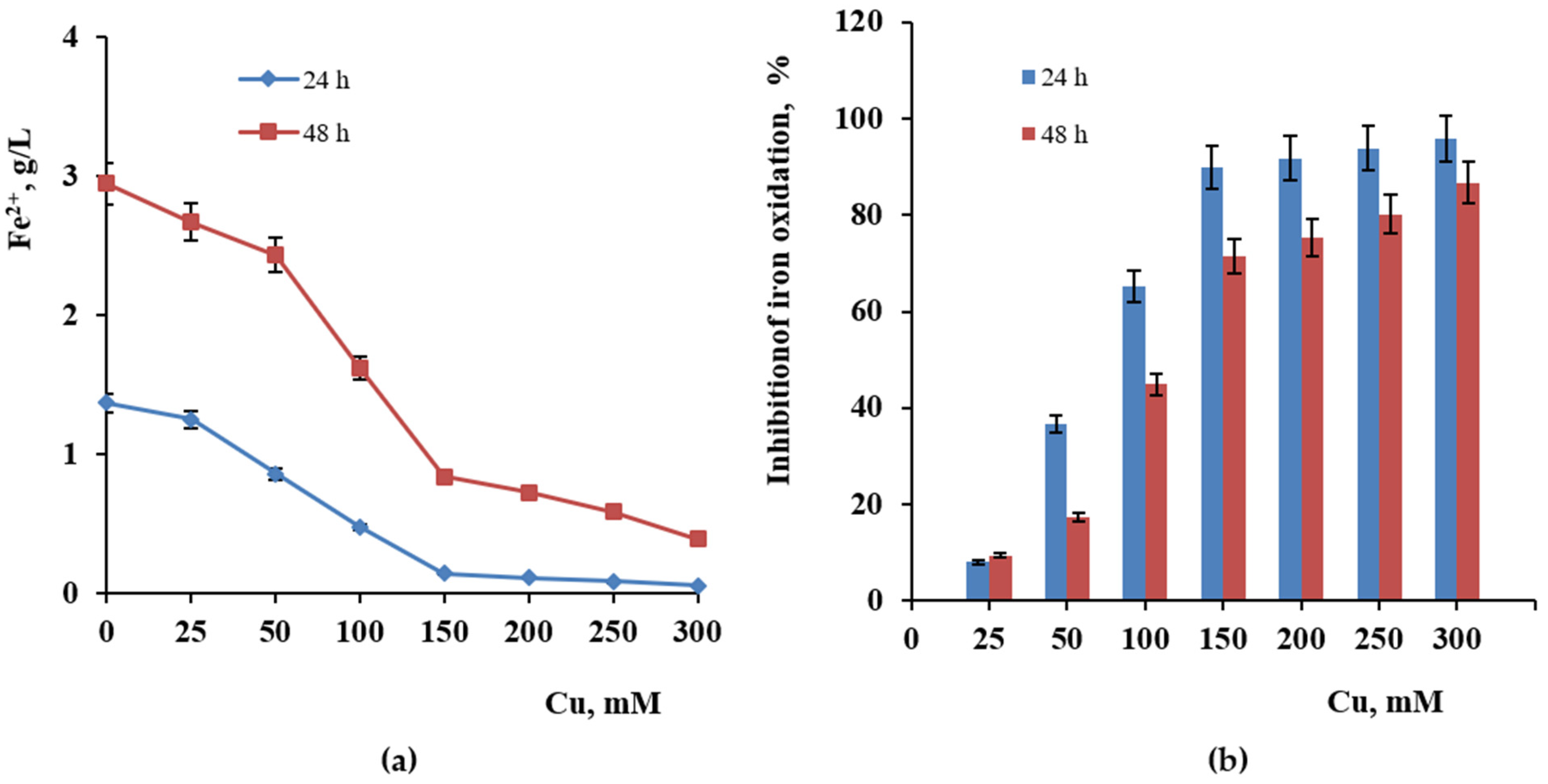
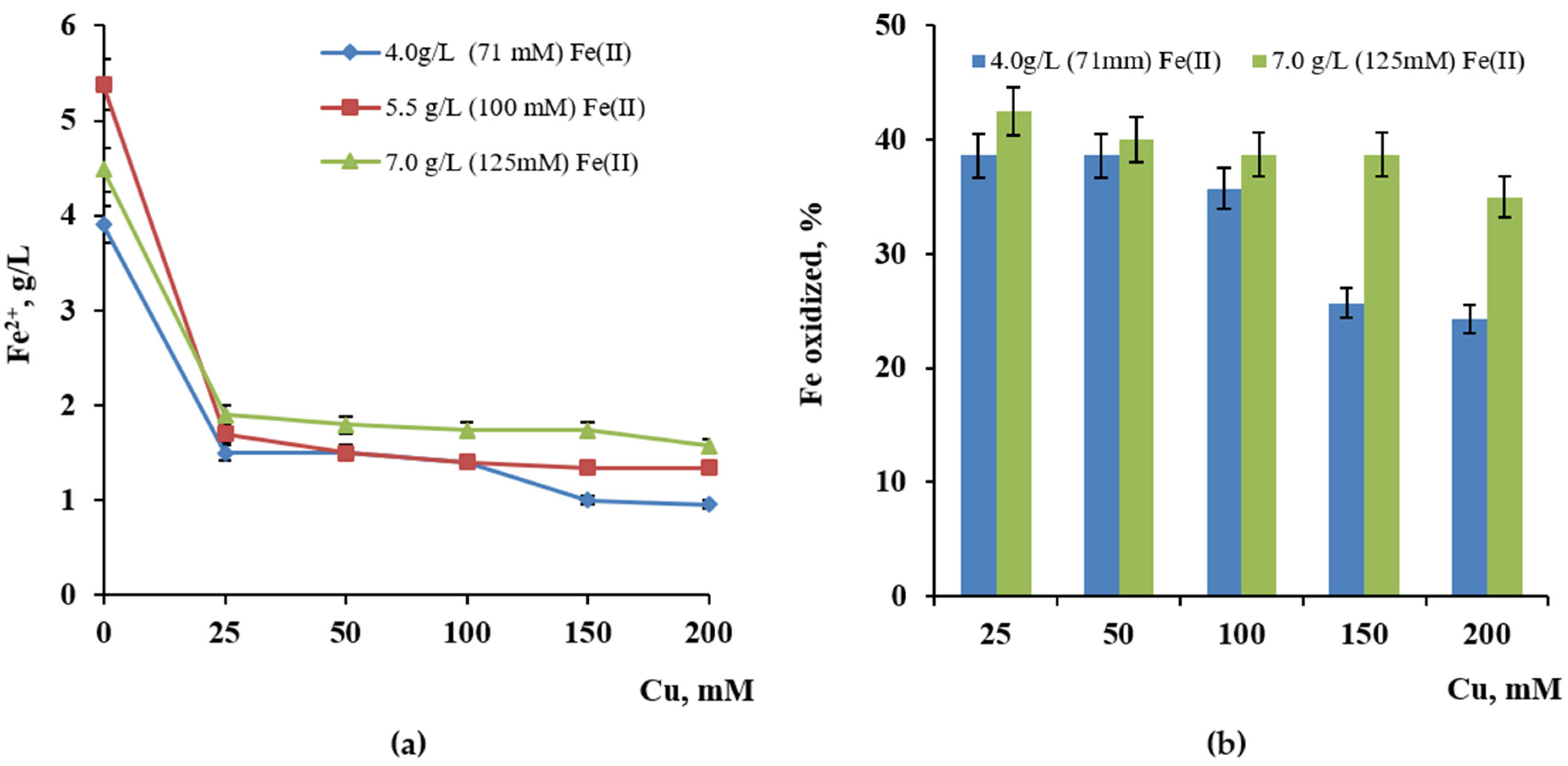
2.4. Influence of Copper and Zinc
2.5. EPS Analysis, Extraction, and Determination
2.5.1. EPS Extraction
2.5.2. Determination of EPS Composition
2.6. DNA Extraction, PCR of 16s rRNA, Sequencing, and Phylogenetic Analysis
2.6.1. DNA Extraction
2.6.2. Extraction and Purification of PCR Product
2.6.3. Construction of Phylogenetic Tree
2.7. Leaching Experiments
3. Results
3.1. Isolation of Strain Ksh-L
3.2. Morphology
3.3. Optimal pH and Temperature for Growth
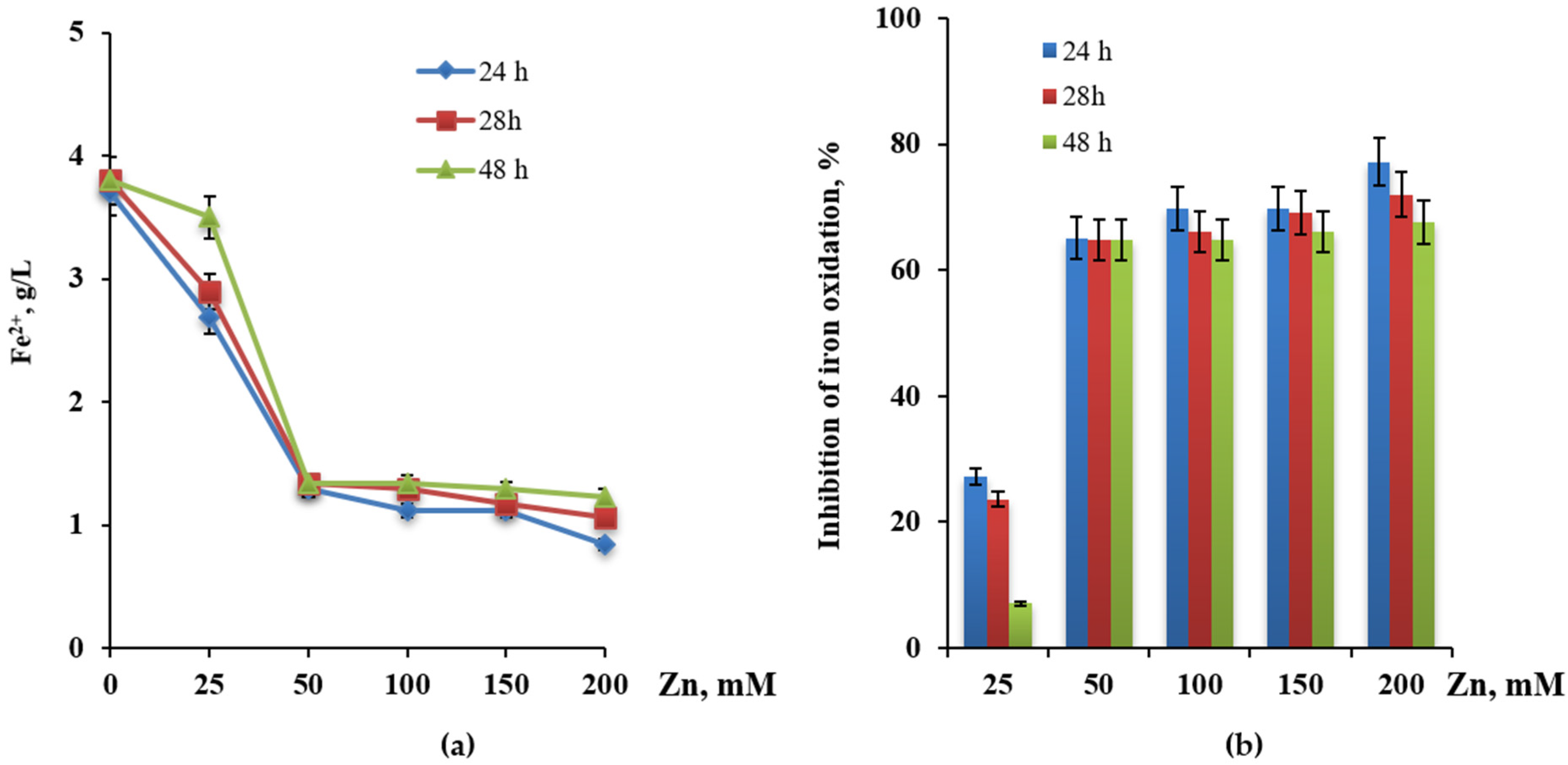
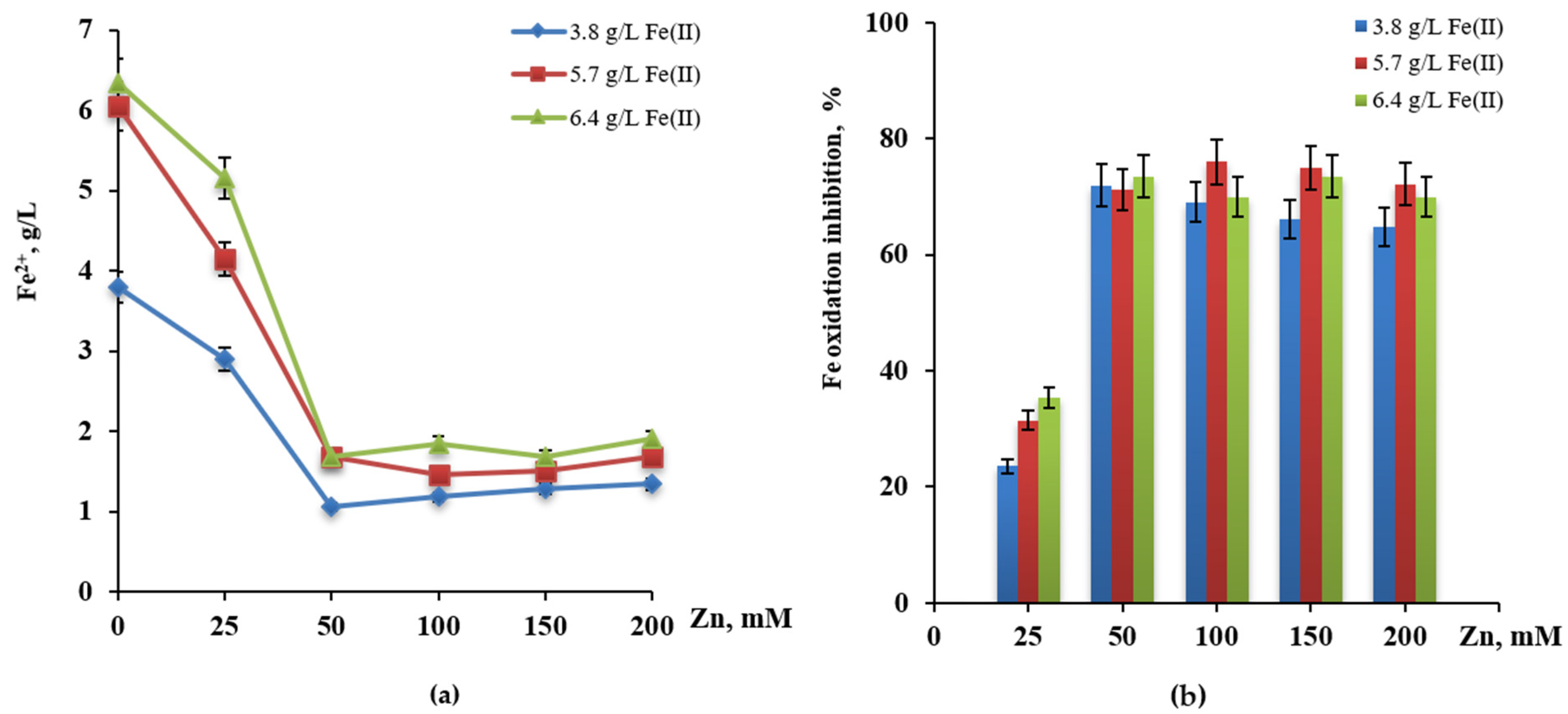
3.4. Influence of Copper and Zinc
3.5. Adaptation of L. ferrodiazotrophum Ksh-L
3.6. EPS Analysis
3.7. Phylogenetic Analysis of 16S rRNA
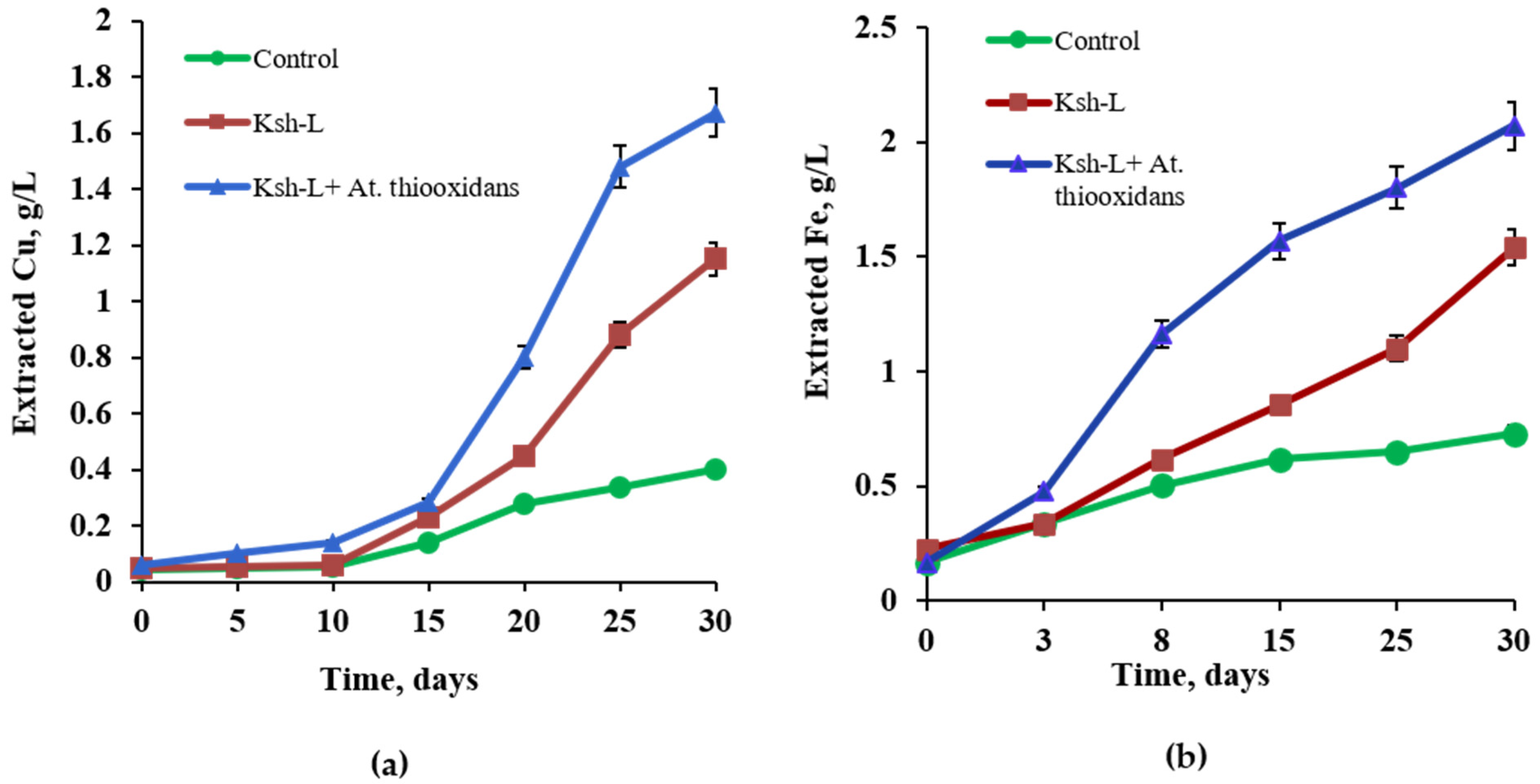
3.8. Bioleaching of Chalcopyrite
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, H.; Zhang, Y.; Zhang, X.; Qian, L.; Sun, M.; Yang, Y.; Zhang, Y.; Wang, J.; Kim, H.; Qiu, G. The dissolution and passivation mechanism of chalcopyrite in bioleaching: An overview. Miner. Eng. 2019, 136, 140–154. [Google Scholar] [CrossRef]
- Jones, S.; Santini, J.M. Mechanisms of bioleaching: Iron and sulfur oxidation by acidophilic microorganisms. Essays Biochem. 2023, 67, 685–699. [Google Scholar] [CrossRef]
- Rohwerder, T.; Gehrke, T.; Kinzler, K.; Sand, W. Bioleaching review part A: Progress in bioleaching: Fundamentals and mechanisms of bacterial metal sulfide oxidation. Appl. Microbiol. Biotechnol. 2003, 63, 239–248. [Google Scholar] [CrossRef]
- Huang, C.; Qin, C.; Feng, X.; Liu, X.; Yin, H.; Jiang, L.; Liang, Y.; Liu, H.; Tao, J. Chalcopyrite bioleaching of an in situ leaching system by introducing different functional oxidizers. RSC Adv. 2018, 8, 37040. [Google Scholar] [CrossRef]
- Watling, H.R.; Watkin, E.L.; Ralphe, D.E. The resilience and versatility of acidophiles that contribute to the bio-assisted extraction of metals from mineral sulphides. Environ. Technol. 2010, 31, 915–933. [Google Scholar] [CrossRef]
- Spolaore, P.; Joulian, C.; Gouin, J.; Morin, D.; D’Hugues, P. Relationship between bioleaching performance, bacterial community structure and mineralogy in the bioleaching of a copper concentrate in stirred-tank reactors. Appl. Microb. Biotechnol. 2011, 89, 441–448. [Google Scholar] [CrossRef]
- Ghorbani, Y.; Franzidis, J.P.; Petersen, J. Heap leaching technology—Current State, innovations, and future directions: A review. Miner. Process. Extr. Metall. Rev. 2016, 37, 73–119. [Google Scholar] [CrossRef]
