
Towards the Chemical Analysis of Diatoms’ Silicon Storage Pools: A Differential Centrifugation-Based Separation Approach

Abstract
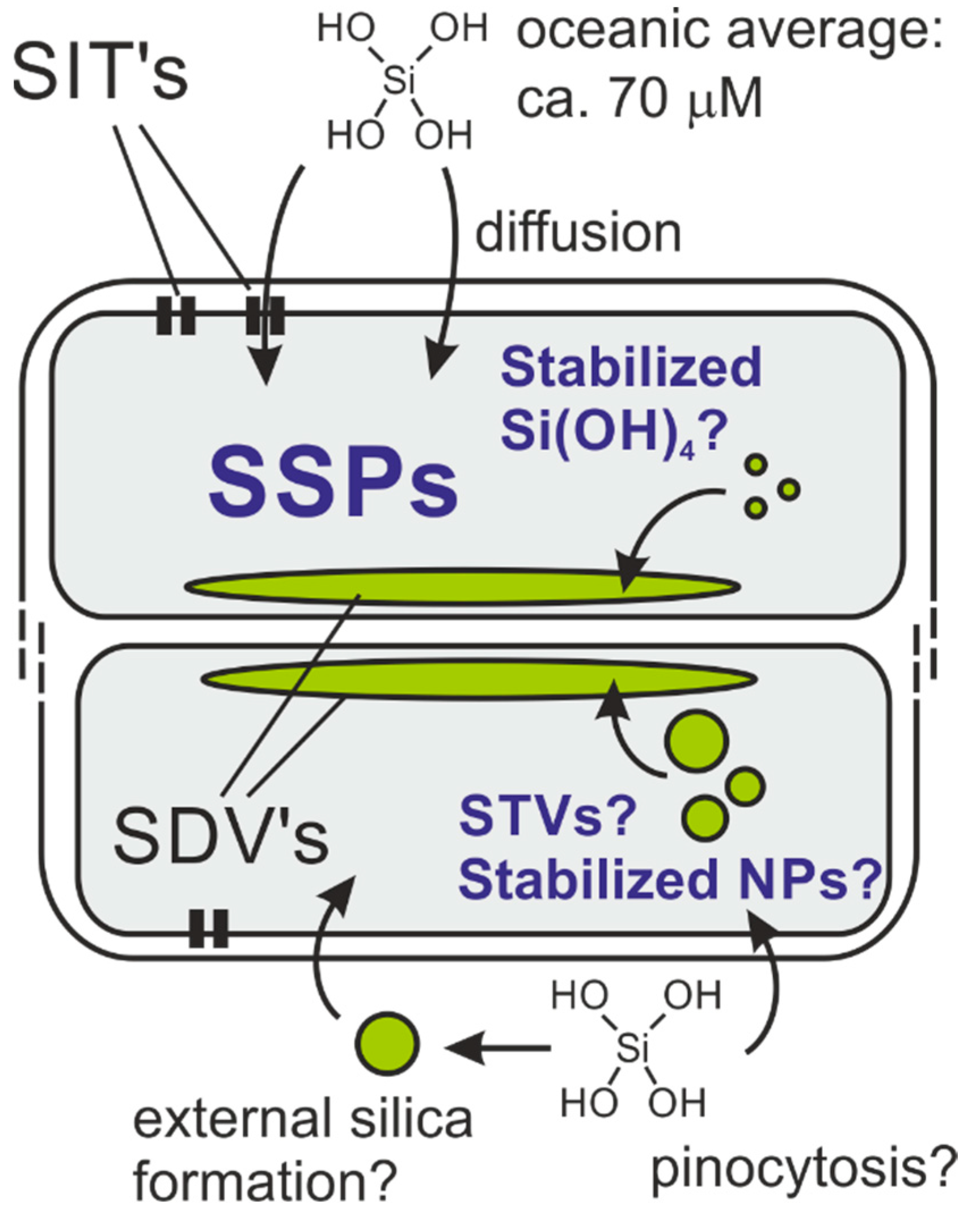
:1. Introduction
2. Materials and Methods
2.1. Cultivation
2.2. Molybdenum Blue Assay (MBA)
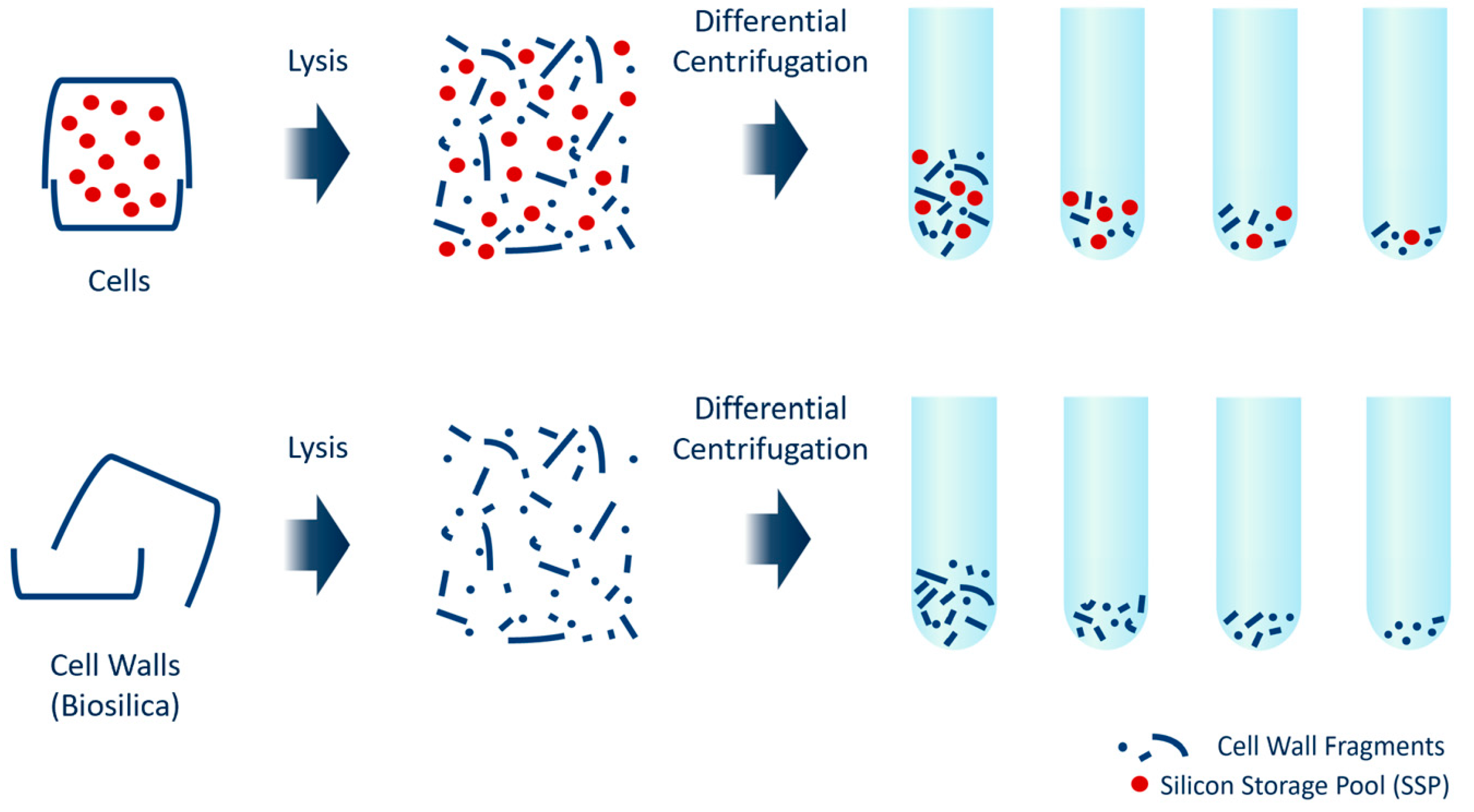
2.3. Preparation of Biosilica
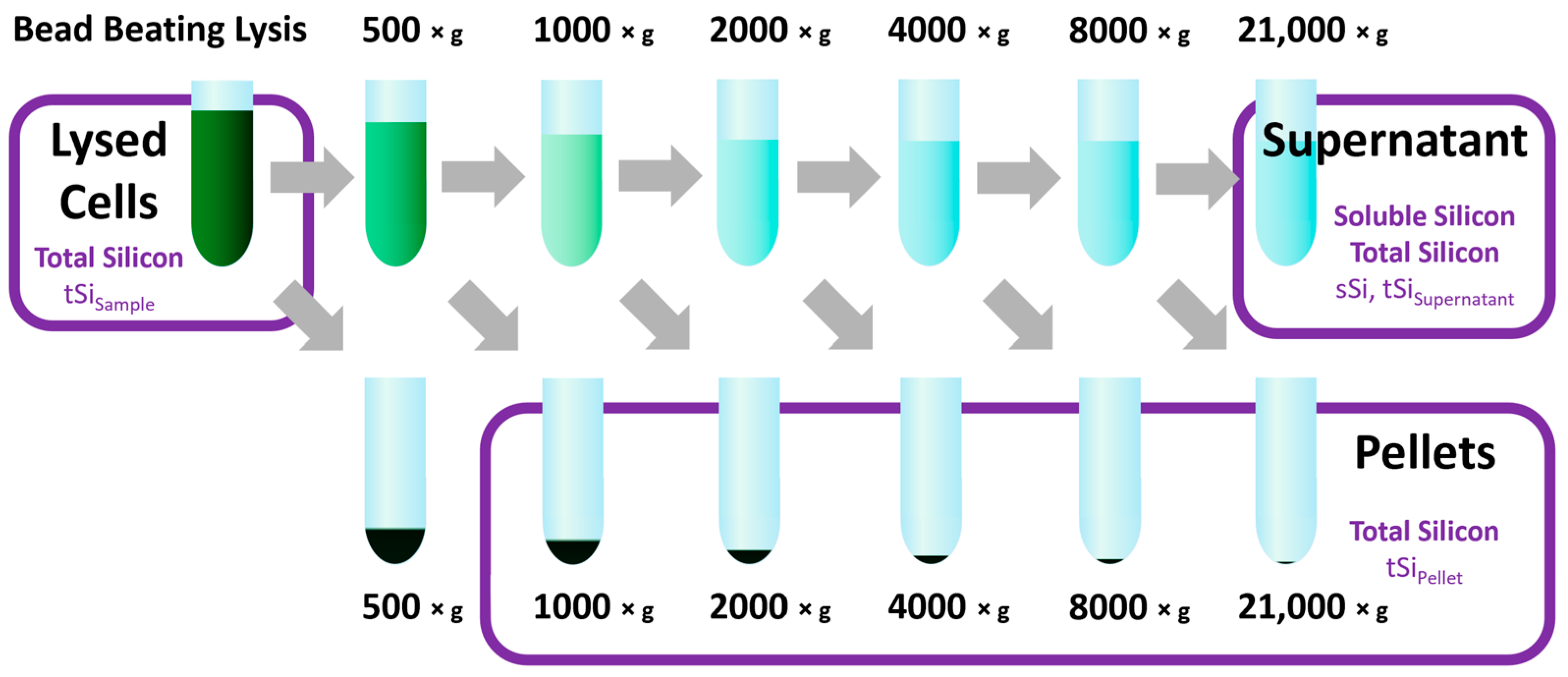
2.4. Bead-Beating Lysis
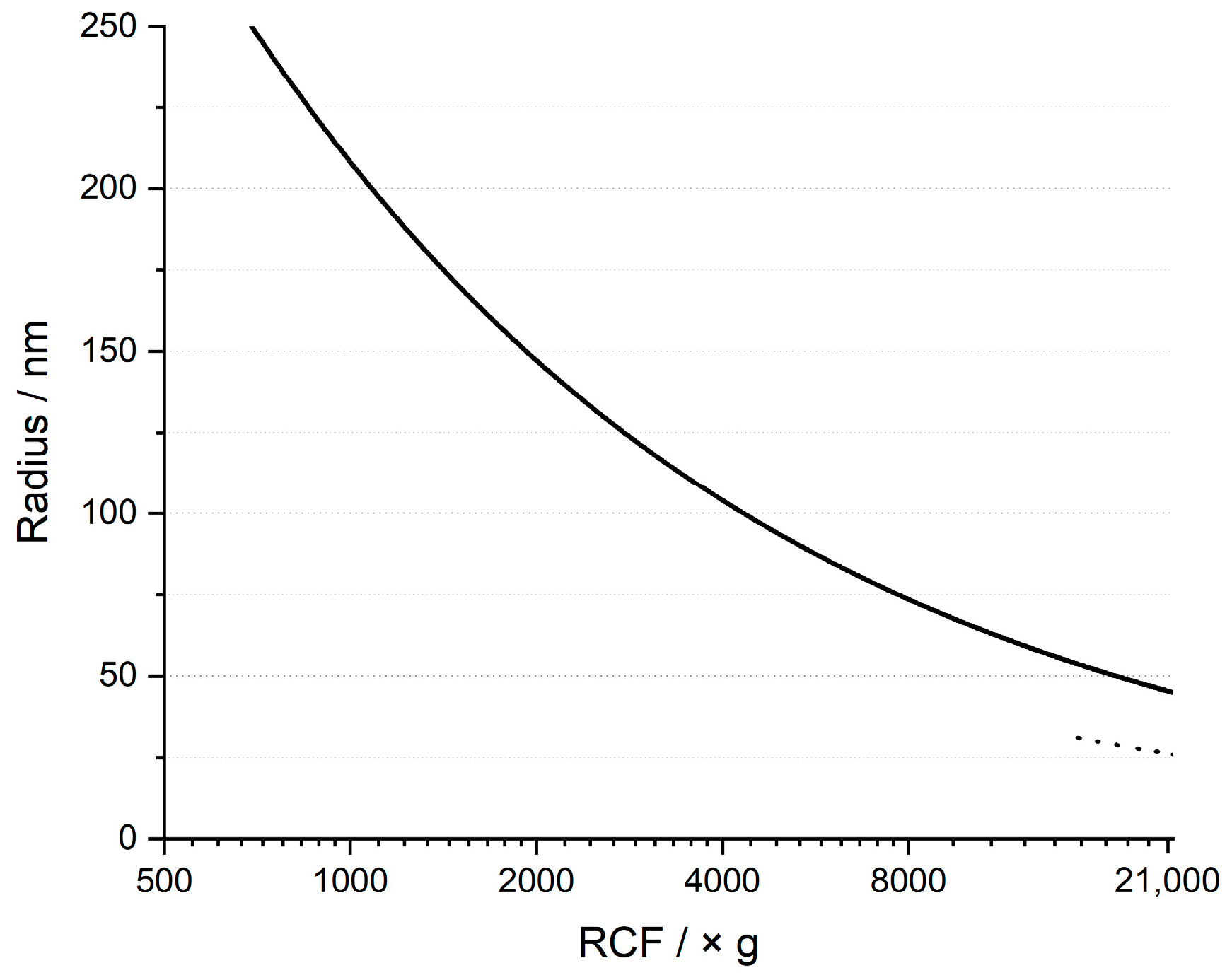
2.5. Differential Centrifugation
2.6. Sample Preparation and Calibration
2.7. Alkaline Treatment
2.7.1. Standard Procedure—Mild Alkaline Treatment
2.7.2. Autoclave-Induced Digestion (AID)
2.7.3. Oven-Induced Digestion (OID)
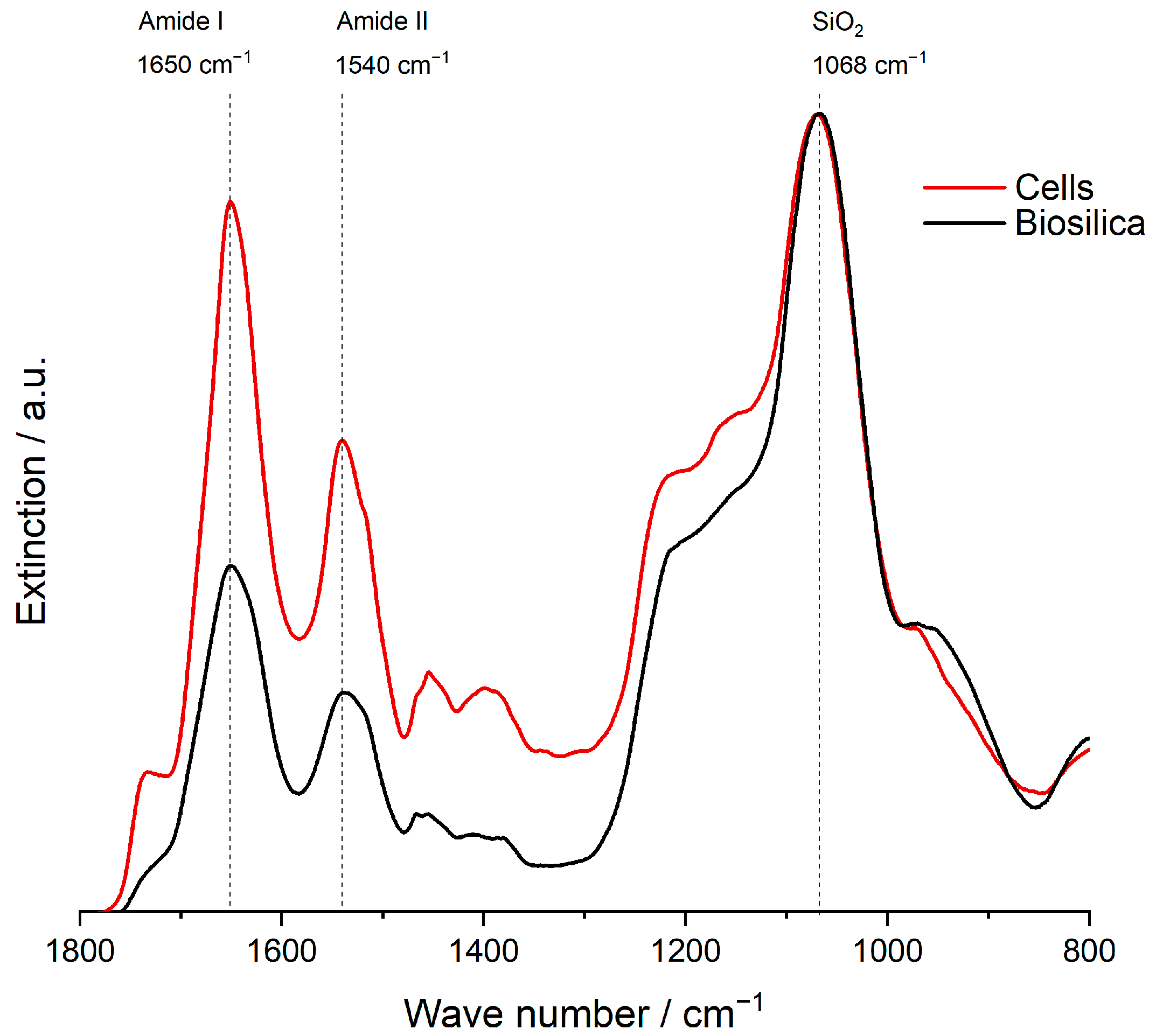
2.8. Characterization
3. Results
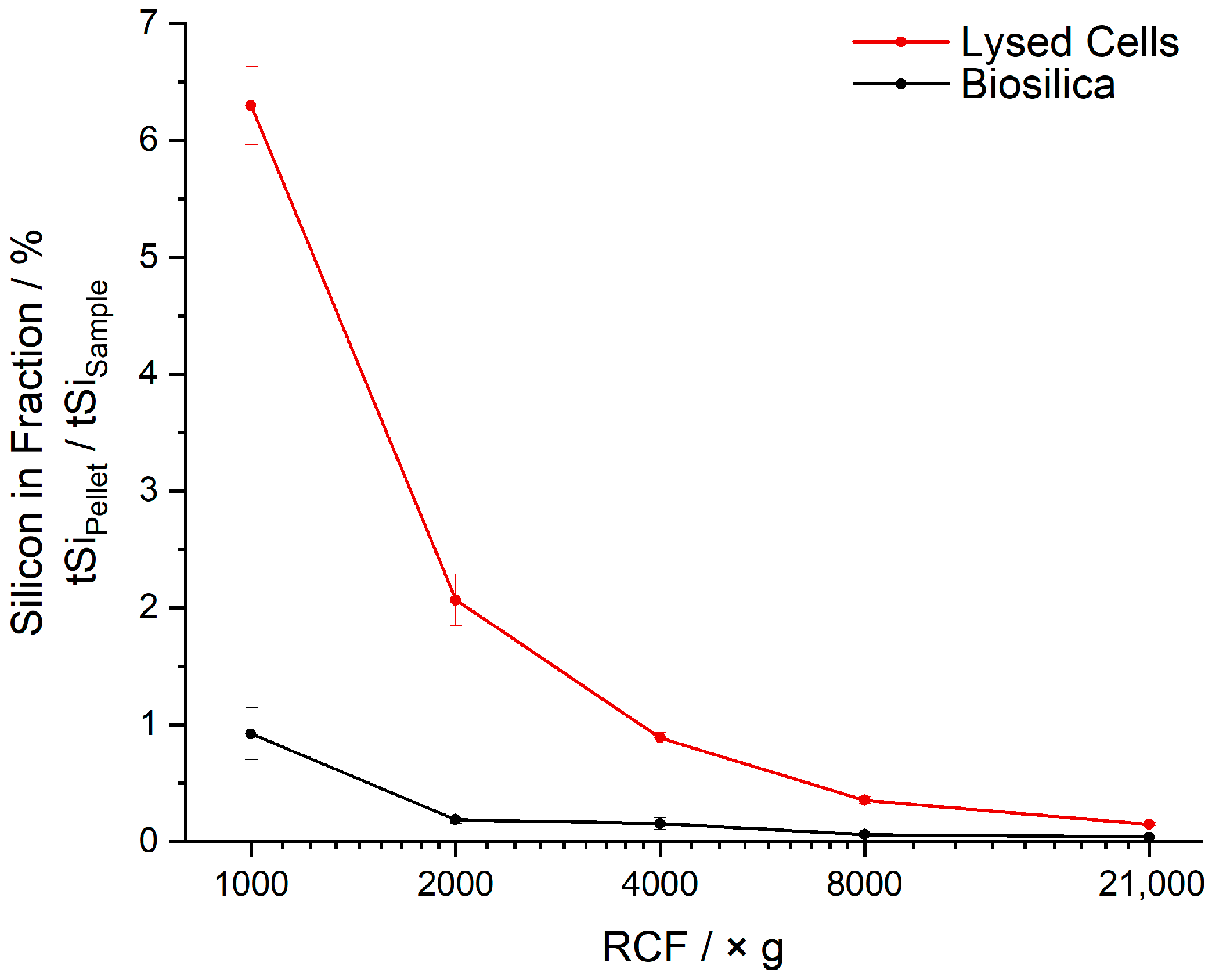
3.1. Cell Lysis, Differential Centrifugation and Silicon Quantification
3.2. Sedimentation Behavior of Cells and Biosilica after Bead-Beating Treatment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Armbrust, E.V. The life of diatoms in the world’s oceans. Nature 2009, 459, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Butler, T.; Kapoore, R.V.; Vaidyanathan, S. Phaeodactylum tricornutum: A diatom cell factory. Trends biotechnol. 2020, 38, 606–622. [Google Scholar] [CrossRef] [PubMed]
- Sonak, S.; Patil, K.; Devi, P.; D’Souza, L. Causes, human health impacts and control of harmful algal blooms: A comprehensive review. Environ. Pollut. Prot. 2018, 3, 40–55. [Google Scholar] [CrossRef]
- Levitan, O.; Dinamarca, J.; Hochman, G.; Falkowski, P.G. Diatoms: A fossil fuel of the future. Trends biotechnol. 2014, 32, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Merz, C.R.; Main, K.L. Microalgae (diatom) production—The aquaculture and biofuel nexus. In Proceedings of the 2014 Oceans—St. John’s, St. John’s, NL, Canada, 14–19 September 2014; pp. 1–10. [Google Scholar] [CrossRef]
- Yi, Z.; Xu, M.; Di, X.; Brynjolfsson, S.; Fu, W. Exploring valuable lipids in diatoms. Front. Mar. Sci. 2017, 4, 17. [Google Scholar] [CrossRef]
- Round, F.E.; Crawford, R.M.; Mann, D.G. The Diatoms; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar] [CrossRef]
- Reka, A.A.; Smirnov, P.V.; Belousov, P.; Durmishi, B.; Abbdesettar, L.; Aggrey, P.; Kabra Malpani, S.; Idrizi, H. Diatomaceous earth: A literature review. J. Nat. Sci. Math. UT 2022, 7, 256–268. [Google Scholar]
- Wang, Y.; Cai, J.; Jiang, Y.; Jiang, X.; Zhang, D. Preparation of biosilica structures from frustules of diatoms and their applications: Current state and perspectives. Appl. Microbiol. Biotechnol. 2013, 97, 453–460. [Google Scholar] [CrossRef]
- Zhang, D.Y.; Wang, Y.; Zhang, W.Q.; Pan, J.F.; Cai, J. Enlargement of diatom frustules pores by hydrofluoric acid etching at room temperature. J. Mater. Sci. 2013, 46, 5665–5671. [Google Scholar] [CrossRef]
- Gurel, A.; Yildiz, A. Diatom communities, lithofacies characteristics and paleo environmental interpretation of Pliocene diatomite deposits in the lhlara–Selime plain (Aksaray, Central Anatolia, Turkey). J. Asian Earth Sci. 2007, 30, 170–180. [Google Scholar] [CrossRef]
- Losic, D. Diatom Nanotechnology: Progress and Emerging Applications; The Royal Society of Chemistry: London, UK, 2018. [Google Scholar] [CrossRef]
- Losic, D.; Rosengarten, G.; Mitchell, J.G.; Voelcker, N.H. Pore architecture of diatom frustules: Potential nanostructured membranes for molecular and particle separations. J. Nanosci. Nanotechnol. 2006, 6, 982–988. [Google Scholar] [CrossRef]
- Grimann, M.; Fuhrmann-Lieker, T. Biological Photonic Crystals. In Organic and Hybrid Photonic Crystals; Comoretto, D., Ed.; Springer: Cham, Switzerland, 2015; pp. 57–74. [Google Scholar] [CrossRef]
- Kröger, N.; Brunner, E. Complex-Shaped Microbial Biominerals for Nanotechnology. WIREs Nanomed. Nanobiotechnol. 2014, 6, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Tramontano, C.; Chianese, G.; Terracciano, M.; de Stefano, L.; Rea, I. Nanostructured biosilica of diatoms: From water world to biomedical applications. Appl. Sci. 2020, 10, 6811. [Google Scholar] [CrossRef]
- Saoud, H.A.A.; Sprynskyy, M.; Pashaei, R.; Kawalec, M.; Pomastowski, P.; Buszewski, B. Diatom biosilica: Source, physical-chemical characterization, modification, and application. J. Sep. Sci. 2022, 45, 3362–3376. [Google Scholar] [CrossRef] [PubMed]
- Zobi, F. Diatom Biosilica in Targeted Drug Delivery and Biosensing Applications: Recent Studies. Micro 2022, 2, 342–360. [Google Scholar] [CrossRef]