- Veloso, T.C.; Peixoto, J.J.M.; Pereira, M.S.; Leao, V.A. Kinetics of chalcopyrite leaching in either ferric sulphate or cupric sulphate media in the presence of NaCl. Int. J. Miner. Process 2016, 148, 147–154. [Google Scholar] [CrossRef]
- Rawlings, D.E.; Johnson, D.B. The microbiology of biomining: Development and optimization of mineral-oxidizing microbial consortia. Microbiology 2007, 153, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.Q.; Li, W.Z.; Lan, Z.Y.; Qiu, G.Z. Simulated small-scale pilot heap leaching of low-grade oxide zinc ore with integrated selective extraction of zinc. Miner. Eng. 2007, 20, 694–700. [Google Scholar] [CrossRef]
- Giaveno, A.; Lavalle, L.; Chiacchiarini, P.; Donati, E. Bioleaching of zinc from low-grade complex sulphide ores in an airlift by isolated Leptospirillum ferrooxidans. Hydrometallurgy 2007, 89, 117–126. [Google Scholar] [CrossRef]
- Gericke, M.; Muller, H.H.; van Staden, P.J.; Pinches, A. Development of a tank bioleaching process for the treatment of complex Cu-polymetallic concentrates. Hydrometallurgy 2008, 94, 23–28. [Google Scholar] [CrossRef]
- Patel, B.C.; Tippe, D.R.; Dave, S.R. Optimization of copper and bioregeneration under metallic stress. Hydrometallurgy 2012, 117–118, 18–23. [Google Scholar] [CrossRef]
- Pradhan, N.; Nathsarma, K.C.; Srinivasa Rao, K.; Sukla, L.B.; Mishra, B.K. Heap bioleaching of chalcopyrite: A review. Miner. Eng. 2008, 21, 355–365. [Google Scholar] [CrossRef]
- Abhilash; Mehta, K.D.; Pandey, B.D. Bacterial Leaching Kinetics for Copper Dissolution from a Low- Grade Indian Chalcopyrite Ore. Rev. Esc. Minas 2013, 66, 245–250. [Google Scholar] [CrossRef]
- Panda, S.; Akcil, A.; Pradhan, N.; Deveci, H. Current scenario of chalcopyrite bioleaching: A review on the recent advances to its heap-leach technology. Bioresour. Technol. 2015, 196, 694–706. [Google Scholar] [CrossRef]
- Lawson, E.N. The composition of mixed populations of leaching bacteria active in gold and nickel recovery from sulphide ores. In ternational Biohydrometallurgy Symposium. In Biomine 97; Australian Mineral Foundation: Glenside, Australia, 1997; pp. QP4.1–QP4.10. [Google Scholar]
- Rawlings, D.E.; Tributsch, H.; Hansford, G.S. Reasons why ‘Leptospirillum’-like species rather than Thiobacillus ferrooxidans are the dominant iron-oxidizing bacteria in many commercial processes for the biooxidation of pyrite and related ores. Microbiol. Read. 1999, 145, 5–13. [Google Scholar] [CrossRef]