- Perry, C.C.; Keeling-Tucker, T. Biosilicification: The role of the organic matrix in structure control. J. Biol. Inorg. Chem. 2000, 5, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Kolbe, F.; Brunner, E. Silicic acid uptake and storage by diatoms. In The Molecular Life of Diatoms, 1st ed.; Falciatore, A., Mock, T., Eds.; Springer: Cham, Switzerland, 2022; p. 349. [Google Scholar] [CrossRef]
- Heintze, C.; Formanek, P.; Pohl, D.; Hauptstein, J.; Rellinghaus, B.; Kröger, N. An intimate view into the silica deposition vesicles of diatoms. BMC Mater. 2020, 2, 11. [Google Scholar] [CrossRef]
- Lewin, J.C. Silicon metabolism in diatoms. II. Sources of silicon for growth of Navicula pelliculosa. Plant Physiol. 1955, 30, 129–134. [Google Scholar] [CrossRef]
- Lewin, J.C. Silicon metabolism in diatoms: III. Respiration and silicon uptake in Navicula pelliculosa. J. Gen. Physiol. 1955, 39, 1–10. [Google Scholar] [CrossRef]
- Azam, F.; Hemmingsen, B.B.; Volcani, B.E. Role of silicon in diatom metabolism. V. Silicic acid transport and metabolism in the heterotrophic diatom Nitzschia Alba. Arch. Microbiol. 1974, 97, 103–114. [Google Scholar] [CrossRef]
- Azam, F. Silicic-acid uptake in diatoms studied with [68Ge]germanic acid as tracer. Planta 1974, 121, 205–212. [Google Scholar] [CrossRef]
- Sullivan, C.W. Diatom mineralization of silicic acid. I. Si(OH)4 transport characteristics in Navicula Pelliculosa. J. Phycol. 1976, 12, 390–396. [Google Scholar] [CrossRef]
- Sullivan, C.W. Diatom mineralization of silicic acid. V. Energetic and macromolecular requirements for Si(OH)4 mineralization events during the cell cycle of Navicula Pelliculosa. J. Phycol. 1980, 16, 321–328. [Google Scholar] [CrossRef]
- Blank, G.S.; Sullivan, C.W. Diatom mineralization of silicic acid: III. Si(OH)4 binding and light dependent transport in Nitzschia angularis. Arch. Microbiol. 1979, 123, 157–164. [Google Scholar] [CrossRef]
- Sullivan, C.W. Diatom mineralization of silicic acid. IV. Kinetics of soluble Si pool formation in exponentially growing and synchronized Navicula Pelliculosa. J. Phycol. 1979, 15, 210–216. [Google Scholar] [CrossRef]