- Rawlings, D.E.; Coram, N.J.; Gardner, M.N.; Deane, S.M. Thiobacillus caldus and Leptospirillum ferrooxians are widely distributed in continuous-flow biooxidation tanks used to treat a variety of metalcontaining ores and concentrates. In Biohydrometrallurgy and the Environment toward the Mining of the 21st Century, Part A; Amils, R., Balleste, A., Eds.; Elsevier Press: Amsterdam, The Netherlands, 1999; pp. 773–778. [Google Scholar]
- Coram, N.J.; Rawlings, D.E. Molecular relationship between two groups of the genus Leptospirillum and the finding that Leptospirillum ferriphilum sp. nov. dominates in South African commercial bio-oxidation tanks that operate at 40 °C. App Environ. Microbiol. 2002, 68, 838–845. [Google Scholar] [CrossRef]
- Schippers, A. Microorganisms involved in bioleaching and nucleic acid-based molecular methods for their identification and quantification. In Microbial Processing of Metal Sulfides; Donati, R.E., Sand, W., Eds.; Springer: New York, NY, USA, 2007; pp. 3–33. [Google Scholar]
- Dave, S.R. Selection of Leptospirillum ferrooxidans SRPCBL and development for enhanced ferric regeneration in stirred tank and airlift column reactor. Bioresour. Technol. 2008, 99, 7803–7806. [Google Scholar] [CrossRef]
- Markosyan, G.E. New iron oxidizing bacteria Leptospirillum ferrooxidans nov. gen. nov. sp. Biol. J. Armen. 1972, 25, 26–29. (In Russian) [Google Scholar]
- Vardanyan, N.; Akopyan, V. Leptospirillum-like bacteria and evaluation of their role in pyrite oxidation. Microbiology 2003, 72, 438–442. [Google Scholar] [CrossRef]
- Ram, R.J.; Verberkmoes, N.C.; Thelen, M.P.; Tyson, G.W.; Baker, B.J.; Blake, R.C., II; Shah, M.; Hettich, R.L.; Banfield, J.F. Community proteomics of a natural microbial biofilm. Science 2005, 308, 1915–1920. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.S.; Xie, X.H.; Xiao, S.M.; Wang, X.M.; Zhao, W.J.; Tian, Z.L. Isolation of Leptospirillum ferriphilum by single-layered solid medium. J. Centr South. Univ. Technol. 2007, 14, 467–473. [Google Scholar] [CrossRef]
- Bond, P.L.; Banfield, J.F. Design and performance of rRNA targeted oligonucleotide probes for in situ detection and phylogenetic identification of microorganisms inhabiting acid mine drainage environments. Microb. Ecol. 2001, 41, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Hippe, H. Leptospirillium gen. nov (ex Markoysan 1972), nom. rev., including Leptospirillium ferrooxidans sp. nov. (ex Markoysan 1972), nom. rev. and Leptospirillium thermoferrooxidans sp. nov. (Golovacheva et al. 1992). Int. J. Syst. Evol. Microbiol. 2000, 50, 501–503. [Google Scholar] [CrossRef] [PubMed]
- Bond, P.L.; Smriga, S.P.; Banfield, J.F. Phylogeny of microorganisms populating a thick, subaerial predominantly lithitrophic biofilm at an extreme acid mine drainage site. Appl. Environ. Microbiol. 2000, 66, 3842–3849. [Google Scholar] [CrossRef]