- Hildebrand, M. Diatoms, biomineralization processes, and genomics. Chem. Rev. 2008, 108, 4855–4874. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, M.; Volcani, B.E.; Gassmann, W.; Schroeder, J.I. A gene family of silicon transporters. Nature 1997, 385, 688–689. [Google Scholar] [CrossRef]
- Hildebrand, M.; Dahlin, K.; Volcani, B.E. Characterization of a silicon transporter gene family in Cylindrotheca fusiformis: Sequences, expression analysis, and identification of homologs in other diatoms. Mol. Gen. Genet. MGG 1998, 260, 480–486. [Google Scholar] [CrossRef]
- Thamatrakoln, K.; Hildebrand, M. Analysis of Thalassiosira pseudonana silicon transporters indicates distinct regulatory levels and transport activity through the cell cycle. Eukaryot. Cell 2007, 6, 271–279. [Google Scholar] [CrossRef]
- Knight, M.J.; Senior, L.; Nancolas, B.; Ratcliffe, S.; Curnow, P. Direct evidence of the molecular basis for biological silicon transport. Nat. Commun. 2016, 7, 11926. [Google Scholar] [CrossRef]
- Thamatrakoln, K.; Hildebrand, M. Approaches for functional characterization of diatom silicic acid transporters. J. Nanosci. Nanotechnol. 2005, 5, 158–166. [Google Scholar] [CrossRef]
- Thamatrakoln, K.; Hildebrand, M. Silicon uptake in diatoms revisited: A model for saturable and nonsaturable uptake kinetics and the role of silicon transporters. Plant Physiol. 2008, 146, 1397–1407. [Google Scholar] [CrossRef] [PubMed]
- Brasser, H.J.; van der Strate, H.J.; Gieskes, W.W.C.; Krijger, G.C.; Vrieling, E.G.; Wolterbeek, H.T. Compartmental analysis suggests macropinocytosis at the onset of diatom valve formation. Silicon 2012, 4, 39–49. [Google Scholar] [CrossRef]
- Vrieling, E.G.; Sun, Q.; Tian, M.; Kooyman, P.J.; Gieskes, W.W.C.; van Santen, R.A.; Sommerdijk, N.A.J.M. Salinity-dependent diatom biosilicification implies an important role of external ionic strength. Proc. Natl. Acad. Sci. USA 2007, 104, 10441–10446. [Google Scholar] [CrossRef] [PubMed]
- Annenkov, V.V.; Gordon, R.; Zelinskiy, S.N.; Danilovtseva, E.N. The probable mechanism for silicon capture by diatom algae: Assimilation of polycarbonic acids with diatoms—Is endocytosis a key stage in building of siliceous frustules? J. Phycol. 2020, 56, 1729–1737. [Google Scholar] [CrossRef]