- Tyson, G.W.; Chapman, J.; Hugenholtz, P.; Allen, E.E.; Ram, R.J.; Richardson, P.M.; Solovyev, V.V.; Rubin, E.M.; Rokhsar, D.S.; Banfield, J.F. Community structure and metabolism through reconstruction of microbial genomes from the environment. Nature 2004, 428, 37–43. [Google Scholar] [CrossRef]
- Tyson, G.W.; Lo, I.; Baker, B.J.; Allen, E.E.; Hugenholtz, P.; Banfield, J.F. Genome-directed isolation of the key nitrogen fixer Leptospirillum ferrodiazotrophum sp. nov. from an acidophilic microbial community. Appl. Environ. Microbiol. 2005, 71, 6319–6324. [Google Scholar] [CrossRef]
- Allen, E.E.; Tyson, G.W.; Whitaker, R.J.; Detter, J.C.; Richardson, P.M.; Banfield, J.F. Genome dynamics in a natural archaeal population. Proc. Natl. Acad. Sci. USA 2007, 104, 1883–1888. [Google Scholar] [CrossRef]
- Goltsman, D.S.; Denef, V.J.; Singer, S.W.; VerBerkmoes, N.C.; Lefsrud, M.; Mueller, R.S.; Dick, G.J.; Sun, C.L.; Wheeler, K.E.; Zemla, A.; et al. Community genomic and proteomic analyses of chemoautotrophic iron-oxidizing “Leptospirillum rubarum” (group II) and “Leptospirillum ferrodiazotrophum” (group III) bacteria in acid mine drainage biofilms. Appl. Environ. Microbiol. 2009, 75, 4599–4615. [Google Scholar] [CrossRef]
- Wilmes, P.; Remis, J.P.; Hwang, M.; Auer, M.; Thelen, M.P.; Banfield, J.F. Natural acidophilic biofilm communities reflect distinct organismal and functional organization. ISME J. 2009, 3, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Goltsman, A.D.S.; Dasari, M.; Thomas, B.C.; Shah, M.B.; VerBerkmoes, N.C.; Hettich, R.L.; Banfield, J.F. New group in the Leptospirillum clade: Cultivation-independent community genomics, proteomics, and transcriptomics of the new species “Leptospirillum group IV UBA BS”. Appl. Environ. Microbiol. 2013, 79, 5384–5393. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Dopson, M. Life in acid: pH homeostasis in acidophiles. Trends Microbiol. 2007, 15, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Slonczewski, J.L.; Fujisawa, M.; Dopson, M.; Krulwich, T.A. Cytoplasmic pH measurement and homeostasis in bacteria and archaea. Adv. Microb. Physiol. 2009, 55, 1–79. [Google Scholar] [PubMed]
- Rensing, C.; Grass, G. Escherichia coli mechanism of copper homeostasis in a changing environment. FEMS Microbiol. Rev. 2003, 27, 197–213. [Google Scholar] [CrossRef] [PubMed]
- Outten, F.W.; Huffman, D.L.; Hale, J.A.; Ó Halloran, T.V. The independent cue and cus system confer copper tolerance during aerobic and anaerobic growth in Escherichia coli. J. Biol. Chem. 2001, 276, 30670–30677. [Google Scholar] [CrossRef]
- Puig, S.; Thiele, D.J. Molecular mechanisms of copper uptake and distribution. Curr. Opin. Chem. Biol. 2002, 6, 171–180. [Google Scholar] [CrossRef]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial biofilms. Ann. Rev. Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef]
- Sutherland, I.W. The biofilm matrix--an immobilized but dynamic microbial environment. Trends Microbiol. 2001, 9, 222–227. [Google Scholar] [CrossRef]
- Harneit, K.; Goksel, A.; Kock, D.; Klock, J.H.; Gehrke, T.; Sand, W. Adhesion to metal sulfide surfaces by cells of Acidithiobacillus ferrooxidans, Acidithiobacillus thiooxidans and Leptospirillum ferrooxidans. Hydrometallurgy 2006, 83, 245–254. [Google Scholar] [CrossRef]
- Teitzel, G.M.; Parsek, M.R. Heavy metal resistance of biofilm and planktonic Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2003, 69, 2313–2320. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.J.; Ceri, H.; Turner, R.J. Multimetal resistance and tolerance in microbial biofilms. Nat. Rev. Microbiol. 2007, 5, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Yong, N.K.; Oshima, M.; Blake, R.C.; Sugio, T. Isolation and some properties of an iron-oxidizing bacterium Thiobacillus ferrooxidans resistant to molybdenum ion. Biosci. Biotechnol. Biochem. 1997, 61, 1523–1526. [Google Scholar] [CrossRef]
- Valdés, J.; Pedroso, I.; Quatrini, R.; Dodson, R.J.; Tettelin, H.; Blake, R., II; Eisen, J.A.; Holmes, D.S. Acidithiobacillus ferrooxidans metabolism: From genome sequence to industrial applications. BMC Genom. 2008, 9, 597. [Google Scholar]
- Mi, S.; Song, J.; Lin, J.; Che, Y.; Zheng, H.; Lin, J. Complete genome of Leptospirillum ferriphilum ML-04 provides insight into its physiology and environmental adaptation. J. Microbiol. 2011, 49, 890–901. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas, J.P.; Lazcano, M.; Ossandon, F.J.; Corbett, M.; Holmes, D.S.; Watkin, E. Draft genome sequence of the iron-oxidizing acidophile Leptospirillum ferriphilum type strain DSM 14647. Genome Announc 2014, 2, e01153-14. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Liang, Y.; Yin, H.; Xiao, Y.; Guo, X.; Xu, Y.; Hu, Q.; Liu, H.; Liu, X. Effects of arsenite resistance on the growth and functional gene expression of Leptospirillum ferriphilum and Acidithiobacillus thiooxidans in pure culture and coculture. Biomed. Res. Int. 2015, 2015, 203197. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, L.; Chi, A.; Beard, S.; Orell, A.; Guiliani, N.; Shabanowitz, J.; Hunt, D.F.; Jerez, C.A. Genomics, metagenomics and proteomics in biomining microorganisms. Biotechnol. Adv. 2006, 24, 197–211. [Google Scholar] [CrossRef]