- Thamatrakoln, K.; Kustka, A.B. When to say when: Can excessive drinking explain silicon uptake in diatoms? BioEssays 2009, 31, 322–327. [Google Scholar] [CrossRef]
- Tréguer, P.; Nelson, D.M.; Van Bennekom, A.J.; DeMaster, D.J.; Leynaert, A.; Quéguiner, B. The silica balance in the world ocean: A reestimate. Science 1995, 268, 375–379. [Google Scholar] [CrossRef]
- Iler, R.K. The Chemistry of Silica: Solubility, Polymerization, Colloid and Surface Properties and Biochemistry of Silica; John Wiley & Sons Ltd.: New York, NY, USA, 1979. [Google Scholar] [CrossRef]
- Morey, G.W.; Fournier, R.O.; Rowe, J.J. The solubility of amorphous silica at 25 °C. J. Geophys. Res. 1964, 69, 1995–2002. [Google Scholar] [CrossRef]
- Coombs, J.; Darley, W.M.; Holm-Hansen, O.; Volcani, B.E. Studies on the biochemistry and fine structure of silica shell formation in diatoms. Chemical composition of Navicula pelliculosa during silicon-starvation synchrony. Plant Physiol. 1967, 42, 1601–1606. [Google Scholar] [CrossRef]
- Binder, B.J.; Chisholm, S.W. Changes in the soluble silicon pool size in the marine diatom Thalassiosira weissflogii. Mar. Biol. Lett. 1980, 1, 205–212. [Google Scholar]
- Brzezinski, M.A.; Conley, D.J. Silicon deposition during the cell cycle of Thalassiosira weissflogii (Bacillariophyceae) determined using dual rhodamine 123 and propidium iodide staining. J. Phycol. 1994, 30, 45–55. [Google Scholar] [CrossRef]
- Martin-Jézéquel, V.; Hildebrand, M.; Brzezinski, M.A. Silicon metabolism in diatoms: Implications for growth. J. Phycol. 2000, 36, 821–840. [Google Scholar] [CrossRef]
- Chisholm, S.W.; Azam, F.; Eppley, R.W. Silicic acid incorporation in marine diatoms on light:dark cycles: Use as an assay for phased cell division. Limnol. Oceanogr. 1978, 23, 518–529. [Google Scholar] [CrossRef]
- Kumar, S.; Rechav, K.; Kaplan-Ashiri, I.; Gal, A. Imaging and quantifying homeostatic levels of intracellular silicon in diatoms. Sci. Adv. 2020, 6, eaaz7554. [Google Scholar] [CrossRef] [PubMed]