- Moreno-Paz, M.; Gómez, M.J.; Arcas, A.; Parro, V. Environmental transcriptome analysis reveals physiological differences between biofilm and planktonic modes of life of the iron oxidizing bacteria Leptospirillum spp. in their natural microbial community. BMC Genom. 2010, 11, 404. [Google Scholar] [CrossRef]
- Mangold, S.; Potrykus, J.; Björn, E.; Lövgren, L.; Dopson, M. Extreme zinc tolerance in acidophilic microorganisms from the bacterial and archaeal domains. Extremophiles 2013, 17, 75–85. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, X.; Liu, D.; Duan, H.; Fan, H. Sequencing and bioinformatics analysis of the metal-related genes in Acidithiobacillus ferrooxidans strain DC. Folia Microbiol. 2013, 58, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Watling, H.R. The bioleaching of sulphide minerals with emphasis on copper sulphides—A review. Hydrometallurgy 2006, 84, 81–108. [Google Scholar] [CrossRef]
- Orell, A.; Navarro, C.A.; Arancibia, R.; Mobarec, J.C.; Jerez, C.A. Life in blue: Copper resistance mechanisms of bacteria and Archaea used in industrial biomining of minerals. Biotechnol. Adv. 2010, 28, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Orell, A.; Navarro, C.A.; Rivero, M.; Aguilar, J.S.; Jerez, C.A. Inorganic polyphosphates in extremophiles and their possible functions. Extremophiles 2012, 16, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Silverman, M.P.; Lundgren, D.G. Studies on the chemoautotrophic iron bacterium Ferrobacillus ferrooxidans. I. An improved medium and a harvesting procedure for securing high cell yields. J. Bacteriol. 1959, 77, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, P.; Murrayn, R.G.E.; Costilow, R.N.; Nestar, E.W.; Wood, W.A.; Krieg, N.R.; Phillips, G.B. Manual of Methods for General Bacteriology. American Society of Microbiology: Washington, DC, USA, 1981. [Google Scholar]
- Mackintosh, M.E. Nitrogen fixation by Thiobacillus ferrooxidans. J. Gen. Microbiol. 1987, 105, 215–218. [Google Scholar] [CrossRef]
- Castro, L.; Zhang, R.; Munoz, J.A.; Gonzalez, F.; Blazquez, M.L.; Sand, W. Characterization of exopolymeric substances (EPS) produced by Aeromonas hydrophila under reducing conditions. Biofouling 2014, 30, 501–511. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1996, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 84, 5463–5467. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Lucchesi, C.A.; Hirn, C.F. EDTA Titration of total Iron in Iron(II) and Iron(III) mixtures. Application to Iron driers. Anal. Chem. 1960, 32, 1191–1193. [Google Scholar] [CrossRef]
- Zhang, R.Y.; Xia, J.L.; Peng, J.H.; Zhang, Q.; Zhang, C.G.; Nie, Z.Y.; Qui, G.Z. A new strain Leptospirillum ferriphilum YTW315 for bioleaching of metal sulfides ores. Trans. Nonferrous Met. Soc. China 2010, 20, 135–141. [Google Scholar] [CrossRef]
- Valix, M.; Loon, L. Adaptive tolerance behaviour of fungi in heavy metals. Miner. Eng. 2003, 16, 193–198. [Google Scholar] [CrossRef]
- Vartanyan, N.S.; Karavaiko, G.I.; Pivovarova, T.A.; Dorofeev, A.G. Resistance of Sulfobacillus thermosulfidooxidans subspecies asporogenes to Cu2+, Zn2+ and Ni2+ ions. Microbiology 1990, 59, 399–404, (English translation of Mikrobiologiya). [Google Scholar]
- Sand, W.; Gehrke, T. Analysis and function of the EPS from the strong acidophile Thiobacillus ferrooxidans. In Microbial Extracellular Polymeric Substances; Wingender, J., Neu, T.R., Flemming, H.C., Eds.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 127–140. [Google Scholar]
- Sheng, G.P.; Yu, H.Q.; Li, X.Y. Extracellular polymeric substances (EPS) of microbial aggregates in biological wastewater treatment systems: A review. Biotechnol. Adv. 2010, 28, 882–894. [Google Scholar] [CrossRef]
- Vardanyan, A.; Vardanyan, N.; Markosyan, L.; Sand, W.; Vera, M.; Zhang, R. Biofilm formation and extracellular polymeric substances (EPS) analysis by new isolates of Leptospirillum, Acidithiobacillus and Sulfobacillus from Armenia. Adv. Mater. Res. 2015, 1130, 153–156. [Google Scholar] [CrossRef]
- Vardanyan, A.; Vardanyan, N.; Khachatryan, A.; Zhang, R.; Sand, W. Adhesion to mineral surfaces by cells of Leptospirillum, Acidithiobacillus and Sulfobacillus from Armenian sulfide ores. Minerals 2019, 9, 69. [Google Scholar] [CrossRef]
- Vardanyan, N.; Badalyan, H.; Markosyan, L.; Vardanyan, A.; Zhang, R.; Sand, W. Newly isolated Acidithiobacillus sp. Ksh from Kashen copper ore: Peculiarities of EPS and colloidal exopolysaccharide. Front. Microbiol. 2020, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Cao, L.; Qiu, G.; Wang, D.; Kellogg, L.; Zhou, J.; Liu, X.; Dai, Z.; Ding, J.; Liu, X. Molecular diversity of 16S rRNA and gyrB genes in copper mines. Arch. Microbiol. 2008, 189, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Corbett, M.K. Leptospirillum: A Study of the Nitrogen Fixing Capabilities. Ph.D. Thesis, Philosophy of Curtin University, Perth, Australia, 2011; pp. 1–202. Available online: https://espace.curtin.edu.au/handle/20.500.11937/81 (accessed on 31 December 2011).