- Kinrade, S.D.; Gillson, A.M.; Knight, C.T. Silicon-29 NMR evidence of a transient hexavalent silicon complex in the diatom Navicula pelliculosa. J. Chem. Soc. Dalton Trans. 2002, 3, 307–309. [Google Scholar] [CrossRef]
- Werner, D. Die Kieselsäure im Stoffwechsel von Cyclotella cryptica Reimann, Lewin und Guillard. Arch. Mikrobiol. 1966, 55, 278–308. [Google Scholar] [CrossRef]
- Spinthaki, A.; Demadis, K.D. Modeling Silicon Pools in Diatoms Using the Chemistry Toolbox. In Diatom Morphogenesis; Wiley Online Library: Hoboken, NJ, USA, 2021; pp. 365–382. [Google Scholar] [CrossRef]
- Kröger, N.; Deutzmann, R.; Bergsdorf, C.; Sumper, M. Species-specific polyamines from diatoms control silica morphology. Proc. Natl. Acad. Sci. USA 2000, 97, 14133–14138. [Google Scholar] [CrossRef]
- Bridoux, M.C.; Keil, R.G.; Ingalls, A.E. Analysis of natural diatom communities reveals novel insights into the diversity of long chain polyamine (LCPA) structures involved in silica precipitation. Org. Geochem. 2012, 47, 9–21. [Google Scholar] [CrossRef]
- Gröger, C.; Sumper, M.; Brunner, E. Silicon uptake and metabolism of the marine diatom Thalassiosira pseudonana: Solid-state 29Si NMR and fluorescence microscopic studies. J. Struct. Biol. 2008, 161, 55–63. [Google Scholar] [CrossRef]
- Brunner, E.; Gröger, C.; Lutz, K.; Richthammer, P.; Spinde, K.; Sumper, M. Analytical studies of silica biomineralization: Towards an understanding of silica processing by diatoms. Appl. Microbiol. Biotechnol. 2009, 84, 607–616. [Google Scholar] [CrossRef]
- Schmid, A.M.M.; Schulz, D. Wall morphogenesis in diatoms: Deposition of silica by cytoplasmic vesicles. Protoplasma 1979, 100, 267–288. [Google Scholar] [CrossRef]
- Sullivan, C.W. Silicification by diatoms. In Ciba Foundation Symposium 121-Silicon Biochemistry: Silicon Biochemistry: Ciba Foundation Symposium 121; Evered, D., O’Connor, M., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2007; p. 64. [Google Scholar] [CrossRef]
- Grachev, M.A.; Bedoshvili, Y.D.; Gerasimov, E.Y.; Zaikovskii, V.I.; Gneusheva, K.V.; Likhoshway, Y.V. Silica-containing inclusions in the cytoplasm of diatom Synedra acus. Dokl. Biochem. Biophys. 2017, 472, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Kröger, N.; Bergsdorf, C.; Sumper, M. A new calcium binding glycoprotein family constitutes a major diatom cell wall component. EMBO J. 1994, 13, 4676–4683. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.J.; Waters, R.E.; Taylor, F.J.R. A broad spectrum artificial medium for coastal and open ocean phytoplankton. J. Phycol. 1980, 16, 28–35. [Google Scholar] [CrossRef]
- Bunting, W.E. Determination of soluble silica in very low concentrations. Ind. Eng. Chem. Anal. Ed. 1944, 16, 612–615. [Google Scholar] [CrossRef]
- Koistinen, J.; Sjöblom, M.; Spilling, K. Determining dissolved and biogenic silica. In Biofuels from Algae; Spilling, K., Ed.; Humana: New York, NY, USA, 2018; pp. 95–101. [Google Scholar] [CrossRef]
- Elliott, C.L.; Snyder, G.H. Autoclave-induced digestion for the colorimetric determination of silicon in rice straw. J. Agric. Food Chem. 1991, 39, 1118–1119. [Google Scholar] [CrossRef]
- Kraska, J.E.; Breitenbeck, G.A. Simple, robust method for quantifying silicon in plant tissue. Commun. Soil Sci. Plant Anal. 2010, 41, 2075–2085. [Google Scholar] [CrossRef]
- Clissold, F.J.; Clark, X.; Savage, T.; Simpson, S.J. A rapid, precise and low-cost method to quantify silicon for the determination of food intake and utilisation for insect herbivores. Austral Entomol. 2018, 57, 220–227. [Google Scholar] [CrossRef]
- Coradin, T.; Eglin, D.; Livage, J. The silicomolybdic acid spectrophotometric method and its application to silicate/biopolymer interaction studies. Spectroscopy 2004, 18, 567–576. [Google Scholar] [CrossRef]
- Truesdale, V.W.; Smith, C.J. The spectrophotometric characteristics of aqueous solutions of α- and β-molybdosilicic acids. Analyst 1975, 100, 797–805. [Google Scholar] [CrossRef]
- Truesdale, V.W.; Smith, P.J.; Smith, C.J. Kinetics of α- and β-molybdosilicic acid formation. Analyst 1979, 104, 897–918. [Google Scholar] [CrossRef]
- Hildebrand, M.; Lerch, S.J.; Shrestha, R.P. Understanding diatom cell wall silicification—Moving forward. Front. Mar. Sci. 2018, 5, 125. [Google Scholar] [CrossRef]
- Barth, A. Infrared spectroscopy of proteins. Biochim. Biophys. Acta (BBA)-Bioenerg. 2007, 1767, 1073–1101. [Google Scholar] [CrossRef] [PubMed]