- Hallberg, K.B.; Johnson, D.B. Microbiology of a wetland ecosystem constructed to remediate mine drainage from a heavy metal mine. Sci. Total Environ. 2005, 338, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Ehrich, S.; Behrens, D.; Lebedeva, E.; Ludwig, W.; Bock, E. A new obligately chemolithoautotrophic, nitrite-oxidizing bacterium, Nitrospira moscoviensis sp. nov. and its phylogenetic relationship. Arch. Microbiol. 1995, 164, 16–23. [Google Scholar] [CrossRef]
- Akcil, A.; Ciftci, H.; Deveci, H. Role and contribution of pure and mixed cultures of mesophiles in bioleaching of a pyritic chalcopyrite concentrate. Miner. Eng. 2007, 20, 310–318. [Google Scholar] [CrossRef]
- Fu, B.; Zhou, H.; Zhang, R.; Qiu, G. Bioleaching of chalcopyrite by pure and mixed cultures of Acidithiobacillus spp. and Leptospirillum ferriphilum. Int. Biodeterior. Biodegrad. 2008, 62, 109–115. [Google Scholar] [CrossRef]
- Wang, Y.; Zeng, W.; Qiu, G.; Chen, X.; Zhou, H. A moderately thermophilic mixed microbial culture for bioleaching of chalcopyrite concentrate at high pulp density. Appl. Environ. Microbiol. 2014, 80, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Schippers, A.; Sand, W. Bacterial Leaching of Metal Sulfides Proceeds by Two Indirect Mechanisms via Thiosulfate or via Polysulfides and Sulfur. Appl. Env. Microbiol. 1999, 65, 319–321. [Google Scholar] [CrossRef] [PubMed]
- Christel, S.; Dopson, M. Less may be more: Improving chalcopyrite bioleaching kinetics via sequential inoculation of acidophilic model species. Book of abstracts of the 10th International Conference on Establishment of Cooperation between Companies and Institutions in the Nordic Countries, the Baltic Sea Region and the World. In Proceedings of the Linnaeus Eco-Tech, Kalmar, Sweden, 21–23 November 2016; p. 169. [Google Scholar]
- Watling, H.R.; Collinson, D.M.; Li, J.; Mutch, L.A.; Perrot, F.A.; Rea, S.M.; Reith, F.; Watkin, E.L.J. Bioleaching of a low-grade copper ore, linking leach chemistry and microbiology. Miner. Eng. 2014, 56, 35–44. [Google Scholar] [CrossRef]
- Tao, J.; Liu, X.; Luo, X.; Teng, T.; Jiang, C.; Drewniak, L.; Yang, Z.; Yin, H. An integrated insight into bioleaching performance of chalcopyrite mediated by microbial factors: Functional types and biodiversity. Bioresour. Technol. 2021, 319, 124219. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Fe | Cu | S |
|---|---|---|---|
| Chalcopyrite | 29.7 | 30.2 | 33.8 |
| EPS Composition | Protein (µg/L) | Carbohydrates (µg/L) | Uronic Acids |
|---|---|---|---|
| Capsular | 16.3 | 6.0 | BDL * |
| Colloidal | 8.3 | 3.9 | BDL * |
| Isolated Strain | Strain Name, GenBank (Accession Number) | Identity, % | Reference |
|---|---|---|---|
| L. ferrodiazotrophum Ksh-L | Uncultured bacterium clone YTW-66-06 (EF409823.1) | 99.52 | [81] |
| Uncultured bacterium clone SLS-53-06 (EF409827.1) | 99.66 | [81] | |
| Uncultured bacterium clone YTW-96-06 (EF409843.1) | 99.59 | [81] | |
| Leptospirillum ferrodiazotrophum 5C (JN007036.1) | 96.60 | [82] | |
| Leptospirillum ferriphilum ATCC 49,881 (AF356829.1) | 91.02 | [20] | |
| Leptospirillum ferrooxidans WJ71 (AY495960.1) | 89.60 | [83] | |
| Nitrospira moscoviensis (X82558.1) | 81.22 | [84] |
| Bacteria | Extraction of Fe 30 Days | Extraction of Cu | Final | |||||
|---|---|---|---|---|---|---|---|---|
| g/L | % | g/L | % | pH | ORP, mV | |||
| Fe3+ | Fe2+ | Fe Total | Fe Total | |||||
| Control (uninoculated) | 0 | 0.672 | 0.672 | 5.6 | 0.52 | 4.1 | 1.8 | 520 |
| L. ferrodiazotropum Ksh-L | 1.096 | 0.448 | 1.544 | 12.8 | 0.884 | 6.9 | 1.7 | 600 |
| L. ferrodiazotropum Ksh-L + At. thiooxidans | 2.016 | 0.616 | 2.744 | 23.0 | 1.67 | 13.4 | 1.5 | 720 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khachatryan, A.; Vardanyan, N.; Willscher, S.; Sevoyan, G.; Zhang, R.; Vardanyan, A. Bioleaching of Chalcopyrite by a New Strain Leptospirillum ferrodiazotrophum Ksh-L Isolated from a Dump-Bioleaching System of Kashen Copper-Molybdenum Mine. Minerals 2024, 14, 26. https://doi.org/10.3390/min14010026
Khachatryan A, Vardanyan N, Willscher S, Sevoyan G, Zhang R, Vardanyan A. Bioleaching of Chalcopyrite by a New Strain Leptospirillum ferrodiazotrophum Ksh-L Isolated from a Dump-Bioleaching System of Kashen Copper-Molybdenum Mine. Minerals. 2024; 14(1):26. https://doi.org/10.3390/min14010026
Chicago/Turabian StyleKhachatryan, Anna, Narine Vardanyan, Sabine Willscher, Garegin Sevoyan, Ruiyong Zhang, and Arevik Vardanyan. 2024. "Bioleaching of Chalcopyrite by a New Strain Leptospirillum ferrodiazotrophum Ksh-L Isolated from a Dump-Bioleaching System of Kashen Copper-Molybdenum Mine" Minerals 14, no. 1: 26. https://doi.org/10.3390/min14010026