- Kammer, M.; Hedrich, R.; Ehrlich, H.; Popp, J.; Brunner, E.; Krafft, C. Spatially resolved determination of the structure and composition of diatom cell walls by Raman and FTIR imaging. Anal. Bioanal. Chem. 2010, 398, 509–517. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RCF × g | Time min | Centrifuged Volume µL | Supernatant µL | Pellet µL |
|---|---|---|---|---|
| 500 | 5 | 1400 | 1200 | 200 a |
| 1000 | 5 | 1200 | 1100 | 100 b |
| 2000 | 5 | 1100 | 1050 | 50 b |
| 4000 | 5 | 1050 | 1000 | 50 b |
| 8000 | 5 | 1000 | 950 | 50 b |
| 21,000 | 15 | 950 | 900 | 50 b,c |
| RCF × g | tSiPellet/tSiSample in %, Lysed Cells | tSiPellet/tSiSample in %, Biosilica | ||||
|---|---|---|---|---|---|---|
| Sample A | Sample B | Average | Sample A | Sample B | Average | |
| 1000 | 6.63 | 5.97 | 6.30 | 1.15 | 0.70 | 0.92 |
| 2000 | 2.29 | 1.85 | 2.07 | 0.22 | 0.16 | 0.19 |
| 4000 | 0.94 | 0.84 | 0.89 | 0.10 | 0.21 | 0.16 |
| 8000 | 0.39 | 0.32 | 0.36 | 0.06 | 0.06 | 0.06 |
| 21,000 | 0.17 | 0.13 | 0.15 | 0.04 | 0.04 | 0.04 |
| Sum | 10.41 | 9.11 | 9.76 | 1.57 | 1.17 | 1.37 |
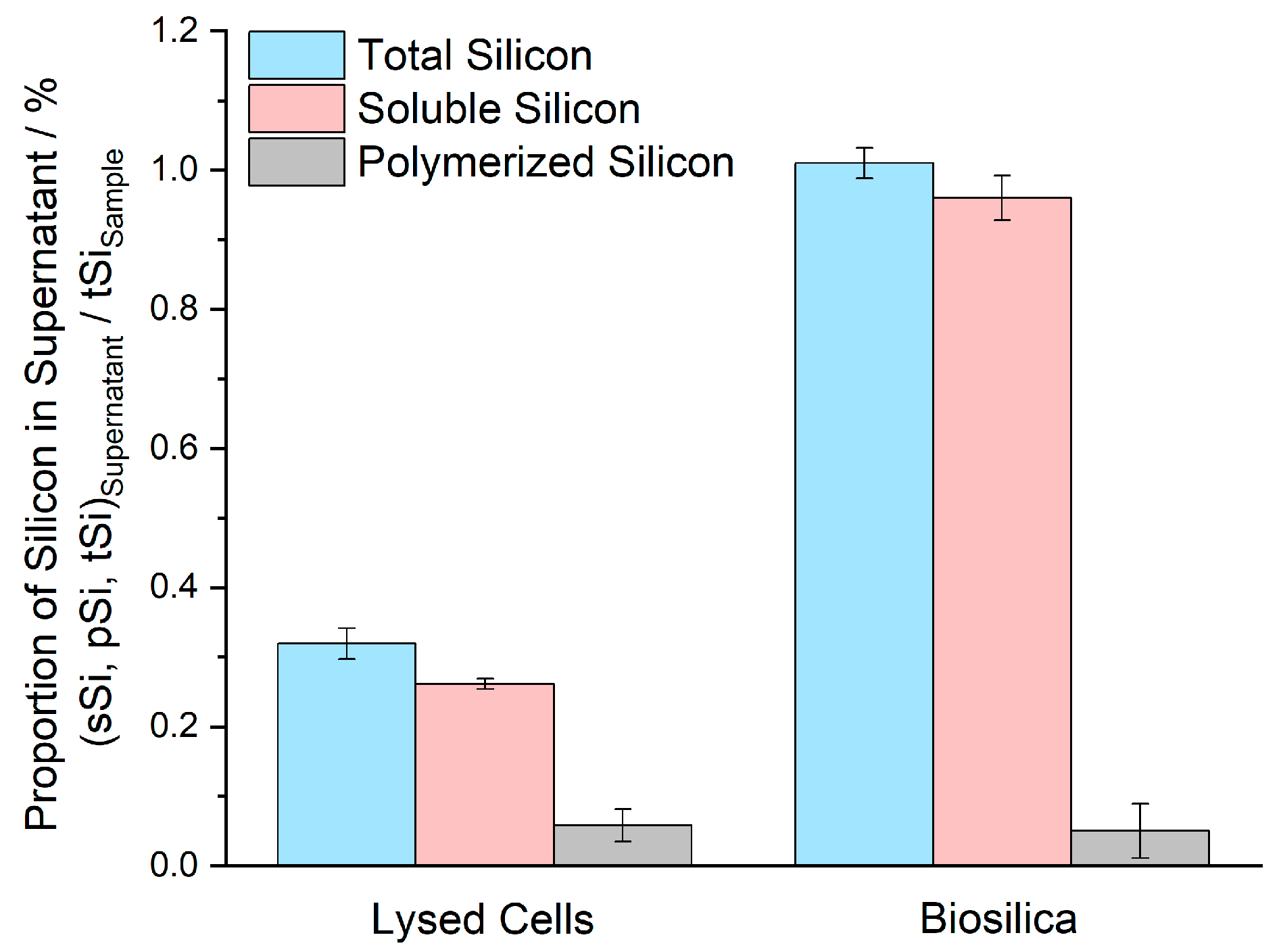
| Silicon in Supernatant | Lysed Cells % of tSisample | Biosilica % of tSiSample | ||||
|---|---|---|---|---|---|---|
| Sample A | Sample B | Average | Sample A | Sample B | Average | |
| tSi | 0.34 | 0.30 | 0.32 | 0.99 | 1.03 | 1.01 |
| sSi | 0.27 | 0.25 | 0.26 | 0.93 | 0.99 | 0.96 |
| pSi a | 0.07 | 0.04 | 0.06 | 0.06 | 0.04 | 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reichelt, T.; Bode, T.; Jordan, P.-F.; Brunner, E. Towards the Chemical Analysis of Diatoms’ Silicon Storage Pools: A Differential Centrifugation-Based Separation Approach. Minerals 2023, 13, 653. https://doi.org/10.3390/min13050653
Reichelt T, Bode T, Jordan P-F, Brunner E. Towards the Chemical Analysis of Diatoms’ Silicon Storage Pools: A Differential Centrifugation-Based Separation Approach. Minerals. 2023; 13(5):653. https://doi.org/10.3390/min13050653
Chicago/Turabian StyleReichelt, Tobias, Tobias Bode, Paul-Felix Jordan, and Eike Brunner. 2023. "Towards the Chemical Analysis of Diatoms’ Silicon Storage Pools: A Differential Centrifugation-Based Separation Approach" Minerals 13, no. 5: 653. https://doi.org/10.3390/min13050653